
Impact of Various Visible Spectra on Attached Microalgal Growth on Palm Decanter Cake in Triggering Protein, Carbohydrate, and Lipid to Biodiesel Production

 ,
, 
 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Microalgal Stock
2.2. Characterization of the Palm Decanter Cake
2.3. Experimental Setup for Attached Microalgal Growth
2.4. Analytical Methods
2.4.1. Attached Microalgal Biomass Productivity
2.4.2. Attached Microalgal Biochemical Productivity
2.4.3. Profile of Fatty Acid Methyl Esters
2.4.4. Analysis of the Net Energy Ratio (NER)
3. Results and Discussion
3.1. Characterization of Palm Decanter Cake
3.2. Attached Microalgal Growth on Palm Decanter Cake under Various Visible Spectra of Irradiation
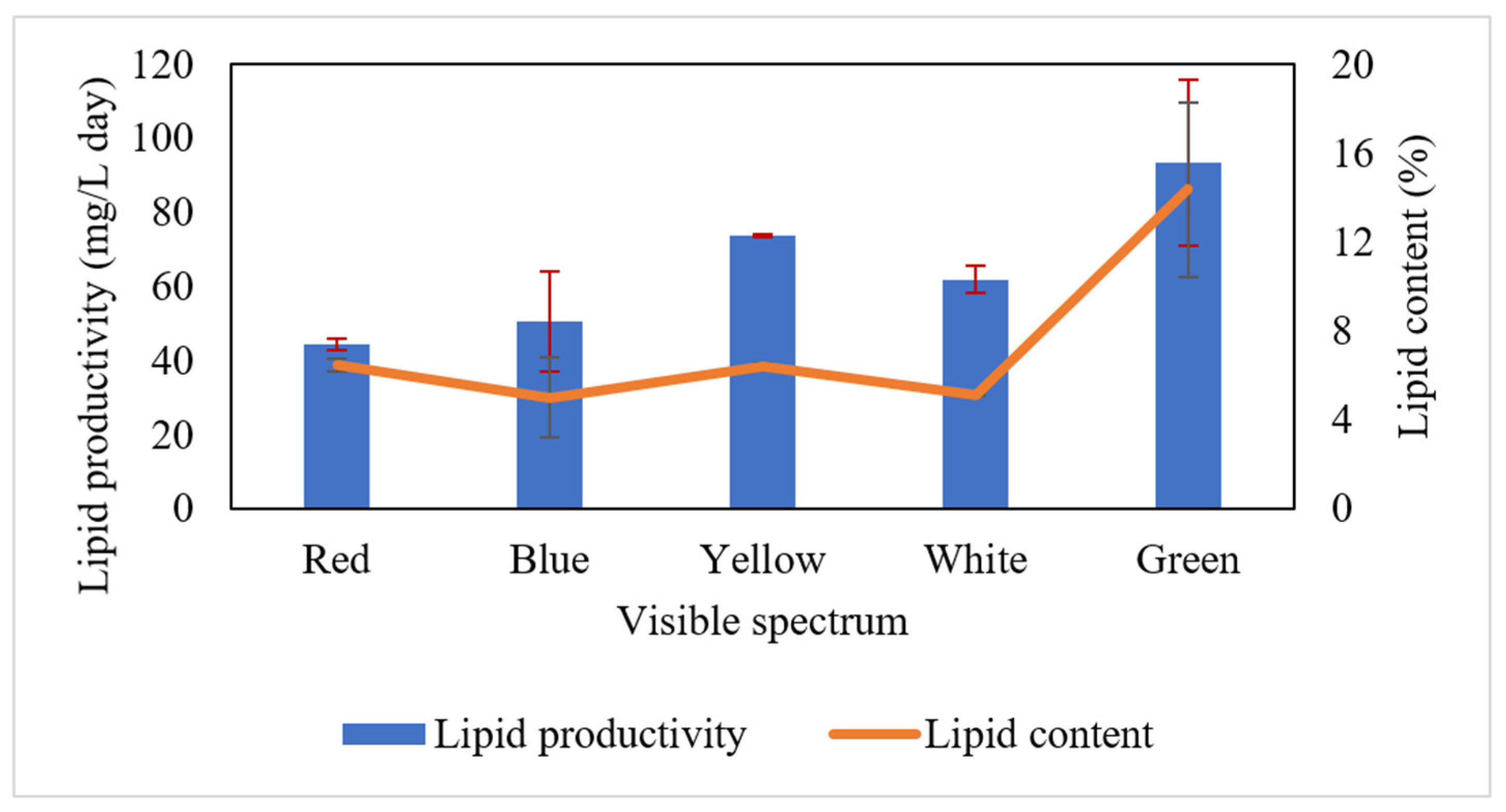
3.3. Lipid Accumulation from Attached Microalgae Grown on Palm Decanter Cake under Various Visible Spectra of Irradiation
3.4. Protein and Carbohydrate Contents of Attached Microalgae Grown on Palm Decanter Cake under Various Visible Spectra of Irradiation
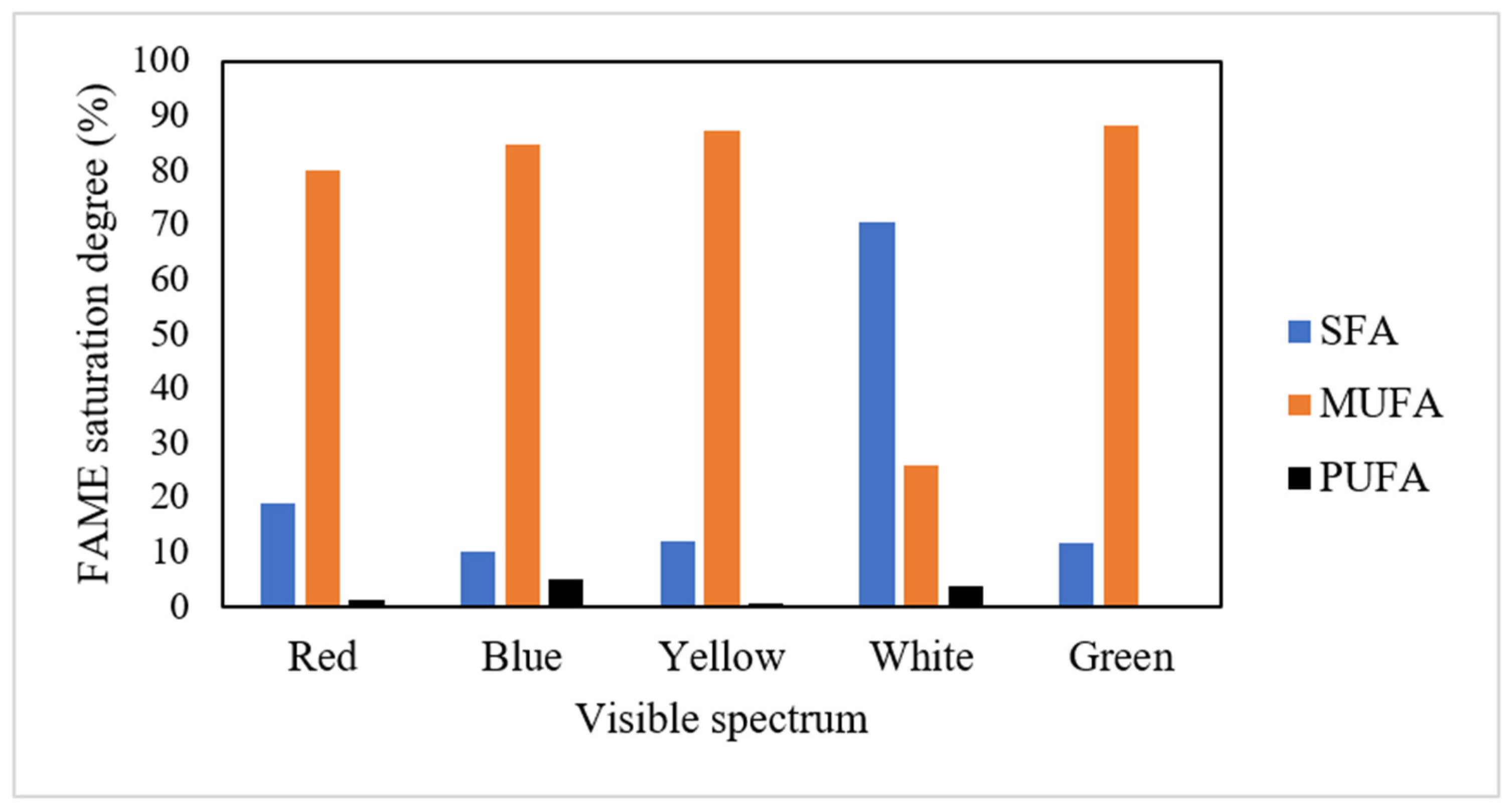
3.5. Biodiesel Profile Derived from Attached Microalgal Grown on Palm Decanter Cake under Various Visible Spectra of Irradiation
3.6. Energy Feasibility of Lipid Production from Attached Microalgal Grown on Palm Decanter Cake under Various Visible Spectra of Irradiation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Palm decanter cake | PDC |
| Bold’s Basal medium | BBM |
| Net energy ratio | NER |
| Light-emitting diode | LED |
| Photosynthetic active radiation | PAR |
| Fatty acid methyl ester | FAME |
| Monounsaturated fatty acid | MUFA |
| Polyunsaturated fatty acid | PUFA |
| Saturated fatty acid | SFA |
| Life cycle analysis | LCA |
References
- Chhandama, M.V.L.; Satyan, K.B.; Changmai, B.; Vanlalveni, C.; Rokhum, S.L. Microalgae as a Feedstock for the Production of Biodiesel: A Review. Bioresour. Technol. Rep. 2021, 15, 100771. [Google Scholar] [CrossRef]
- Zhu, L.; Nugroho, Y.K.; Shakeel, S.R.; Li, Z.; Martinkauppi, B.; Hiltunen, E. Using Microalgae to Produce Liquid Transportation Biodiesel: What Is Next? Renew. Sustain. Energy Rev. 2017, 78, 391–400. [Google Scholar] [CrossRef]
- Mohd-Sahib, A.A.; Lim, J.W.; Lam, M.K.; Uemura, Y.; Isa, M.H.; Ho, C.D.; Kutty, S.R.M.; Wong, C.Y.; Rosli, S.S. Lipid for Biodiesel Production from Attached Growth Chlorella Vulgaris Biomass Cultivating in Fluidized Bed Bioreactor Packed with Polyurethane Foam Material. Bioresour. Technol. 2017, 239, 127–136. [Google Scholar] [CrossRef]
- Daneshvar, E.; Sik Ok, Y.; Tavakoli, S.; Sarkar, B.; Shaheen, S.M.; Hong, H.; Luo, Y.; Rinklebe, J.; Song, H.; Bhatnagar, A. Insights into Upstream Processing of Microalgae: A Review. Bioresour. Technol. 2021, 329, 124870. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, X.; Jiang, Z.; Wang, X.; Wang, Y.; Cao, L.; Zhang, X. Effect of Light Spectra on Microalgal Biofilm: Cell Growth, Photosynthetic Property, and Main Organic Composition. Renew. Energy 2020, 157, 83–89. [Google Scholar] [CrossRef]
- Dalirian, N.; Abedini Najafabadi, H.; Movahedirad, S. Surface Attached Cultivation and Filtration of Microalgal Biofilm in a Ceramic Substrate Photobioreactaor. Algal Res. 2021, 55, 102239. [Google Scholar] [CrossRef]
- Embrandiri, A.; Fatemeh Rupani, P.; Ahmed Ismail, S.; Pratap Singh, R.; Hakimi Ibrahim, M.; Omar Abd Kadir, M. The Effect of Oil Palm Decanter Cake on the Accumulation of Nutrients and the Stomatal Opening of Solanum Melongena (Brinjal) Plants. Int. J. Recycl. Org. Waste Agric. 2016, 5, 141–147. [Google Scholar] [CrossRef]
- Abomohra, A.E.F.; Shang, H.; El-Sheekh, M.; Eladel, H.; Ebaid, R.; Wang, S.; Wang, Q. Night Illumination Using Monochromatic Light-Emitting Diodes for Enhanced Microalgal Growth and Biodiesel Production. Bioresour. Technol. 2019, 288, 121514. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef]
- Rawindran, H.; Leong, W.H.; Suparmaniam, U.; Liew, C.S.; Raksasat, R.; Kiatkittipong, W.; Mohamad, M.; Ghani, N.A.; Abdelfattah, E.A.; Lam, M.K.; et al. Residual Palm Kernel Expeller as the Support Material and Alimentation Provider in Enhancing Attached Microalgal Growth for Quality Biodiesel Production. J. Environ. Manag. 2022, 316, 115225. [Google Scholar] [CrossRef]
- Hemamalini, R.; Lim, J.W.; Leong, W.H.; Chew, T.L.; Lam, M.K.; Uganeeswary, S.; Mohamad, M.; Almendrala, M.C.; Shamsuddin, R. Utilization of Solid Palm Kernel Expeller for Attached Growth of Chlorella vulgaris Sp. In Proceedings of the AIP Conference Proceedings; American Institute of Physics Inc.: College Park, MD, USA, 9 June 2022; Volume 2454. [Google Scholar] [CrossRef]
- Zhong, Y.; Jin, P.; Cheng, J.J. A Comprehensive Comparable Study of the Physiological Properties of Four Microalgal Species under Different Light Wavelength Conditions. Planta 2018, 248, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Boni, J.; Aida, S.; Leila, K.; Selmani, N.; Mirghani, M.E.S.; Alam, M.Z. To Cite This Article: Nabila Selmani et Al. IOP Conf. Ser. Earth Environ. Sci. 2013, 16, 12006. [Google Scholar] [CrossRef]
- Leong, W.H.; Lim, J.W.; Lam, M.K.; Lam, S.M.; Sin, J.C.; Samson, A. Novel Sequential Flow Baffled Microalgal-Bacterial Photobioreactor for Enhancing Nitrogen Assimilation into Microalgal Biomass Whilst Bioremediating Nutrient-Rich Wastewater Simultaneously. J. Hazard. Mater. 2021, 409, 124455. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Wing, M.T.; Silaban, A.; Barnett, J.; Rusch, K.A. Light Irradiance and Spectral Distribution Effects on Microalgal Bioreactors. Eng. Life Sci. 2014, 14, 574–580. [Google Scholar] [CrossRef]
- Adam, M.A.; Sulaiman, A.; Said, C.M.S.; Som, A.M.; Tabatabaei, M. Enhanced Rigidity of Natural Polymer Composite Developed from Oil Palm Decanter Cake. BioResources 2015, 10, 932–942. [Google Scholar] [CrossRef]
- Choi, H.J. Agricultural Biowaste, Rice Bran, as Carbon Source to Enhance Biomass and Lipid Production: Analysis with Various Growth Rate Models. Water Sci. Technol. 2020, 82, 1120–1130. [Google Scholar] [CrossRef]
- Lessard, P.; Bihan, Y. le Fixed Film Processes. In Handbook of Water and Wastewater Microbiology; Academic Press: Cambridge, MA, USA; Elsevier: Cambridge, MA, USA, 2003; pp. 317–336. [Google Scholar] [CrossRef]
- Metsoviti, M.N.; Papapolymerou, G.; Karapanagiotidis, I.T.; Katsoulas, N. Effect of Light Intensity and Quality on Growth Rate and Composition of Chlorella Vulgaris. Plants 2020, 9, 31. [Google Scholar] [CrossRef]
- Blair, M.F.; Kokabian, B.; Gude, V.G. Light and Growth Medium Effect on Chlorella Vulgaris Biomass Production. J. Environ. Chem. Eng. 2014, 2, 665–674. [Google Scholar] [CrossRef]
- Yun, Y.S.; Park, J.M. Attenuation of Monochromatic and Polychromatic Lights in Chlorella Vulgaris Suspensions. Appl. Microbiol. Biotechnol. 2001, 55, 765–770. [Google Scholar] [CrossRef]
- Lysenko, V.; Kosolapov, A.; Usova, E.; Tatosyan, M.; Varduny, T.; Dmitriev, P.; Rajput, V.; Krasnov, V.; Kunitsina, A. Chlorophyll Fluorescence Kinetics and Oxygen Evolution in Chlorella Vulgaris Cells: Blue vs. Red Light. J. Plant Physiol. 2021, 258–259, 153392. [Google Scholar] [CrossRef]
- Acuapan-Hernandez, J.; Cañizares-Villanueva, R.O.; Cristiani-Urbina, E. Red Light and Nitrogen Depletion Stimulate the Synthesis of Lipids and N-Alkadienes Susceptible to Be Used as Biofuels in Botryococcus Braunii UTEX 2441 (Race A). Biotechnol. Ind. J. 2017, 13, 155. [Google Scholar]
- Sharma, N.; Fleurent, G.; Awwad, F.; Cheng, M.; Meddeb-Mouelhi, F.; Budge, S.M.; Germain, H.; Desgagné-Penix, I. Red Light Variation an Effective Alternative to Regulate Biomass and Lipid Profiles in Phaeodactylum Tricornutum. Appl. Sci. 2020, 10, 2531. [Google Scholar] [CrossRef]
- Yoshioka, M.; Yago, T.; Yoshie-Stark, Y.; Arakawa, H.; Morinaga, T. Effect of High Frequency of Intermittent Light on the Growth and Fatty Acid Profile of Isochrysis Galbana. Aquaculture 2012, 338–341, 111–117. [Google Scholar] [CrossRef]
- Belaiba, A.; Bouharat, D.; Malvis, A.; Hodaifa, G. Feasibility of the Hybrid Use of Chlorella Vulgaris Culture with the Conventional Biological Treatment in Urban Wastewater Treatment Plants. Processes 2021, 9, 1640. [Google Scholar] [CrossRef]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for Oil: Strain Selection, Induction of Lipid Synthesis and Outdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef]
- Chen, T.; Liu, J.; Guo, B.; Ma, X.; Sun, P.; Liu, B.; Chen, F. Light Attenuates Lipid Accumulation While Enhancing Cell Proliferation and Starch Synthesis in the Glucose-Fed Oleaginous Microalga Chlorella Zofingiensis. Sci. Rep. 2015, 5, 14936. [Google Scholar] [CrossRef]
- Muñoz, I.L.; Bernard, O. Modeling the Influence of Temperature, Light Intensity and Oxygen Concentration on Microalgal Growth Rate. Processes 2021, 9, 496. [Google Scholar] [CrossRef]
- Wong, Y. Effect of Different Light Sources on Algal Biomass and Lipid Production in Internal Leds-Illuminated Photobioreactor. J. Mar. Biol. Aquac. 2016, 2, 8. [Google Scholar] [CrossRef]
- Vadiveloo, A.; Moheimani, N.R.; Cosgrove, J.J.; Bahri, P.A.; Parlevliet, D. Effect of Different Light Spectra on the Growth and Productivity of Acclimated Nannochloropsis Sp. (Eustigmatophyceae). Algal Res. 2015, 8, 121–127. [Google Scholar] [CrossRef]
- Mutaf, T.; Oz, Y.; Kose, A.; Elibol, M.; Oncel, S.S. The Effect of Medium and Light Wavelength towards Stichococcus Bacillaris Fatty Acid Production and Composition. Bioresour. Technol. 2019, 289, 121732. [Google Scholar] [CrossRef]
- Sánchez-Zurano, A.; Ciardi, M.; Lafarga, T.; Fernández-Sevilla, J.M.; Bermejo, R.; Molina-Grima, E. Role of Microalgae in the Recovery of Nutrients from Pig Manure. Processes 2021, 9, 11. [Google Scholar] [CrossRef]
- Siddiki, S.Y.A.; Mofijur, M.; Kumar, P.S.; Ahmed, S.F.; Inayat, A.; Kusumo, F.; Badruddin, I.A.; Khan, T.M.Y.; Nghiem, L.D.; Ong, H.C.; et al. Microalgae Biomass as a Sustainable Source for Biofuel, Biochemical and Biobased Value-Added Products: An Integrated Biorefinery Concept. Fuel 2022, 307, 121782. [Google Scholar] [CrossRef]
- He, Q.; Yang, H.; Wu, L.; Hu, C. Effect of Light Intensity on Physiological Changes, Carbon Allocation and Neutral Lipid Accumulation in Oleaginous Microalgae. Bioresour. Technol. 2015, 191, 219–228. [Google Scholar] [CrossRef]
- Wang, J.H.; Zhuang, L.L.; Xu, X.Q.; Deantes-Espinosa, V.M.; Wang, X.X.; Hu, H.Y. Microalgal Attachment and Attached Systems for Biomass Production and Wastewater Treatment. Renew. Sustain. Energy Rev. 2018, 92, 331–342. [Google Scholar] [CrossRef]
- Maniam, G.P.; Hindryawati, N.; Nurfitri, I.; Jose, R.; Mohd, M.H.; Dahalan, F.A.; Yusoff, M.M. Decanter Cake as a Feedstock for Biodiesel Production: A First Report. Energy Convers. Manag. 2013, 76, 527–532. [Google Scholar] [CrossRef]
- Abomohra, A.E.F.; Zheng, X.; Wang, Q.; Huang, J.; Ebaid, R. Enhancement of Biodiesel Yield and Characteristics through In-Situ Solvo-Thermal Co-Transesterification of Wet Microalgae with Spent Coffee Grounds. Bioresour. Technol. 2021, 323, 124640. [Google Scholar] [CrossRef] [PubMed]
- Lin-Lan, Z.; Jing-Han, W.; Hong-Ying, H. Differences between Attached and Suspended Microalgal Cells in SsPBR from the Perspective of Physiological Properties. J. Photochem. Photobiol. B Biol. 2018, 181, 164–169. [Google Scholar] [CrossRef]
- Zou, X.; Xu, K.; Chang, W.; Qu, Y.; Li, Y. A Novel Microalgal Biofilm Reactor Using Walnut Shell as Substratum for Microalgae Biofilm Cultivation and Lipid Accumulation. Renew. Energy 2021, 175, 676–685. [Google Scholar] [CrossRef]
- Yu, H.; Zhuang, L.L.; Zhang, M.; Zhang, J. The Mechanism Study of Attached Microalgae Cultivation Based on Reverse Osmosis Concentrated Water (WROC). Resour. Conserv. Recycl. 2021, 179, 106066. [Google Scholar] [CrossRef]
- Palma, H.; Killoran, E.; Sheehan, M.; Berner, F.; Heimann, K. Assessment of Microalga Biofilms for Simultaneous Remediation and Biofuel Generation in Mine Tailings Water. Bioresour. Technol. 2017, 234, 327–335. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, H.; Guan, L.; Wang, X.; Wang, Y.; Jiang, Z.; Cao, L.; Zhang, X. Influence of Photoperiods on Microalgae Biofilm: Photosynthetic Performance, Biomass Yield, and Cellular Composition. Energies 2019, 12, 3724. [Google Scholar] [CrossRef]
- Valchev, D.; Ribarova, I. A Review on the Reliability and the Readiness Level of Microalgae-Based Nutrient Recovery Technologies for Secondary Treated Effluent in Municipal Wastewater Treatment Plants. Processes 2022, 10, 399. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernández, C.M.; Casas, A.; Rodríguez, L.; Pérez, Á. Influence of Fatty Acid Composition of Raw Materials on Biodiesel Properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef]
- Miraboutalebi, S.M.; Kazemi, P.; Bahrami, P. Fatty Acid Methyl Ester (FAME) Composition Used for Estimation of Biodiesel Cetane Number Employing Random Forest and Artificial Neural Networks: A New Approach. Fuel 2016, 166, 143–151. [Google Scholar] [CrossRef]
- Folayan, A.J.; Anawe, P.A.L.; Aladejare, A.E.; Ayeni, A.O. Experimental Investigation of the Effect of Fatty Acids Configuration, Chain Length, Branching and Degree of Unsaturation on Biodiesel Fuel Properties Obtained from Lauric Oils, High-Oleic and High-Linoleic Vegetable Oil Biomass. Energy Rep. 2019, 5, 793–806. [Google Scholar] [CrossRef]
- Rosli, S.S.; Wong, C.Y.; Yunus, N.M.; Lam, M.K.; Show, P.L.; Cheng, C.K.; Wang, D.K.; da Oh, W.; Lim, J.W. Optimum Interaction of Light Intensity and CO2 Concentration in Bioremediating N-Rich Real Wastewater via Assimilation into Attached Microalgal Biomass as the Feedstock for Biodiesel Production. Process Saf. Environ. Prot. 2020, 141, 355–365. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, X.; Guo, D.; Ye, T.; Xiong, M.; Zhu, L.; Liu, C.; Jin, S.; Hu, Z. Operation of a Vertical Algal Biofilm Enhanced Raceway Pond for Nutrient Removal and Microalgae-Based Byproducts Production under Different Wastewater Loadings. Bioresour. Technol. 2018, 253, 323–332. [Google Scholar] [CrossRef]
- Borges, L.; Morón-Villarreyes, J.A.; D’Oca, M.G.M.; Abreu, P.C. Effects of Flocculants on Lipid Extraction and Fatty Acid Composition of the Microalgae Nannochloropsis Oculata and Thalassiosira Weissflogii. Biomass Bioenergy 2011, 35, 4449–4454. [Google Scholar] [CrossRef]
- Gross, M.; Henry, W.; Michael, C.; Wen, Z. Development of a Rotating Algal Biofilm Growth System for Attached Microalgae Growth with in Situ Biomass Harvest. Bioresour. Technol. 2013, 150, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Tizvir, A.; Shojaeefard, M.H.; Zahedi, A.; Molaeimanesh, G.R. Performance and Emission Characteristics of Biodiesel Fuel from Dunaliella Tertiolecta Microalgae. Renew. Energy 2022, 182, 552–561. [Google Scholar] [CrossRef]
- Kim, S.; Moon, M.; Kwak, M.; Lee, B.; Chang, Y.K. Statistical Optimization of Light Intensity and CO2 Concentration for Lipid Production Derived from Attached Cultivation of Green Microalga ettlia Sp. Sci. Rep. 2018, 8, 15390. [Google Scholar] [CrossRef] [PubMed]
- Fakhry, E.M.; Maghraby, D.M. Fatty Acids Composition and Biodiesel Characterization of Dunaliella salina. J. Water Resour. Prot. 2013, 05, 894–899. [Google Scholar] [CrossRef]
- Santhana Kumar, V.; das Sarkar, S.; Das, B.K.; Sarkar, D.J.; Gogoi, P.; Maurye, P.; Mitra, T.; Talukder, A.K.; Ganguly, S.; Nag, S.K.; et al. Sustainable Biodiesel Production from Microalgae Graesiella Emersonii through Valorization of Garden Wastes-Based Vermicompost. Sci. Total Environ. 2022, 807, 150995. [Google Scholar] [CrossRef] [PubMed]
- Leong, W.H.; Azella Zaine, S.N.; Ho, Y.C.; Uemura, Y.; Lam, M.K.; Khoo, K.S.; Kiatkittipong, W.; Cheng, C.K.; Show, P.L.; Lim, J.W. Impact of Various Microalgal-Bacterial Populations on Municipal Wastewater Bioremediation and Its Energy Feasibility for Lipid-Based Biofuel Production. J. Environ. Manag. 2019, 249, 109384. [Google Scholar] [CrossRef] [PubMed]
- Ryckaert, W.R.; Smet, K.A.G.; Roelandts, I.A.A.; van Gils, M.; Hanselaer, P. Linear LED Tubes versus Fluorescent Lamps: An Evaluation. Energy Build. 2012, 49, 429–436. [Google Scholar] [CrossRef]
- Huang, R.; Li, J.; Tang, Y.; Song, W.; Yu, Y.; Yang, W.; Cheng, J. Comparative Life-Cycle Assessment of Microalgal Biodiesel Production via Various Emerging Wet Scenarios: Energy Conversion Characteristics and Environmental Impacts. Energy Convers. Manag. 2022, 257, 115427. [Google Scholar] [CrossRef]
- Razon, L.F.; Tan, R.R. Net Energy Analysis of the Production of Biodiesel and Biogas from the Microalgae: Haematococcus Pluvialis and Nannochloropsis. Appl. Energy 2011, 88, 3507–3514. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Visible Spectrum | Intensity (µmol/m2 s) | Microalgal Species | Stimulating Effect in Decreasing Order | Reference |
|---|---|---|---|---|
| White, red, blue, green, yellow | 100 | Chlorella vulgaris | White > yellow > blue > red > green | Current study |
| White, red, blue, green | 60 | Botryococcus braunii | Red > white > blue > green | [23] |
| White, red, blue, green, yellow | 100 | Chlorella vulgaris | White > red > blue > yellow > green | [22] |
| Diacronema lutheri | Blue > red > green > yellow > white | |||
| Porphyridium purpureum | Green > blue > yellow > red > white | |||
| White, red, blue, green, yellow | - | Chlorella vulgaris | White > red > blue > yellow > green | [23] |
| - | Tetradesmus obliquus | Red > blue > white > yellow > green | ||
| - | Arthospira platensis | Red > white > blue > yellow > green | ||
| White, red, blue, | - | Auxenochlorella pyrenoidosa, Scenedesmus quadricaud, Tetradesmus obliquus | Blue > red > white | [24] |
| White, red, yellow | - | Phaeodactylum tricornutum | Red > white > yellow | [24] |
| White, red, blue | 52 | Isochrysis galbana | Blue > red > white | [25] |
| White, red, blue, green | 100 | Chlorella sp., Nannochloris oculata | Blue > red > white > green | [5] |
| Visible Spectrum | Intensity (µmol/m2 s) | Microalgal Species | Stimulating Effect in Decreasing Order | Reference |
|---|---|---|---|---|
| White, red, blue, green, yellow | 100 | Chlorella vulgaris | Green > red > yellow > white > blue | Current study |
| White, red, blue, green | 60 | Botryococcus braunii | Red > white > blue > green | [23] |
| White, red, blue, green, yellow | - | Chlorella vulgaris | Blue > red > white > green > yellow | [23] |
| - | Tetradesmus obliquus | Blue > red > white > green > yellow | ||
| - | Arthospira platensis | Blue > red > white > green > yellow | ||
| White, red, yellow | - | Phaeodactylum tricornutum | Red > white | [24] |
| White, red, blue, green | 100 | Chlorella sp. | Blue > red > white > green | [5] |
| Nannochloris oculata | Blue > white > red > green | |||
| White | 100 | Nannochloropsis sp. | Blue > white > blue-green > pink > red | [31] |
| Red | 75 | |||
| Blue | 15 | |||
| Blue-green | 20 | |||
| Pink | 85 | |||
| White, red, blue | 70 | Streptomyces. bacillaris | Red > white > blue | [32] |
| White, red, blue | 50 | Chlorella vulgaris | Blue > white > red | [30] |
| White, red, blue | 104 | Isochrysis galbana | Blue > white > red | [25] |
| Supporting Material | Microalgal Species | Protein Content (%) | Carbohydrate Content (%) | Reference |
|---|---|---|---|---|
| Palm decanter cake | Chlorella vulgaris | 34.10 ± 0.65 | 60.91 ± 1.15 | Current study |
| Spent coffee grounds | Chlorella pyremoidosa | 20.36 ± 1.13 | 20.69 ± 0.89 | [38] |
| Cottom, linen, mohair | Scenedesmus. sp | 50.10 ± 10.1 | 37.60 ± 10.5 | [39] |
| Walnut shells | Chlorella vulgaris | 27.70 ± 1.25 | 34.50 ± 1.31 | [40] |
| Scenedesmus obliquus | 34.80 ± 0.55 | 32.50 ± 0.59 | ||
| Cellulose acetate membrane | Chlorella vulgaris | 52.84 ± 2.50 | 37.93 ± 1.15 | [41] |
| Glass slide | Consortia | 12.00 ± 1.13 | 60.00 ± 2.07 | [42] |
| Cellulose acetate–nitrate membrane | Nannochloris oculata | 19.20 ± 2.59 | 9.10 ± 0.49 | [43] |
| Chlorella sp. | 19.20 ± 2.57 | 3.70 ± 0.24 | ||
| Chlorella pyrenoidosa | 31.00 ± 2.01 | 9.00 ± 0.50 |
| FAME Species | FAME Content (%) | Saturation Degree |
|---|---|---|
| Methyl palmitate (C 16:0) | 51.36 | SFA |
| Methyl stearate (C18:0) | 5.36 | SFA |
| Methyl lingnocerate (C24:0) | 13.66 | SFA |
| cis-Methyl oleate and trans-Methyl 9-octadecenoate (C18:1) | 25.97 | MUFA |
| Methyl cis-11,14,17-eicosatrienoate (C20:3) | 1.88 | PUFA |
| Methyl cis-5,8,11,14,17-eicosapentaenoate (C20:5) | 1.77 | PUFA |
| Microalgal Species | Supporting Material/ Culture Medium | Total C16:0, C18:0, C18:1, C18:2, C18:3 (%) | SFA (%) | MUFA (%) | PUFA (%) | Reference |
|---|---|---|---|---|---|---|
| Chlorella vulgaris | Palm decanter cake | 82.69 ± 1.40 | 70.38 ± 2.30 | 25.97 ± 1.12 | 3.65 ± 0.28 | Current study |
| Chlorella vulgaris, Oscillatoria tenus, Scenedesmus obliquus | Wastewater | 78.67 | 61.57 | 20.23 | 11.92 | [49] |
| Nannochloropsis oculata | - | 44.00 ± 4.30 | 43.03 ± 2.20 | 37.41 ± 3.40 | 15.12 ± 1.30 | [50] |
| Thalassiosira weissflogii | 29.69 ± 3.55 | 26.52 ± 4.65 | 34.25 ± 5.00 | 36.95 ± 4.50 | ||
| Chlorella vulgaris | Polyurethane foam | 68.70 | 35.60 | 43.10 | 19.70 | [3] |
| Chlorella vulgaris, | Walnut shells | 77.85 ± 5.05 | 32.60 ± 3.96 | 2.85 ± 0.11 | 58.66 ± 5.67 | [40] |
| Scenedesmus obliquus | 78.67 ± 5.80 | 25.99 ± 2.53 | 15.34 ± 1.60 | 53.66 ± 4.81 | ||
| Chlorella vulgaris | Cotton | 92.89 ± 0.93 | 32.37 ± 0.39 | 28.04 ± 0.40 | 39.57 ± 0.34 | [51] |
| Dunaliella tertiolecta | - | 97.8 | 32.17 | 19.45 | 47.25 | [52] |
| Phaeodactylum tricornutum | - | 18.69 ± 3.62 | 25.77 ± 3.04 | 31.65 ± 1.94 | 29.85 ± 5.18 | [9] |
| Ettlia sp. | Porous membrane | 90.10 | 23.60 | 29.20 | 43.50 | [53] |
| Dunaliella salina | 53.33 | 34.76 | 32.18 | 33.06 | [54] | |
| Graesiella emersonni | Vermicompost | 82.70 ± 5.49 | 47.10 ± 2.43 | 15.30 ± 1.01 | 37.60 ± 1.66 | [55] |
| Microalgae Species | Cultivation System | NER | Reference |
|---|---|---|---|
| Chlorella vulgaris | Attached | 0.30 | Current study |
| Chlorella vulgaris | Suspended | 0.27 | [56] |
| Consortium | Suspended | 0.73 | [58] |
| Haematococcus pluvialis | Suspended | 0.40 | [59] |
| Nannochloropsis sp. | 0.12 |
| Process | Amount | Energy Equivalent (MJ) |
|---|---|---|
| Cultivation: | ||
| Water | 0.9 kg | 5.25 |
| Electricity | 0.89 kWh | 3.20 |
| Harvesting: | ||
| Electricity | 26.36 kWh | 94.90 |
| Lipid extraction: | ||
| Electricity | 3.435 kWh | 12.37 |
| Total cumulative energy demand | - | 115.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiong, Z.W.; Rawindran, H.; Leong, W.H.; Liew, C.S.; Wong, Y.Y.; Kiatkittipong, W.; Abdelfattah, E.A.; Show, P.L.; Rahmah, A.U.; Tong, W.Y.; et al. Impact of Various Visible Spectra on Attached Microalgal Growth on Palm Decanter Cake in Triggering Protein, Carbohydrate, and Lipid to Biodiesel Production. Processes 2022, 10, 1583. https://doi.org/10.3390/pr10081583
Tiong ZW, Rawindran H, Leong WH, Liew CS, Wong YY, Kiatkittipong W, Abdelfattah EA, Show PL, Rahmah AU, Tong WY, et al. Impact of Various Visible Spectra on Attached Microalgal Growth on Palm Decanter Cake in Triggering Protein, Carbohydrate, and Lipid to Biodiesel Production. Processes. 2022; 10(8):1583. https://doi.org/10.3390/pr10081583
Chicago/Turabian StyleTiong, Zhi Wei, Hemamalini Rawindran, Wai Hong Leong, Chin Seng Liew, Yi Ying Wong, Worapon Kiatkittipong, Eman Alaaeldin Abdelfattah, Pau Loke Show, Anisa Ur Rahmah, Woei Yenn Tong, and et al. 2022. "Impact of Various Visible Spectra on Attached Microalgal Growth on Palm Decanter Cake in Triggering Protein, Carbohydrate, and Lipid to Biodiesel Production" Processes 10, no. 8: 1583. https://doi.org/10.3390/pr10081583
APA StyleTiong, Z. W., Rawindran, H., Leong, W. H., Liew, C. S., Wong, Y. Y., Kiatkittipong, W., Abdelfattah, E. A., Show, P. L., Rahmah, A. U., Tong, W. Y., & Lim, J. W. (2022). Impact of Various Visible Spectra on Attached Microalgal Growth on Palm Decanter Cake in Triggering Protein, Carbohydrate, and Lipid to Biodiesel Production. Processes, 10(8), 1583. https://doi.org/10.3390/pr10081583