
In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents
 , ,
, ,  , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Collection and Identification of the Plant Material
2.3. Extraction and Fractionation of the Total Extract
2.3.1. Helichrysum cymosum
2.3.2. Helichrysum pandurifolium
2.4. Structure Elucidation and Identification of the Isolated Compounds
2.5. Alpha-Glucosidase Assay
2.6. Alpha-Amylase Assay
2.7. Antioxidant Assays
2.7.1. Ferric-Ion Reducing Antioxidant Power (FRAP) Assay
2.7.2. Trolox Equivalent Absorbance Capacity (TEAC) Assay
2.8. Statistical Analysis
2.9. Molecular Docking and Molecular Mechanics-Generalized Born Surface Area (MM-GBSA) Calculations
3. Results
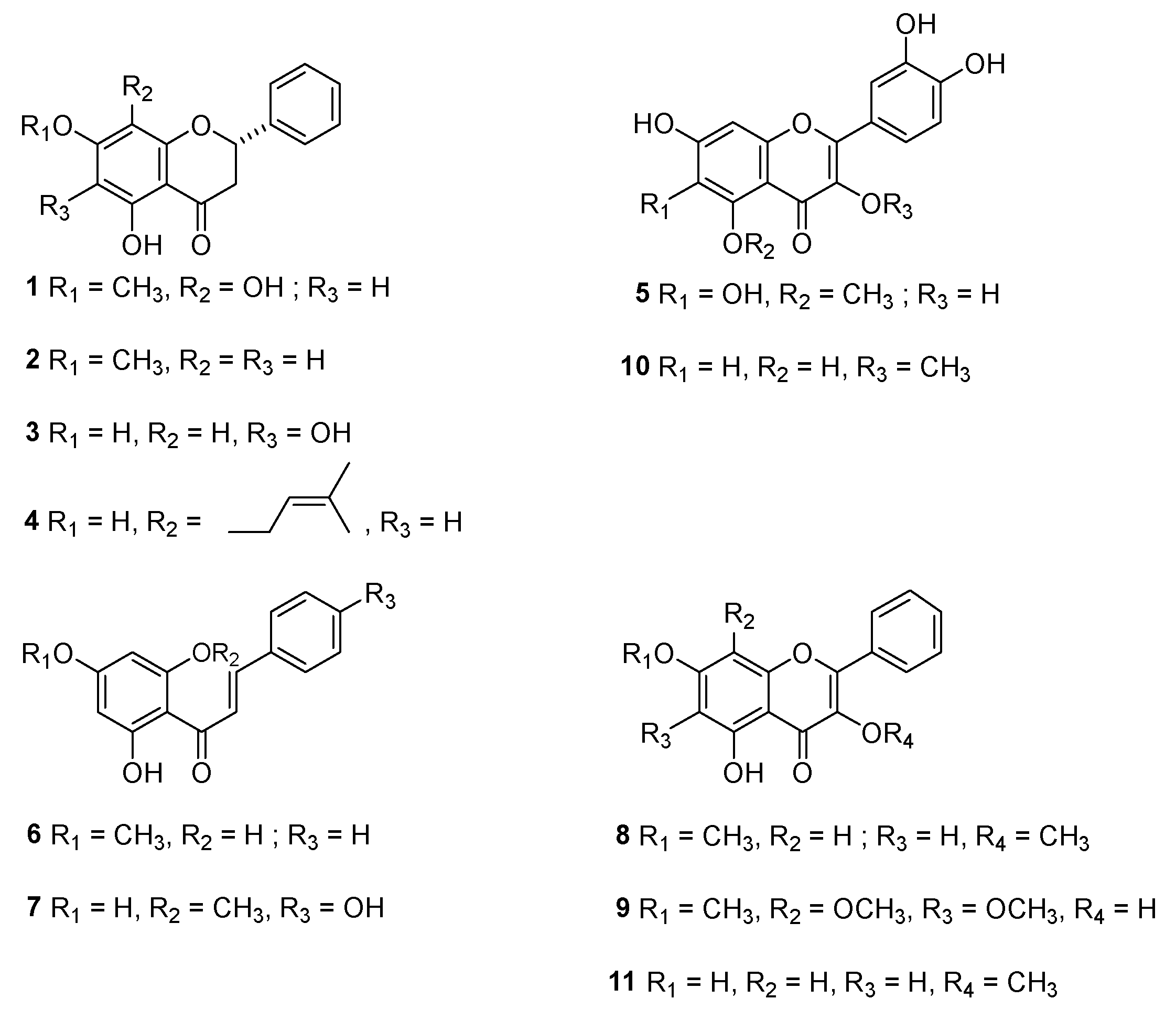
3.1. Isolation and Chemical Characterization of Compound Isolated from H. cymosum and H. pandurifolium
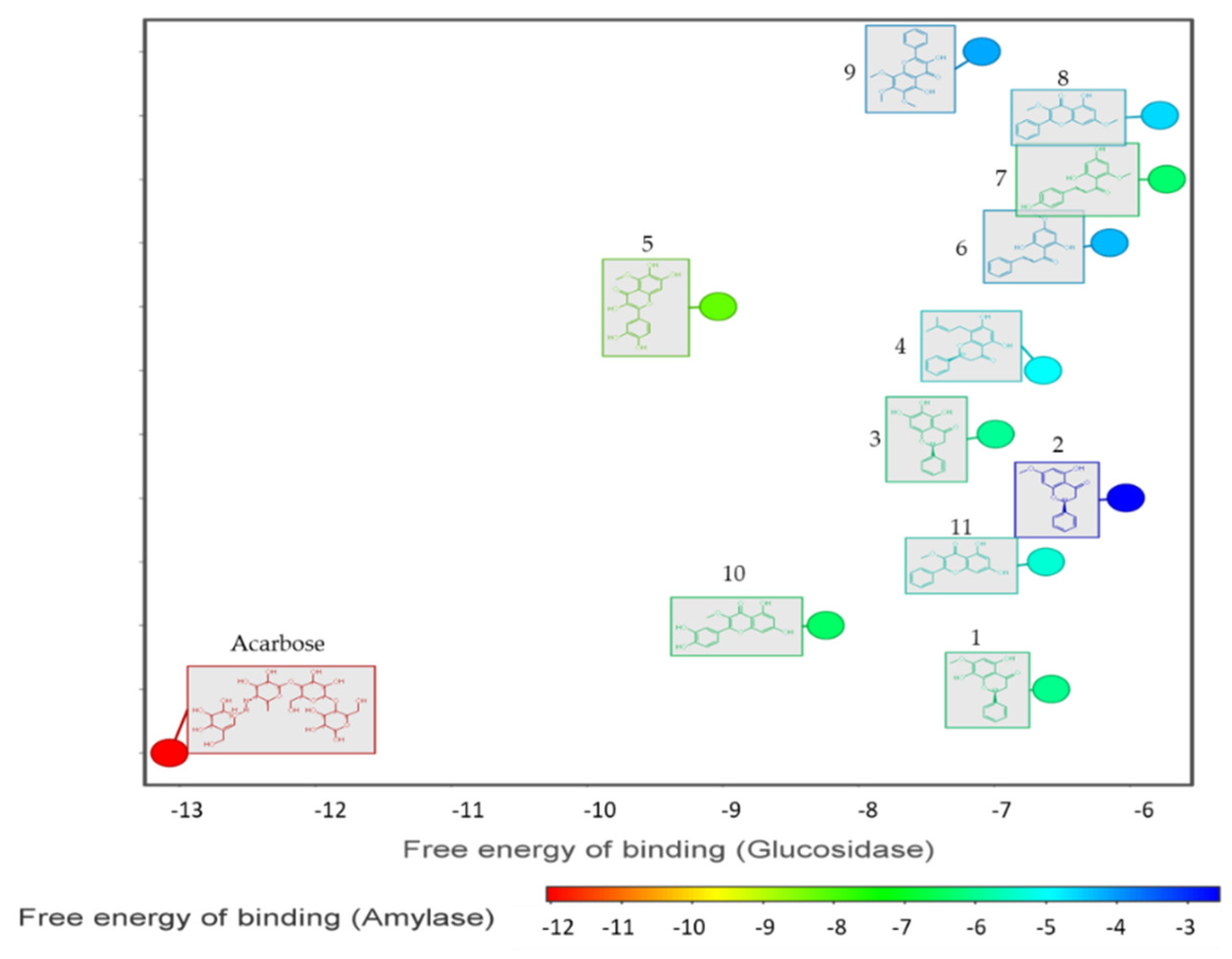
3.2. Alpha-Glucosidase and Alpha-Amylase Activity
3.3. Antioxidant Activity
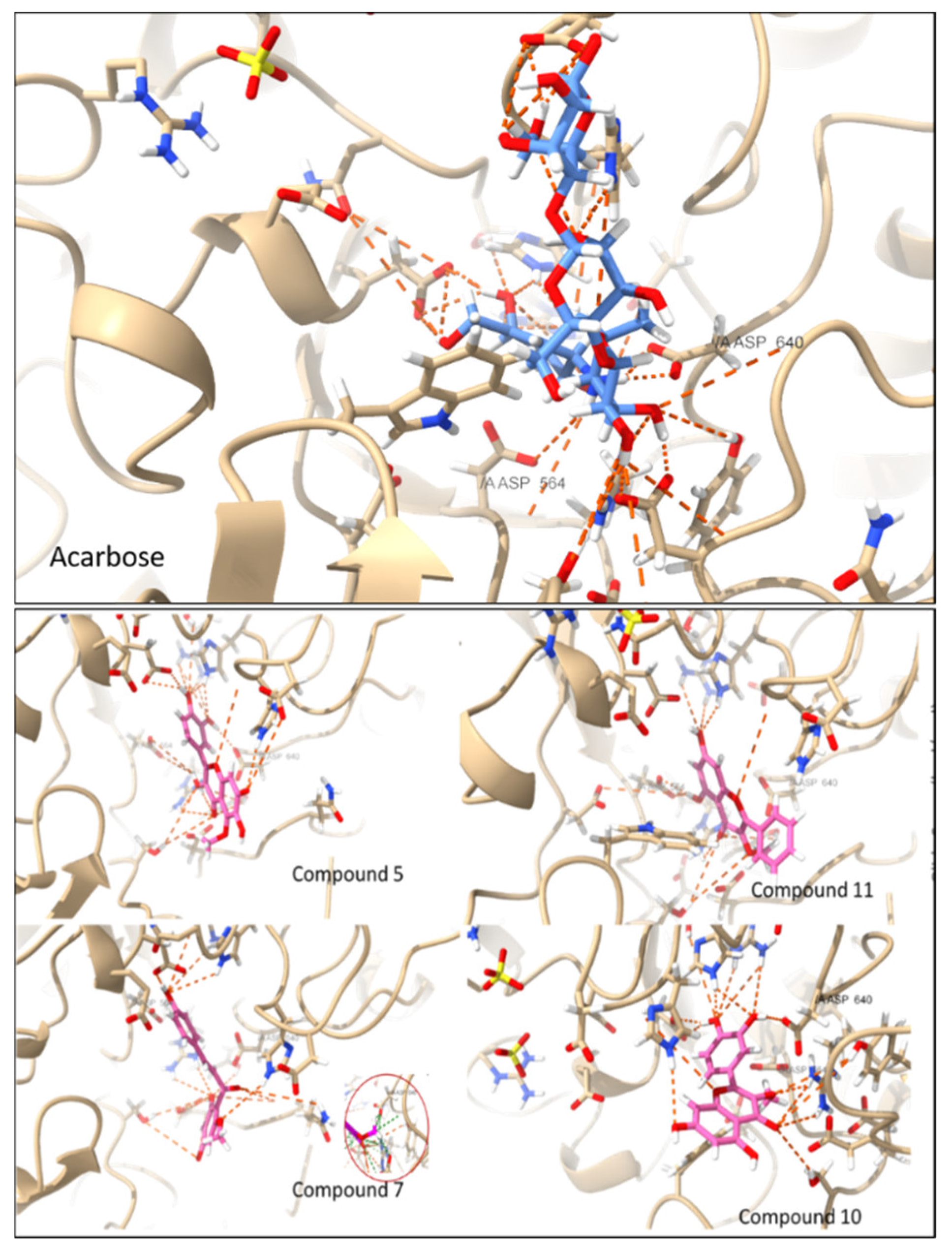
3.4. Molecular Docking
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Astrup, A. Healthy lifestyles in Europe: Prevention of obesity and type II diabetes by diet and physical activity. Public Health Nutr. 2001, 4, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Duckworth, W.C. Hyperglycemia and cardiovascular disease. Curr. Atheroscler. Rep. 2001, 3, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Zheng, C.; Wang, T.; Zhao, H.; Wang, J.; Wang, Z.; Zhai, X.; Jia, Z.; Chen, J.; Zhou, Y.; et al. 1-Deoxynojirimycin: Occurrence, extraction, chemistry, oral pharmacokinetics, biological activities and in silico target fishing. Molecules 2016, 21, 1600. [Google Scholar] [CrossRef] [Green Version]
- Mamun-or-Rashid, A.N.; Hossain, M.S.; Hassan, N.; Dash, B.K.; Sapon, M.A.; Sen, M.K. A review on medicinal plants with antidiabetic activity. J. Pharmacogn. Phytochem. 2014, 3, 149–159. [Google Scholar]
- Ullah, A.; Khan, A.; Khan, I. Diabetes mellitus and oxidative stress—A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical review of antidiabetic drugs: Implications for type 2 diabetes mellitus management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Park, M.; Lee, H.C.; Kang, Y.H.; Kang, E.S.; Kim, S.K. Antidiabetic agents from medicinal plants. Curr. Med. Chem. 2006, 13, 1203–1218. [Google Scholar] [CrossRef] [Green Version]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-glucosidase and α-amylase by flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, D.; Kumar, V.; Sharma, M.; Verma, A. Target guided isolation, in-vitro antidiabetic, antioxidant activity and molecular docking studies of some flavonoids from Albizzia lebbeck Benth. bark. BMC Complement. Altern. Med. 2014, 14, 155. [Google Scholar] [CrossRef] [Green Version]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.; Sousa, J.L.; Tomé, S.M.; Ramos, M.J.; Silva, A.M.; Fernandes, P.A.; Fernandes, E. α-Glucosidase inhibition by flavonoids: An in vitro and in silico structure–activity relationship study. J. Enzyme Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proença, C.; Freitas, M.; Ribeiro, D.; Tomé, S.M.; Oliveira, E.F.; Viegas, M.F.; Araújo, A.N.; Ramos, M.J.; Silva, A.M.; Fernandes, P.A.; et al. Evaluation of a flavonoids library for inhibition of pancreatic α-amylase towards a structure–activity relationship. J. Enzym. Inhib. Med. Chem. 2019, 34, 577–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, G.; Al Harrasi, A.; Hussain, H.; Hamaed, A.; Supuran, C.T. The management of diabetes mellitus-imperative role of natural products against dipeptidyl peptidase-4, α-glucosidase and sodium-dependent glucose co-transporter 2 (SGLT2). Bioorg. Chem. 2019, 86, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Lourens, A.C.; Viljoen, A.M.; Van Heerden, F.R. South African Helichrysum species: A review of the traditional uses, biological activity and phytochemistry. J. Ethnopharmacol. 2008, 119, 630–652. [Google Scholar] [CrossRef]
- Mhlongo, L.S.; Van Wyk, B.E. Zulu medicinal ethnobotany: New records from the Amandawe area of KwaZulu-Natal, South Africa. S. Afr. J. Bot. 2019, 122, 266–290. [Google Scholar] [CrossRef]
- Maroyi, A. Medicinal uses, biological and phytochemical properties of Helichrysum foetidum (L.) Moench (Asteraceae). Asian J. Pharm. Clin. Res. 2019, 12, 13–18. [Google Scholar] [CrossRef]
- Heyman, H.M. Identification of Anti-HIV Compounds in Helichrysum Species (Asteraceae) by Means of NMR-Based Metabolomic Guided Fractionation. Doctoral Dissertation, University of Pretoria, Tswane, South Africa, 2013. [Google Scholar]
- Maroyi, A.L. Helichrysum cymosum (L.) D. Don (Asteraceae): Medicinal uses, chemistry, and biological activities. Asian J. Pharm. Clin. Res. 2019, 12, 19–26. [Google Scholar] [CrossRef]
- Jakupovic, J.; Zdero, C.; Grenz, M.; Tsichritzis, F.; Lehmann, L.; Hashemi-Nejad, S.M.; Bohlmann, F. Twenty-one acylphloroglucinol derivatives and further constituents from South African Helichrysum species. Phytochemistry 1989, 28, 1119–1131. [Google Scholar] [CrossRef]
- Van Vuuren, S.F.; Viljoen, A.M.; Van Zyl, R.L.; Van Heerden, F.R.; Başer, K.H. The antimicrobial, antimalarial and toxicity profiles of helihumulone, leaf essential oil and extracts of Helichrysum cymosum (L.) D. Don subsp. cymosum. S. Afr. J. Bot. 2006, 72, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Popoola, O.K.; Marnewick, J.L.; Rautenbach, F.; Ameer, F.; Iwuoha, E.I.; Hussein, A.A. Inhibition of oxidative stress and skin aging-related enzymes by prenylated chalcones and other flavonoids from Helichrysum teretifolium. Molecules 2015, 20, 7143–7155. [Google Scholar] [CrossRef]
- Lourens, A.C.; Van Vuuren, S.F.; Viljoen, A.M.; Davids, H.; Van Heerden, F.R. Antimicrobial activity and in vitro cytotoxicity of selected South African Helichrysum species. S. Afr. J. Bot. 2011, 77, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Akinyede, K.A.; Cupido, C.N.; Hughes, G.D.; Oguntibeju, O.O.; Ekpo, O.E. Medicinal Properties and In Vitro Biological Activities of Selected Helichrysum Species from South Africa: A Review. Plants 2021, 10, 1566. [Google Scholar] [CrossRef] [PubMed]
- Etsassala, N.G.; Badmus, J.A.; Marnewick, J.L.; Egieyeh, S.; Iwuoha, E.; Nchu, F.; Hussein, A.A. Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities, Molecular Docking, and Antioxidant Capacities of Plectranthus ecklonii Constituents. Antioxidants 2022, 11, 378. [Google Scholar] [CrossRef] [PubMed]
- Etsassala, N.G.; Badmus, J.A.; Marnewick, J.L.; Iwuoha, E.I.; Nchu, F.; Hussein, A.A. Alpha-glucosidase and alpha-amylase inhibitory activities, molecular docking, and antioxidant capacities of Salvia aurita constituents. Antioxidants 2020, 9, 1149. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Cerretani, L.; Bendini, A. Rapid assays to evaluate the antioxidant capacity of phenols in virgin olive oil. In Olives and Olive Oil in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2010; pp. 625–635. [Google Scholar] [CrossRef]
- Nicoletta, P. Screening of dietary carotenoids and carotenoid-rich fruit extracts for antioxidant activities applying 2,2’-azinobis (3-ethylbenzothiazoline-6-sulfonic acid) radical cation decolorization assay. Method Enzymol. 1999, 299, 379–389. [Google Scholar] [CrossRef]
- Zhong, Y.; Shahidi, F. Methods for the assessment of antioxidant activity in foods. In Handbook of Antioxidants for Food Preservation; Woodhead Publishing: Sawston, UK, 2015; pp. 287–333. [Google Scholar] [CrossRef]
- Schrödinger Release 2021-4: Maestro; Schrödinger, LLC: New York, NY, USA, 2021.
- Schrödinger Release 2021-4: LigPrep; Schrödinger, LLC: New York, NY, USA, 2021.
- Schrödinger Release 2021-4: Protein Preparation Wizard; Epik, Schrödinger, LLC: New York, NY, USA, 2021.
- Schrödinger Release 2021-4: Impact; Schrödinger, LLC: New York, NY, USA, 2021.
- Schrödinger Release 2021-4: Prime; Schrödinger, LLC: New York, NY, USA, 2021.
- Schrödinger Release 2021-4: Glide; Schrödinger, LLC: New York, NY, USA, 2021.
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Gericke, B.; Schecker, N.; Amiri, M.; Naim, H.Y. Structure-function analysis of human sucrase-isomaltase identifies key residues required for catalytic activity. J. Biol. Chem. 2017, 292, 11070–11078. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.K.; Kang, S.C. Tyrosinase and α-glucosidase inhibitory effects of an abietane type diterpenoid taxodone from Metasequoia glyptostroboides. Natl. Acad. Sci. Lett. 2015, 38, 399–402. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [Green Version]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant activity/capacity measurement. 2. Hydrogen atom transfer (HAT)-based, mixed-mode (electron transfer (ET)/HAT), and lipid peroxidation assays. J. Agric. Food. Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Fons, L.; Garzon, M.T.; Micol, V. Relationship between the antioxidant capacity and effect of rosemary (Rosmarinus officinalis L.) polyphenols on membrane phospholipid order. J. Agric. Food. Chem. 2010, 58, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Karade, S.S.; Hill, M.L.; Kiappes, J.L.; Manne, R.; Aakula, B.; Zitzmann, N.; Warfield, K.L.; Treston, A.M.; Mariuzza, R.A. N-Substituted Valiolamine Derivatives as Potent Inhibitors of Endoplasmic Reticulum α-Glucosidases I and II with Antiviral Activity. J. Med. Chem. 2021, 64, 18010–18024. [Google Scholar] [CrossRef]
- Hammami, S.; Jannet, H.B.; Bergaoui, A.; Ciavatta, L.; Cimino, G.; Mighri, Z. Isolation and structure elucidation of a flavanone, a flavanone glycoside and vomifoliol from Echiochilon fruticosum growing in Tunisia. Molecules 2004, 9, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Carrera, B.; Clares, B.; Noé, V.; Mallandrich, M.; Calpena, A.C.; García, M.L.; Garduño-Ramírez, M.L. Cytotoxic evaluation of (2S)-5,7-dihydroxy-6-prenylflavanone derivatives loaded PLGA nanoparticles against MiaPaCa-2 cells. Molecules 2017, 22, 1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drewes, S.E.; van Vuuren, S.F. Antimicrobial acylphloroglucinols and dibenzyloxy flavonoids from flowers of Helichrysum gymnocomum. Phytochemistry 2008, 69, 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Marliyana, S.D.; Mujahidin, D.; Syah, Y.M. Pinostrobin Derivatives from Prenylation Reaction and their Antibacterial Activity against Clinical Bacteria. IOP Conf. Ser. Mater. Sci. Eng. 2018, 349, 012057. [Google Scholar] [CrossRef]
- Woo, S.U.; Jang, H.R.; Chin, Y.W.; Yim, H. 7-O-Methylwogonin from Scutellaria baicalensis disturbs mitotic progression by inhibiting Plk1 activity in Hep3B cells. Planta Med. 2019, 85, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Chae, H.S.; Xu, R.; Won, J.Y.; Chin, Y.W.; Yim, H. Molecular targets of genistein and its related flavonoids to exert anticancer effects. Int. J. Mol. Sci. 2019, 20, 2420. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Maheshwari, D.K.; Khandelwal, G. Antibacterial activity of Glycyrrhiza glabra roots against certain gram-positive and gram-negative bacterial strains. J. Appl. Nat. Sci. 2013, 5, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Fan, L.; Peng, C.; Zhang, Q.; Wang, L.; Li, L.; Wang, J.; Zhang, D.; Peng, W.; Wu, C. Traditional uses, botany, phytochemistry, pharmacology, pharmacokinetics and toxicology of Xanthium strumarium L.: A review. Molecules 2019, 24, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faizi, S.; Siddiqi, H.; Bano, S.; Naz, A.; Lubna; Mazhar, K.; Nasim, S.; Riaz, T.; Kamal, S.; Ahmad, A.; et al. Antibacterial and antifungal activities of different parts of Tagetes patula: Preparation of patuletin derivatives. Pharm. Biol. 2008, 46, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Bremner, P.D.; Meyer, J.J. Pinocembrin chalcone: An antibacterial compound from Helichrysum trilineatum. Planta Med. 1998, 64, 777. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.M.; Pereira, S.L.; Paiva, R.A.; Cavalcante, C.V.; Sudo, S.Z.; Tinoco, L.W.; Moreira, D.L.; Guimaraes, E.F.; Sudo, R.T.; Kaplan, M.A.; et al. Hypoglycemic effect of the methanol flower extract of piper claussenianum and the major constituent 2′,6′-dihydroxy-4′-methoxychalcone in streptozotocin diabetic rats. Indian J. Pharm. Sci. 2015, 77, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.F.; Karsani, S.A.; Yong, W.K.; Abd Malek, S.N. Induction of apoptosis and cell cycle blockade by helichrysetin in a549 human lung adenocarcinoma cells. Evid. Based Complement. Altern. Med. 2013, 2013, 857257. [Google Scholar] [CrossRef]
- Sutthanut, K.; Sripanidkulchai, B.; Yenjai, C.; Jay, M. Simultaneous identification and quantitation of 11 flavonoid constituents in Kaempferia parviflora by gas chromatography. J. Chromatogr. A 2007, 1143, 227–233. [Google Scholar] [CrossRef]
- Tomás-Lorente, F.; Iniesta-Sanmartín, E.; Tomás-Barberán, F.A.; Trowitzsch-Kienast, W.; Wray, V. Antifungal phloroglucinol derivatives and lipophilic flavonoids from Helichrysum decumbens. Phytochemistry 1989, 28, 1613–1615. [Google Scholar] [CrossRef]
- Wang, J.; Gao, H.; Zhao, J.; Wang, Q.; Zhou, L.; Han, J.; Yu, Z.; Yang, F. Preparative separation of phenolic compounds from Halimodendron halodendron by high-speed counter-current chromatography. Molecules 2010, 15, 5998–6007. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.E.; Han, M.J.; Kim, D.H. 3-Methylethergalangin isolated from Alpinia officinarum inhibits pancreatic lipase. Biol. Pharm. Bull. 2003, 26, 854–857. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Fang, C.; Zhu, R.; Peng, Q.; Li, D.; Wang, M. Inhibitory effect of phloretin on α-glucosidase: Kinetics, interaction mechanism and molecular docking. Int. J. Biol. Macromol. 2017, 95, 520–527. [Google Scholar] [CrossRef]
- Liu, J.; Lee, T.; DeFronzo, R.A. Why do SGLT2 inhibitors inhibit only 30–50% of renal glucose reabsorption in humans? Diabetes 2012, 61, 2199–2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Lee, T. SGLT2 inhibitors for type 2 diabetes. In Annual Reports in Medicinal Chemistry; Academic Press: Cambridge, MA, USA, 2011; Volume 46, pp. 103–115. [Google Scholar] [CrossRef]
- Shen, X.; Zhou, N.; Mi, L.; Hu, Z.; Wang, L.; Liu, X.; Zhang, S. Phloretin exerts hypoglycemic effect in streptozotocin-induced diabetic rats and improves insulin resistance in vitro. Drug Des. Dev. Ther. 2017, 11, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Du, Y.J.; Song, H.C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Den Hartogh, D.J.; Tsiani, E. Antidiabetic properties of naringenin: A citrus fruit polyphenol. Biomolecules 2019, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Shindo, K.; Kato, M.; Kinoshita, A.; Kobayashi, A.; Koike, Y. Analysis of antioxidant activities contained in the Boesenbergia pandurata Schult. rhizome. Biosci. Biotechnol. Biochem. 2006, 70, 2281–2284. [Google Scholar] [CrossRef]
- Francois, T.; Lambert, S.M.; Michel, J.D.P.; Gaby, N.M.E.; Fabrice, F.B.; Zaché, N.; Henri, A.Z.P.; Chantal, M. Composition, radical scavenging and antifungal activities of essential oils from 3 Helichrysum species growing in Cameroon against Penicillium oxalicum a yam rot fungi. Afr. J. Agric. Res. 2010, 5, 121–127. [Google Scholar] [CrossRef]
- Souza de Oliveira, F.F.; Coleho, P.N.; Gomes, G.J.; de Amorim, M.L.; Santos, P.S.; Mendonça, C.V.; Gonçalves, R.R.; Lins, M.G.; Pio, W.T.; Fuzer, C.F. Antioxidant Activity and Phytochemical Screening of Extracts of Erythroxylum suberosum A. St.-Hil (Erythroxylaceae). Res. J. Phytochem. 2015, 9, 68–78. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.L.; Kang, X.; He, X.; Dong, M.; Zhang, Q.; Liu, R.H. Cellular antioxidant activity of common fruits. J. Agric. Food Chem. 2008, 56, 8418–8426. [Google Scholar] [CrossRef]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Albayrak, S.; Aksoy, A.; Sagdic, O.; Hamzaoglu, E. Compositions, antioxidant and antimicrobial activities of Helichrysum (Asteraceae) species collected from Turkey. Food Chem. 2010, 119, 114–122. [Google Scholar] [CrossRef]
- Nyambe-Silavwe, H.; Villa-Rodriguez, J.A.; Ifie, I.; Holmes, M.; Aydin, E.; Jensen, J.M.; Williamson, G. Inhibition of human α-amylase by dietary polyphenols. J. Funct. Foods 2015, 19, 723–732. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | α-Glucosidase Inhibition IC50 (μM) | α-Amylase Inhibition IC50 (μM) |
|---|---|---|
| 1 | NA | NA |
| 2 | 204.23 ± 1.1 | NA |
| 3 | 44.44 ± 0.2 | NA |
| 4 | NA | NA |
| 5 | 12.94 ± 0.2 | NA |
| 6 | 206.45 ± 0.7 | NA |
| 7 | 18.16 ± 1.2 | NA |
| 8 | 103.59 ± 2.4 | NA |
| 9 | 86.20 ± 11.6 | NA |
| 10 | 9.24 ± 0.4 | 230.66 ± 15.8 |
| 11 | 16.00 ± 2.4 | NA |
| H. cymosum (TE) * | 35.8 ± 0.3 | NA |
| H. pandurifolium (TE) * | 69.28 ± 14.3 | NA |
| Acarbose | 945.46 ± 1.0 | 15.80 ± 0.6 |
| Sample | FRAP (µmol AAE/mmol) | TEAC (µmol TE/mmol) |
|---|---|---|
| 1 | 185.72 ± 1.1 | NA |
| 2 | 53.35 ± 0.7 | NA |
| 3 | 8.44 ± 0.24 | NA |
| 4 | 11.75 ± 0.9 | NA |
| 5 | 334.37 ± 1.7 | NA |
| 6 | 25.33 ± 0.6 | NA |
| 7 | 279.93 ± 0.8 | NA |
| 8 | NA | 73.62 ± 2.9 |
| 9 | 93.93 ± 7.1 | 275.39 ± 8.6 |
| 10 | 374.34 ± 69.7 | 301.37 ± 1.5 |
| 11 | 5.00 ± 2.8 | 138.33 ± 3.7 |
| H. cymosum (TE) * | NA | NA |
| H. pandurifolium (TE) * | 248.2 ± 0.1 | 722.4 ± 16.6 |
| Acarbose | 4858.29 ± 4.9 | 2677.00 ± 19.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jadalla, B.M.I.S.; Moser, J.J.; Sharma, R.; Etsassala, N.G.E.R.; Egieyeh, S.A.; Badmus, J.A.; Marnewick, J.L.; Beukes, D.; Cupido, C.N.; Hussein, A.A. In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents. Separations 2022, 9, 190. https://doi.org/10.3390/separations9080190
Jadalla BMIS, Moser JJ, Sharma R, Etsassala NGER, Egieyeh SA, Badmus JA, Marnewick JL, Beukes D, Cupido CN, Hussein AA. In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents. Separations. 2022; 9(8):190. https://doi.org/10.3390/separations9080190
Chicago/Turabian StyleJadalla, Baraa M. I. S., Justin J. Moser, Rajan Sharma, Ninon G. E. R. Etsassala, Samuel Ayodele Egieyeh, Jelili A. Badmus, Jeanine L. Marnewick, Denzil Beukes, Christopher N. Cupido, and Ahmed A. Hussein. 2022. "In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents" Separations 9, no. 8: 190. https://doi.org/10.3390/separations9080190
APA StyleJadalla, B. M. I. S., Moser, J. J., Sharma, R., Etsassala, N. G. E. R., Egieyeh, S. A., Badmus, J. A., Marnewick, J. L., Beukes, D., Cupido, C. N., & Hussein, A. A. (2022). In Vitro Alpha-Glucosidase and Alpha-Amylase Inhibitory Activities and Antioxidant Capacity of Helichrysum cymosum and Helichrysum pandurifolium Schrank Constituents. Separations, 9(8), 190. https://doi.org/10.3390/separations9080190