1. Introduction
The plastid terminal oxidase (PTOX) is a crucial enzyme located within the plastids of plant cells. It functions as a plastid quinone oxidase, pivotal in metabolic pathways that contribute to carotenoid synthesis, photorespiration, and chloroplast respiration [
1]. The gene encoding PTOX is nuclear-encoded, and its protein product is crucial for maintaining the redox balance within the plastid. The
Arabidopsis thaliana leaf white spot mutant was used to facilitate the initial characterization of PTOX [
2]. PTOX regulates the balance between photosynthesis and cellular respiration by modulating electron transfer pathways [
3]. Under optimal light conditions, photosynthesis is the primary energy source in plant cells, using linear electron flow (LEF) to generate ATP and NADPH. However, in situations like high light intensity or temperature stress, LEF can result in excess electrons leading to photo-oxidative damage, and this is where PTOX plays a crucial role. In stressful environments, it diverts excess electrons from linear electron flow (LEF) to cyclic electron flow (CEF), reducing the risk of photo-oxidative damage [
4]. This balance allows plants to adapt to varying conditions, ensuring energy supply while safeguarding against damage. PTOX acts as a crucial regulator, enabling the efficient utilization of light energy and maintaining the overall stability of energy metabolism in plants [
5].
Chlamydomonas reinhardtii is a unicellular green alga that has been extensively studied for its ability to grow rapidly and produce high amounts of biomass rich in proteins and other nutrients [
6]. This makes it a promising candidate for applications in active substance production or protein production for various purposes. In terms of stressful conditions, challenges such as high light intensity, extreme temperatures, nutrient limitations, and oxidative stress can impact the growth, photosynthetic efficiency, and overall productivity of
C. reinhardtii [
7]. Studying the PTOX in
C. reinhardtii holds significant importance due to the potential insights it can provide into the alga’s adaptive mechanisms and its ability to thrive under stressful conditions. In contrast to higher plants, which usually have only one form of ptox, the green alga
C. reinhardtii has two isoforms,
ptox1 and
ptox2 [
8]. PTOX1 is mainly responsible for regenerating plastoquinone (PQ), which is necessary for desaturation reactions during the biosynthesis of phytoene, an early step in the carotenoid biosynthetic pathway [
4]. PTOX1 plays a crucial role in carotenoid production, including astaxanthin, which provides photoprotection and helps the alga cope with high light stress [
9]. In contrast, PTOX2 exhibits a higher rate of plastoquinol oxidation during photosynthesis than PTOX1 [
10]. Plastoquinol is the reduced form of PQ. Its oxidation by PTOX2 helps to maintain the flow of electrons in the photosynthetic electron transport chain, particularly under conditions where the demand for ATP and NADPH exceeds the consumption by the Calvin cycle. This activity of PTOX2 is important for preventing the over-reduction of the photosynthetic electron transport chain, which could lead to the production of harmful ROS [
11]. The existence of two PTOX isoforms in
C. reinhardtii enables a more nuanced regulation of PQ redox state and carotenoid biosynthesis, providing the alga with a flexible response to various environmental conditions. The differential expression and activity of PTOX1 and PTOX2 allow the alga to balance photosynthetic efficiency, redox homeostasis, and stress acclimation responses. However, no further research shows the interaction between these two
PTOX genes.
In this study, the synergistic effects of plastid terminal oxidases 1 and 2 (PTOX1 and PTOX2) in the regulation of astaxanthin under stress conditions were investigated. The research sheds light on the intricate regulation of astaxanthin biosynthesis and photosynthetic efficiency in response to diverse environmental conditions.
2. Methods and Materials
2.1. Organism, Growth Medium, and Culture Conditions
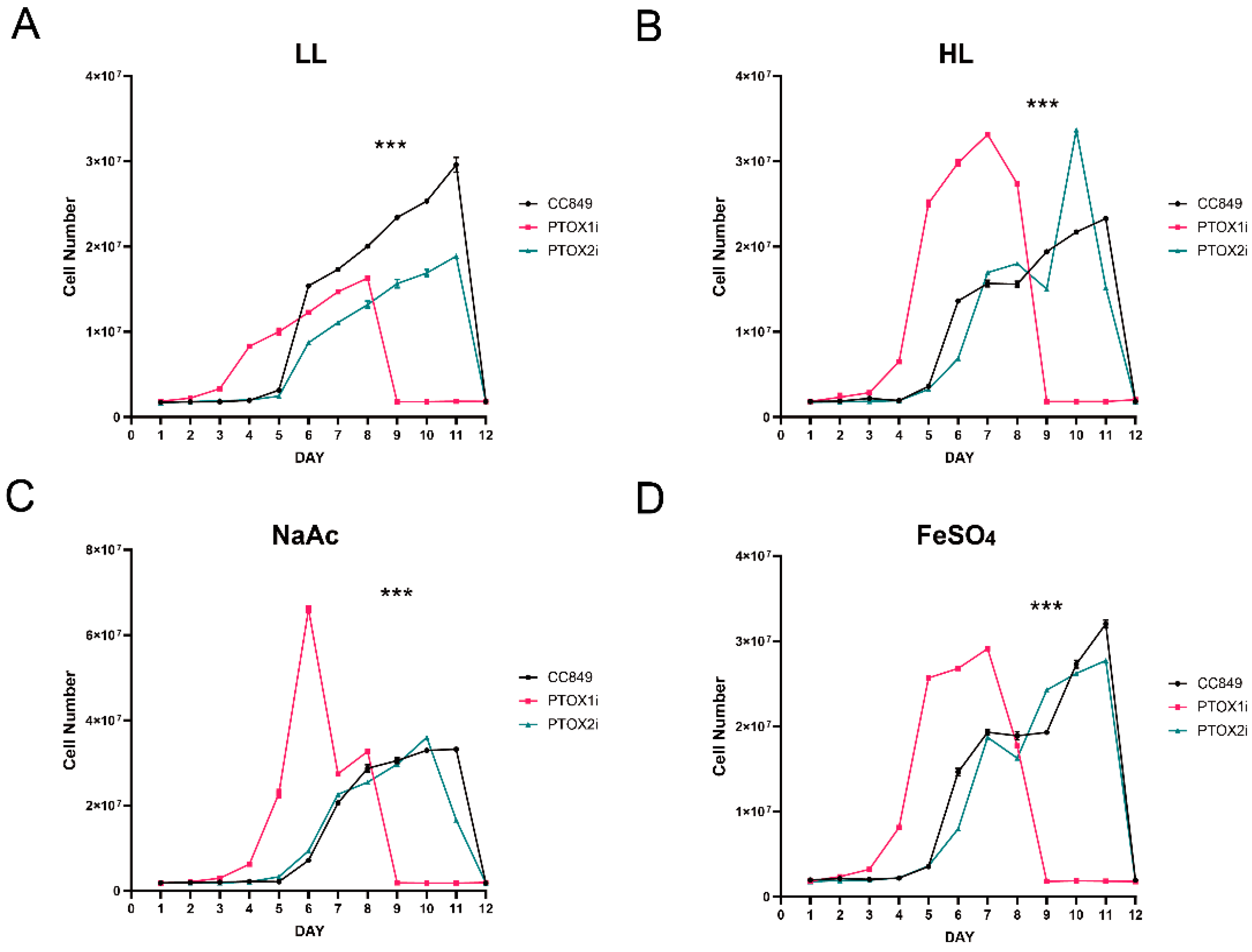
Chlamydomonas reinhardtii cell wall deficient mutant strain CC849 was obtained from the Chlamydomonas Genetic Centre (Duke University, Durham, NC, USA). Microalgal cells were grown using Tris-acetate phosphate (TAP) media with mineral nutrient supplements at 22 ± 1 °C and under continuous cool-white fluorescent lamps (≈20 μmol/m2s). For the analysis of cell growth, C. reinhardtii CC849 and transgenic algal strains are grown under different light conditions: normal light condition (approximately 20 μmol photons/m2s), high light condition (approximately 150 μmol photons/m2s), high light with 450 μM FeSO4 condition, high light with 45 mM NaAC condition, and high light with 450 μM FeSO4 + 45 mM NaAC condition (Macklin Biochemical Technology Co. Ltd., Shanghai, China).
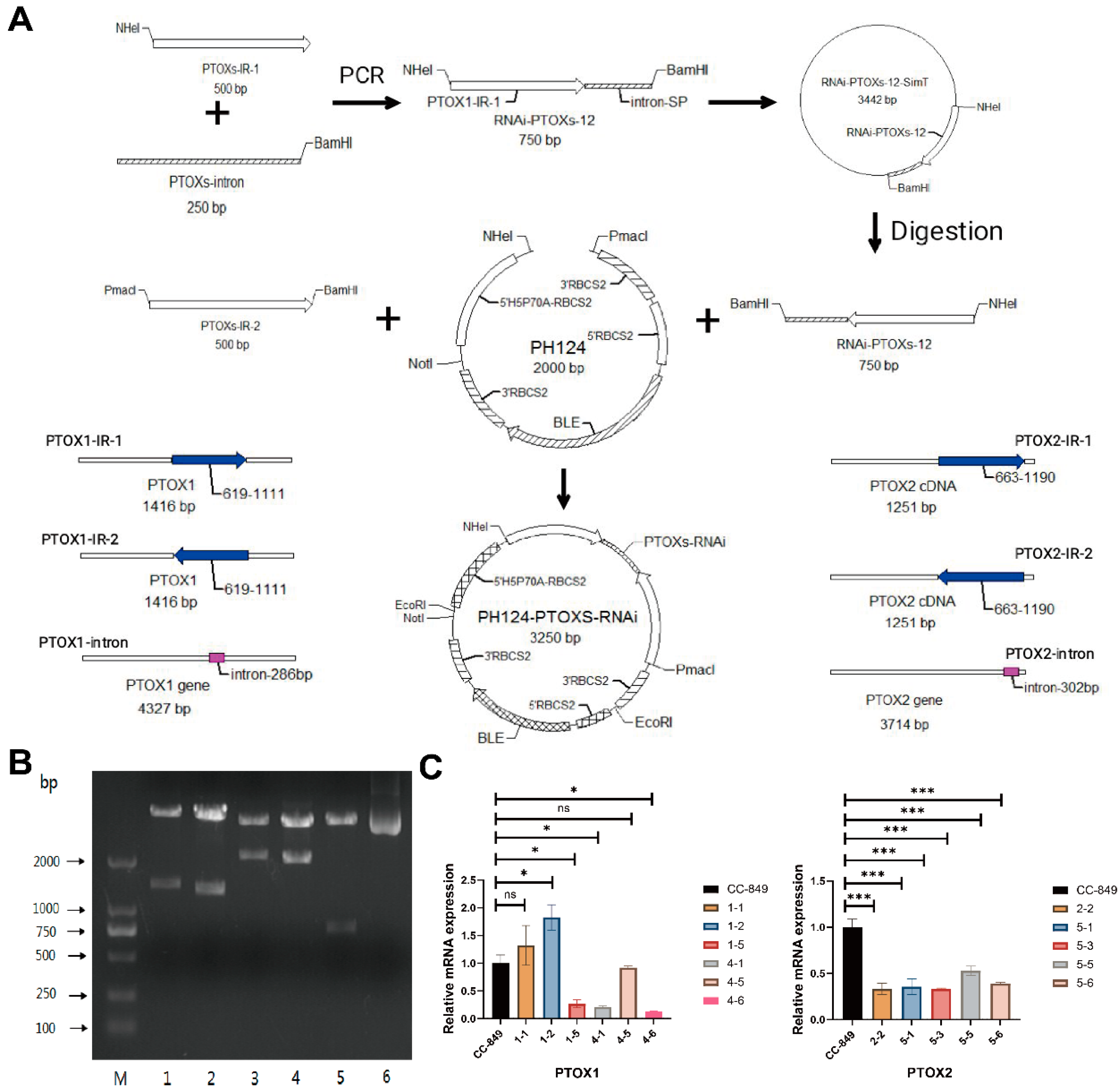
2.2. RNAi Vector Construction and Transformation
For the targeted gene silencing of
ptox1 (GenBank ID: 5718064) and
ptox2 (GenBank ID: 5728910), complementary DNA fragments corresponding to the sense and antisense of the above genes, including their intronic sequences, were amplified using specific primer pairs (Sangon Biotech Co. Ltd., Shanghai, China). The glass bead method was used to transform the plasmid into the UVM4 strain, which is capable of highly efficient expression. In this method,
C. reinhardtii CC849 cells were grown on solid media supplemented with 1.5 mM L-tryptophan, 5 μg/mL paromomycin, and 5 μM 5-fluoroindole (5-FI) (Macklin Biochemical Technology Co. Ltd., Shanghai, China). The coding sequences (CDSs) of
ptox1 and
ptox2 were synthesized and fused to their respective introns. The resulting CDS–intron fusion products were then cloned into the pH124-PTOXs RNAi plasmid vector, as depicted in
Figure 1A. To achieve the knockdown of
ptox1 and
ptox2 transcripts, the resulting plasmids were subjected to Not I restriction digestion and introduced into
C. reinhardtii CC849 cells using the glass bead agitation method. Quantitative reverse transcription-PCR (qRT-PCR) was then used to identify the downregulation of
ptox1 and
ptox2 transcript levels via TaKaRa RNA PCR Kit (code number DRR019A) and SYBR PrimeScript
TM RT-PCR Kit (Perfect Real Time, Takara Biotechnology Co. Ltd., Shiga, Japan) performed on an ABI Prism 7300 Sequence Detection System (Applied Biosystems, Waltham, MA, USA). β-actin was used as the internal reference gene. The lowest relative expression ptoxs were selectively amplified for use in downstream experimental procedures. All primers involved in this study were listed in
Table S1. All experiments were performed in triplicate.
2.3. DNA and RNA Isolations
Genomic DNA was isolated from approximately 3 × 107 cells of C. reinhardtii CC849, which were cultured to the log phase, using the Universal DNA Extraction Kit Ver.3.0 (Takara Biotechnology Co. Ltd., Shiga, Japan). RNA isolation from the same strain of C. reinhardtii CC849 was performed using the FAST200 Kit according to the manufacturer’s instructions (FeiJie Shanghai Biotech. Ltd., Shanghai, China). All experiments were performed in triplicate.
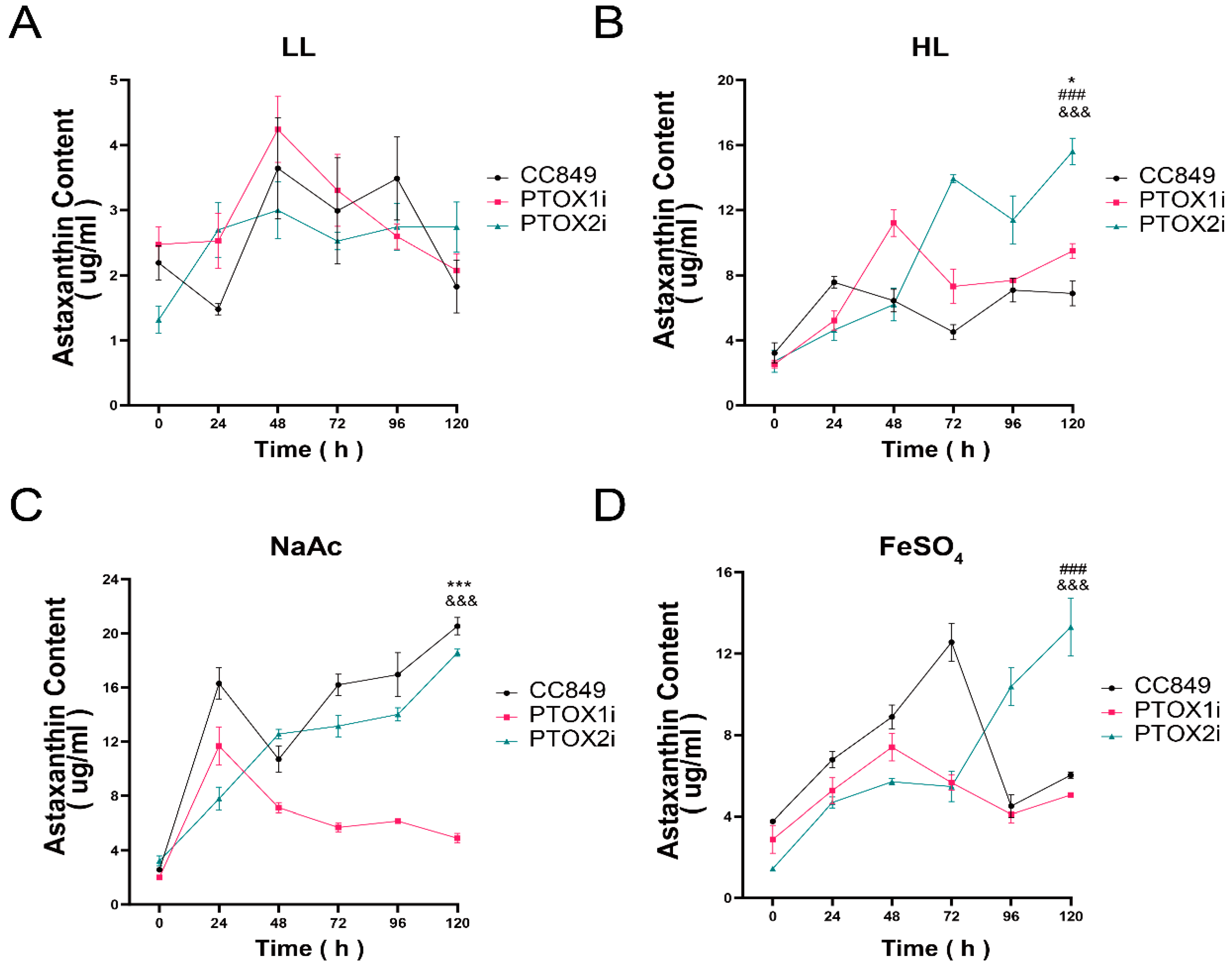
2.4. The Production of Astaxanthin
To analyze the astaxanthin content of C. reinhardtii CC849 and transgenic algal strains, High-Performance Liquid Chromatography (Waters Corp., Milford, MA, USA) was used. The cultures of C. reinhardtii CC849 and transgenic algal strains were grown under different light conditions: normal light condition (approximately 20 μmol photons/m2s), high light condition (approximately 150 μmol photons/m2s), high light with 450 μM FeSO4 condition, high light with 45 mM NaAC condition, and high light with 450 μM FeSO4 + 45 mM NaAC condition. Cells were harvested and the intracellular astaxanthin was extracted by dichloromethane–methanol (v/v = 25:75). The analytical column was a 4.6 × 250 mm HPLC column (Kromasil C18 5 μm, Kromasil Co., Bohus, Sweden). HPLC conditions were as follows: 90/10 methanol/Acetonitrile (v/v); flow rate: 1.0 mL/min; injection volume: 10 μL; and the chromatogram was recorded at 478 nm. All experiments were performed in triplicate.
2.5. Statistical Analysis
Experimental data were expressed as the mean ± standard deviation and analyzed using GraphPad Prism software (version 9.0, GraphPad Software Co. Ltd., Boston, MA. USA) Statistical analyses were conducted using independent sample t-tests for comparisons between two groups and one-way ANOVA for comparisons among three or more groups. A p-value of less than 0.05 was considered statistically significant and indicated as “*” in the bar graphs. The level of significance was further denoted as “*” for p < 0.05, “**” for p < 0.01, “***” for p < 0.001, and “ns” for non-significant results with p > 0.05.
4. Discussions
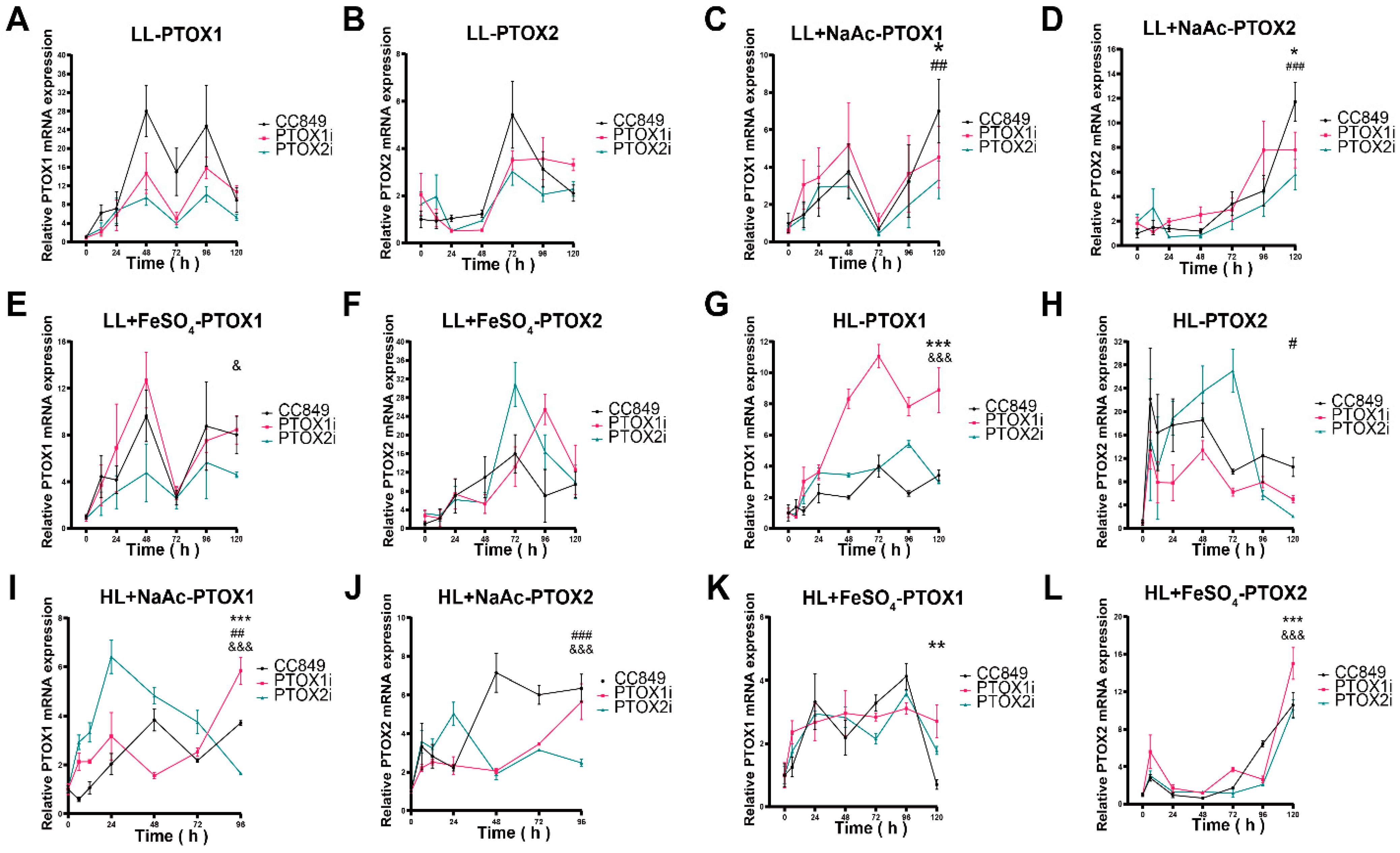
The study shows a significant correlation between the mRNA levels of ptox2 and the alga’s response to high light stress, indicating that ptox2 plays a crucial role in photoprotection under these conditions. Conversely, the behavior of ptox2 under high salt stress suggests an increased level of activity, indicating a role in osmotic stress management. The article demonstrates the relationship between environmental cues and gene expression regulation in C. reinhardtii through the responses of ptox2 to various stressors. Additionally, it clarifies the distinct functions of ptox1 and ptox2 in the alga’s stress response mechanisms, providing new insights into the regulatory processes that control carotenoid biosynthesis and photosynthetic efficiency. The discovery that ptox1 acts as a positive regulator of astaxanthin accumulation, while ptox2 functions as a counter-regulatory gene, enhances our understanding of how this model organism adapts to changing environmental conditions. These findings provide insight into the molecular mechanisms underlying stress responses and metabolic regulation in photosynthetic organisms, contributing to the broader field of plant biology.
The C-terminal region of PTOX exhibits a high degree of sequence homology with the mitochondrial alternative oxidase (AOX) [
12]. While PTOX localizes in the plastid, AOX localizes in the mitochondrion [
13]. Both PTOX and AOX share a significant feature, a hydrophobic domain, which is crucial for their subcellular localization within their respective organelles [
14]. AOX plays a role in bypassing cytochrome c oxidase in the mitochondrion under certain conditions. PTOX in the plastid helps to maintain cellular energy homeostasis and minimize the production of ROS [
15]. Additionally, it manages the redox status of the photosynthetic electron transport chain. In situations such as high light intensity or limited CO₂ availability, it can accept electrons from the PQ pool, preventing over-reduction and potentially harmful ROS production. The shared hydrophobic motif between PTOX and AOX highlights their relatedness and emphasizes their critical roles in modulating electron flow within their respective organelles [
16]. This contributes to the overall cellular redox balance and stress response mechanisms.
Knocking out ptox in plants and microalgae results in severe phenotypes, such as developmental and growth defects, as well as increased photosensitivity. This indicates that ptox plays a crucial role in maintaining normal growth and protecting against light-induced damage. However, downregulating ptox to approximately 3% of the wild-type levels did not affect plant growth under standard laboratory conditions in
Arabidopsis [
17]. Plants exhibit plasticity in response to reduced ptox expression, potentially due to compensatory mechanisms or alternative pathways for managing photosynthetic electron flux. Studies on ptox overexpression have revealed its function. For example, the overexpression of
ptox1 in
Chlamydomonas reinhardtii leads to mutants that are more sensitive to high light stress than wild-type plants [
18,
19]. Similarly, the overexpression of
Arabidopsis ptox in tobacco has been observed to increase susceptibility to oxidative stress [
20,
21]. These findings suggest that while ptox can enhance the ability to handle excess light energy, overexpression can also have detrimental effects, possibly due to an imbalance in the redox state of the photosynthetic electron transport chain. In the case of the salt-tolerant brassica species
Eutrema salsugineum, the overexpression of ptox leads to faster induction and higher tolerance upon exposure to salt stress [
22]. This suggests that ptox may play a role in mitigating the effects of salt stress, possibly by dissipating excess reducing equivalents generated during stress conditions. Moreover, the expression of OsPTOX in the cyanobacterium
Synechocystis does not affect growth under standard conditions with light intensities ranging from 50 to 150 μmol photons/m
2s. This implies that under non-stressful conditions, the modulation of ptox expression may not significantly impact the growth of this organism [
23]. These findings emphasize the significance of finely regulating ptox expression to maintain photosynthetic efficiency and protect against environmental stresses. The diverse responses observed in various organisms and under different conditions highlight the importance of a nuanced comprehension of ptox’s functions in photosynthetic organisms and its potential for enhancing stress tolerance in crops.
Under high light stress, the activation of the light promoter in the pH124 vector inhibited both the ptox1 and ptox2 genes. The increase in the expression of one gene when the other is inhibited suggests a compensatory mechanism between the two, indicating that PTOX1 and PTOX2 may have overlapping functions and can substitute for each other to some extent. The observations suggest that the PTOX genes play a more significant role in response to high light stress than NaAc-induced stress. This is because the ptox1 and ptox2 genes were not significantly inhibited in their respective silenced strains compared to the high-light intensity group. The lack of a significant complementary effect between PTOX1 and PTOX2 under NaAc treatment suggests that these two genes may have similar functions in response to this particular stressor. Inhibiting either gene does not result in a compensatory upregulation of the other gene. Additionally, the experimental group exhibited a reduced silencing effect when exposed to the combined stress of FeSO4 and high light intensity. The statement suggests that the inclusion of FeSO4 as a stressor exacerbates the condition of the cells, necessitating an upregulation of both PTOX genes to cope with the excess reducing equivalents produced during photosynthesis.
The responses of PTOX genes to different stress conditions highlight the intricate regulatory mechanisms involved in maintaining photosynthetic efficiency and cellular homeostasis. These findings underscore the significance of PTOX in mitigating the detrimental effects of environmental stressors, particularly under high light intensity. Exploring the potential of PTOX in responding to diverse stressors opens up promising avenues for future research and possible applications. Future research directions could delve into further elucidating the specific stress-response pathways and signaling networks involving PTOX genes. Investigating the molecular mechanisms underlying the upregulation or downregulation of PTOX expression under different stress conditions would provide valuable insights into their adaptive roles and regulatory mechanisms. Additionally, exploring the genetic diversity and functional variations of PTOX genes in different plant species could enhance our understanding of their evolutionary significance and potential applications. Comparative genomics and population studies may uncover novel PTOX variants with enhanced stress tolerance or superior photosynthetic performance, paving the way for targeted breeding strategies to incorporate these traits into crop plants. Moreover, the potential of PTOX in stress tolerance enhancement could be harnessed through genetic engineering approaches. Modulating PTOX expression levels or engineering PTOX variants with optimized activity under specific stress conditions could confer improved stress tolerance in photosynthetic organisms. These efforts could involve the use of gene editing technologies, such as CRISPR-Cas9, to precisely modify PTOX genes and assess the resulting phenotypic effects.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}



