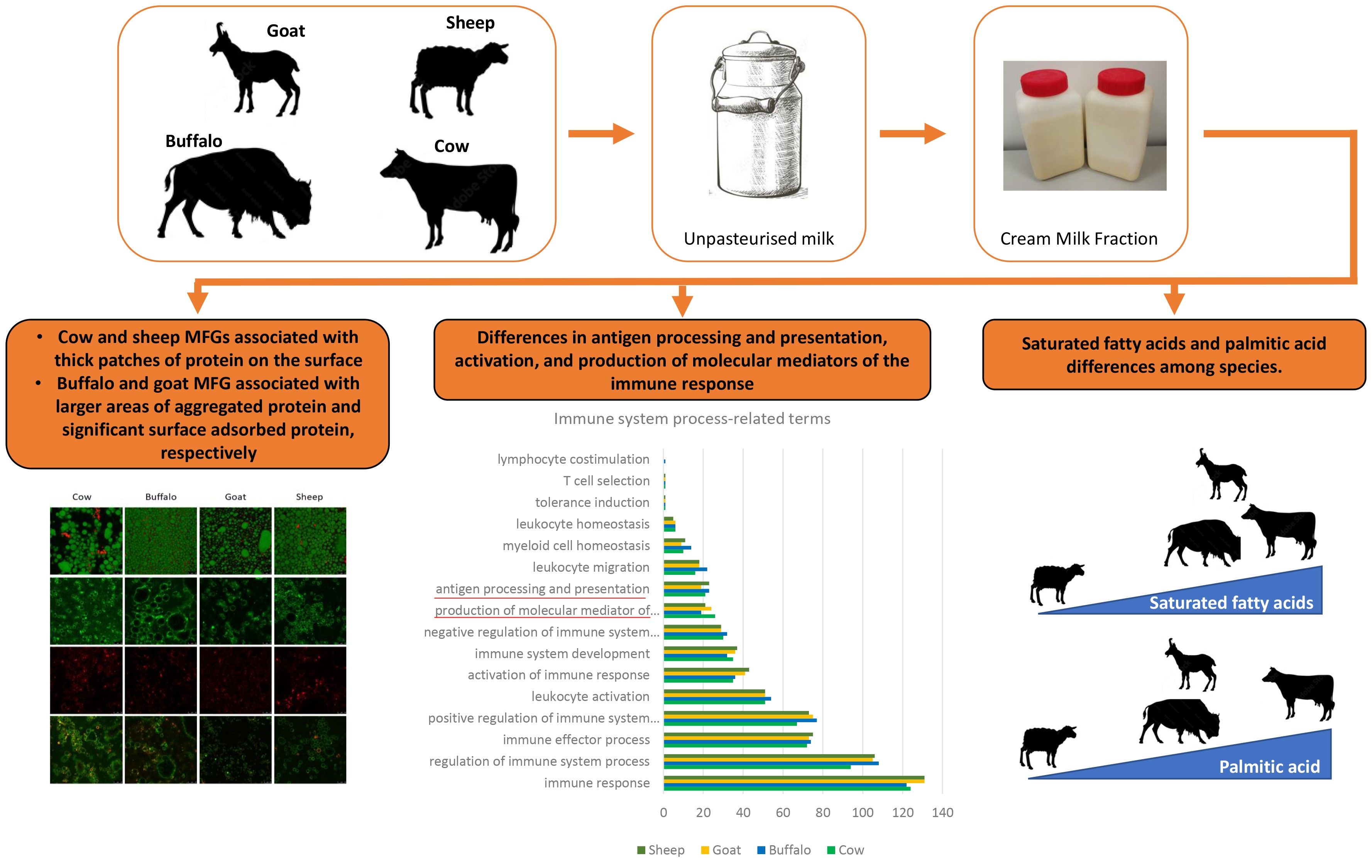
3.3. Functional Analysis of Cream Samples by Species
Functional analysis of all protein groups detected across the different cream samples was conducted using BLAST2GO analysis against a local BLAST database generated from the Bovidae entries in Uniprot (downloaded 24 May 2020) and gene ontology (GO) term mapping and annotation. GO terms are divided into 3 categories: biological process (BP), molecular function (MF) and cellular component (CC). For all species, “biological process” was the category with annotations for the highest number of protein groups. A summary of BP associated terms along with the number of protein groups identified from the cream samples of the associated species is shown in
Figure 3 (
n = 1) and includes: cellular process, biological regulation, metabolic process, regulation of biological process, response to stimulus, localization, multicellular organismal process, positive regulation of biological process, signalling, negative regulation of the biological process, developmental process, immune system process, interspecies interaction between organisms. An additional representation of this data is included alongside (right) showing the protein groups normalised to the total number of functionally annotated protein groups from that species. In addition to biological processes, GO molecular function (
n = 1;
Figure S1) and cellular component (
n = 1;
Figure S2) are shown for the four species comparisons, with absolute numbers of detected proteins (left) and the data normalised to the number of proteins with associated GO category terms (right) for each figure. There were higher numbers of detected proteins associated with developmental process, multicellular organismal process, response to stimulus and regulation of the biological process in the goat and sheep samples compared to buffalo and cow samples. Following normalisation, most terms showed similar levels across each of the species analysed. However, these data were limited to high-level broad BP terms; thus, to distinguish more subtle differences, further investigation of more specific BP terms and individual protein groups was undertaken.
Protein components of cream like those within the milk far globule membrane (MFGM) have been associated with several health benefits, including and predominantly immune function [
26]. Hence, we have further focused our analysis on immune system-related terms. A summary of immune system-related terms is shown in
Figure 4 (
n = 1) and includes regulation of immune system process, immune effector process, activation of immune response, immune system development, antigen processing and presentation, leukocyte migration, and myeloid cell homeostasis. A certain level of variability was observed in the GO terms among the species. Among those, we focused on antigen processing and presentation, activation of immune response and production of molecular mediator of immune response.
The antigen process and presentation was associated with 21 protein groups for cow, 23 for buffalo, 19 for goat and 23 for sheep cream (
n = 1;
Figure 4). When the proteins associated with this GO term were analysed, we observed several members of the RAS family (
n = 15;
Table 2). Of these proteins, eight were present in all species; two were present in cow, goat and sheep but not in buffalo; two were present in buffalo and goat and three proteins were specific for either buffalo, goat or sheep. Members of the RAS family have also been previously detected in human skim milk [
27] as well as in the MFGM fraction of buffalo milk [
28]. Several Ig-like domain-containing proteins were also detected across all species (
Table 2). The proteasome activator complex subunit 1 was only detected in buffalo and sheep while the protein prosaposin was only detected in cow and sheep (
Table 2). Prosaposin is the precursor protein for four lysosomal activator proteins, saposins A-D, which act as sphingolipid activator proteins that facilitate the hydrolysis of sphingolipids via lysosomal hydrolases [
29]. In addition to this function, the full-length protein can also be secreted into several secretory fluids, including milk [
30], where it acts as neurotrophic factor, promoting cell survival, neurite outgrowth and differentiation in a cholinergic cell line [
31,
32]. Prosaposin has been shown to be present in human milk as well as in milk of other species including cow and goat [
30,
33]. However, in our study, this protein was only detected in cow and sheep cream but not in buffalo or goat cream. This could either indicate that it is present at concentrations below the limit of detection or that levels could differ in cream versus milk.
Within the GO term activation of the immune response, 35 proteins were detected for cow, 36 for buffalo, 41 for goat, and 43 for sheep (
n = 1;
Figure 4). From this group, several members of the complement family were detected across all species, with few uniquely detected in some species (
Table 3). The complement system includes plasma proteins that coat extracellular pathogens, facilitating their removal by phagocytes or direct killing. MFGM-related proteins associated with activation of the immune response, such as butyrophilin subfamily 1 member A1 and tyrosine-protein kinase, were also identified across all species (
Table 3). An interesting protein that was observed in all species except cow is pentraxin (
Table 3). This protein family includes the short pentraxins serum amyloid P component, C-reactive protein, and the prototypic long pentraxin PTX3. Pentraxins are part of the pattern recognition receptors (PRRs), which are specialized in recognition of highly conserved motifs expressed by microbes. These receptors are classified based on their localization and include (1) endocytic PRRs, (2) signaling PRRs and (3) soluble PRRs, which include pentraxins, collectins and ficolins. These proteins can bind selected microbes and facilitate their disposal by phagocytes [
34]. PTX3 has been shown to be present in human breast milk, potentially contributing to the protection of infants against infections [
35]. Interestingly, Mudaliar and colleagues have shown upregulation of PTX3 in an experimental model of cow mastitis [
36]. This protein is indeed produced at sites of infection and inflammation by both somatic and immune cells, and its glycosylation has been implicated in modulating protein functions, including the modulation of the complement system through the interaction with the complement component C1q (reviewed by [
37]). Interestingly, our analysis has identified complement C1q A chain and B chain in goat and sheep, and in sheep, respectively, while neither were detected in cow and buffalo (
Table 3). This could again either indicate that these proteins are present at concentrations below the limit of detection.
Another protein that was detected in all species, except cow, is the major prion protein (PrP) (
Table 3). This protein has received considerable attention due to its role in the pathogenesis of prion disease or spongiform encephalopathies (TSEs), affecting both humans and animals. The normal PrP (PrP
C) is apparently benign. However, it is capable of post-translational misfolding into an abnormal and infectious isoform (PrP
Sc) [
38]. The normal PrP
c has been detected, with differences in expression, in the mammary gland of domestic ruminants such as cow, sheep and goat [
39,
40,
41], as well as in humans, cow, sheep and goat milk [
42,
43]. In contrast to these findings, Didier and colleagues [
40], did not detect PrP
C in any of the cow milk fractions analysed in their study, including cream, which is consistent with what we have observed in our study. However, this is probably due to the methodology used in Franscini’s study, which leads to a higher concentration of the prion content. Interestingly, PrP
C was easily detected in sheep and goat milk fractions, with the highest levels observed in the cream fraction [
40]. This evidence suggests that PrP may be present in our cow cream sample but below the limits of detection, while the higher levels present in goat and sheep cream allowed for its detection. No evidence has been shown to date for the presence of PrP in buffalo milk.
Alpha-2-macrogobulin (A2M) is a plasma protein involved in the inhibition of a wide range of serum proteases. While we have only detected this protein in goat and sheep cream (
Table 3), in a previous study, A2M was detected in cow milk, with the highest concentration in the first milking [
44]. This could be related to the capacity of A2M to inhibit the protease, hence preventing the degradation of biologically active proteins (i.e., immunoglobulins) in the intestine of the newborn. Furthermore, A2M plays a key role as a humoral defence barrier against pathogens, binding host or foreign peptides and particles [
45]. Interestingly, higher concentrations of A2M have been detected in mastitic cow’s milk compared to normal milk and this was related to the degree of mastitis [
44].
Toll-like receptor 4 (TLR4) was detected only in buffalo under the GO terms activation of immune response and production of molecular mediator of the immune system (
Table 3 and
Table 4). As with pentraxins, Toll-like receptors are PRRs. TLR4 recognises bacterial lipopolysaccharide (LPS), which leads to the activation of the intracellular signalling pathway, NF-κB and the subsequent production of inflammatory cytokine, activating the innate immune system [
46]. The presence of TLR4 has been previously investigated in breast milk (MFGM and skimmed fraction) by Cattaneo and colleagues [
47]. In this study, TLR4 was not detected in the samples analysed; however, the group concluded that this result cannot assure the total absence of this receptor in milk but could also be due to the limit of detection of the instrument or the methodology used. Interestingly, Cao and colleagues [
48], detected this receptor in both colostrum and mature milk MFGM with higher levels found in the latter. TLR4 was also detected in the sheep milk whey protein fraction [
49], while to our knowledge, there is no evidence of the presence of TLR4 in milk or milk fractions in cow, goat, and buffalo.
Under the GO term production of molecular mediator of the immune system, 26 proteins were detected for cow, 19 for buffalo, 24 for goat and 21 for sheep (
n = 1;
Figure 4). MFGM related proteins such as platelet glycoprotein 4/CD36, Toll-like receptor 2, apolipoprotein A1 have been identified across all species as well as several Ig-like domain-containing proteins (
Table 4).
An interesting protein that was detected in cow, goat and sheep cream but not in buffalo is the transforming growth factor beta-2 proprotein (
Table 4). The protein intensities, however, were low in these three species; thus, it could be the case that even lower levels were present in buffalo that went undetected as opposed to this protein being absent. The transforming growth factor beta-2 proprotein is the precursor of the latency-associated peptide (LAP) and transforming growth factor beta-2 (TGF-β2) chains, which constitute the regulatory and active subunit of TGF-β2, respectively. TGF-β, is the most abundant cytokine in breast milk, and includes TGF-β1 and TGF-β2, with the latter being predominant [
50,
51]. Breast milk TGF-β has gained increasing interest as it is involved in maintaining intestinal homeostasis, regulating inflammatory responses and promoting the development of neonatal oral tolerance [
52,
53]. Breast milk TGFβ2 has also been shown to be associated with the neonatal gut microbial composition and increased richness, evenness, and diversity [
54]. The relationship between TGFβ and allergies prevention is controversial. An initial systematic review showed that high concentrations of TGF-β1 or TGF-β2 in human milk, were positively associated with a reduction in immunological outcomes of allergies in seven of 12 studies included [
55]. However, the result of a recent systemic review was in contrast with this conclusion, not finding strong evidence of associations between any isoform of human milk TGF-β and allergic outcomes [
56]. Both TGF-β isoforms have also been identified in cow milk with TGF-β2 being again the most abundant [
57]. In cow milk both TGF-β2 and TGF-β1 levels has been shown to increase during mastitis induced by
E. coli [
58]. To our knowledge this is the first time that TGF-β2 has been shown in goat and sheep cream.
Another cytokine that was detected in cow, goat and sheep but not in buffalo is the macrophage migration inhibitory factor (MIF) (
Table 4). This cytokine is released in response to proinflammatory stimuli and inhibits the migration of macrophages, enhancing their phagocytic activity. The presence of MIF in human milk was identified for the first time by Magi and colleagues [
59], in the milk aqueous phase and inside milk fat globules. This was further confirmed by Vigh and colleagues [
60], who detected a high concentration of MIF in breast milk, especially during the first month of lactation. No studies to date have shown the presence of MIF in the milk of any of the species subject of this study.
3.4. Lipid Analysis of Cream Samples from Four Species
Milk fat contains several thousand lipid species and is the most complex material in nature in terms of lipid composition. The fatty acid (FA) composition of milk fat triglycerides (which account for approximately 98% of total milk lipids) are affected by several factors, including species, breed, diet, and seasonality [
61]. The milk fat of domesticated animals is composed primarily of two major fractions: long-chain (50–70%) and short-chain FAs (30–50%).
Long-chain FAs (C18-24) are typically derived from the diet, whereas short-chain FAs (C4 to C14 and some C16), are synthesized de novo by the mammary gland. Saturated FAs in ruminant milk accounts for approximately 60 to 70% of the total.
In the present study, total fat (
n = 1;
Table 5) and fatty acid content of all cream samples were determined. The levels of saturated fatty acids (SFA) were lowest in sheep cream and broadly similar in the cream of the other species examined (
n = 1;
Table 6). MacGibbon and Taylor [
62] and Markiewicz-Kęszycka et al. [
63] also reported lower total SFA in sheep milk compared to cow and goat. In terms of short-chain FAs, buffalo cream contained the highest levels of butyric acid (C4:0), whereas the levels of caprylic (C8:0) and capric (C10:0) acids were much higher in sheep and goat milk fat than that of cow and buffalo. Caproic, caprylic and capric acids are so termed because of their high proportions in goat milk. Variations in de novo synthesis of short-chain FAs are controlled by several genes expressed in the mammary gland [
64], and may account for the differences observed between species. Palmitic acid (C16:0) was highest in cow cream (32.2%) and was, with the exception of sheep milk fat (19.7%
w/w), the most abundant of all FAs. Castro-Gomez et al. [
65] reported greater similarities in C16:0 between cow, sheep and goat milks with values of 32, 29 and 28%, respectively. Palmitic acid, along with a number of other high-melting-point FAs, is the main contributor to hardness in milk fat products as it remains solid at room temperature.
Oleic acid (C18:1), the most abundant monounsaturated FA (MUFA) in mammalian milk fat, was highest in sheep milk and was very similar in the other three species. In contrast with C16:0, it is a low-melting FA and thus, contributes to the liquid phase in semi-crystalline dairy products, thereby contributing to softness and spreadability. Other MUFAs in ruminant milk include myristoleic acid (C14:1), which was much lower in sheep and goat cream compared to that of cow or bubalus bubalis; palmitoleic (C16:1), lowest in goat milk, and vaccenic acid (C18:1 trans), a naturally occurring trans FA with cholesterol-lowering functionality.
Generally speaking, milk fat of non-ruminants has a higher level of polyunsaturated FAs (PUFA) than that of ruminants, due predominantly to direct absorption from the diet [
62]. Sheep milk fat had higher levels of PUFA than the other species examined, a finding supported by previous reports [
62,
66,
67]. This difference is attributable, in the main, to the high levels of linoleic acids (C18:2), a diverse group of FA isomers, which includes α-linoleic acid (C18:2
n-6, cis), one of two essential FAs, and a number of conjugated linoleic acid isomers (CLA), a group of FAs associated with a range of health benefits. Serra et al. [
67] reported that feeding with linseed oil significantly increased levels of linolenic (C18:3) and rumenic acid (C18:2 CLA) in sheep milk, with both of these also present in higher quantities at the
sn-2 (middle) position of the triglyceride molecule. Markiewicz-Kęszycka et al. [
63] reported that sheep and goat milks are usually richer in CLA than cow milk and that the concentration of CLA in the milk fat of sheep milk is greater than that in goat milk, with this effect due to differences in the mRNA of their mammary adipocytes. The level of C18:3, the other fatty acid essential to human nutritional needs, was significantly lower in goat cream, compared to the other species. Arachidonic acid (C20:4,
n-6) was approximately twice as high in sheep cream as cow, goat and buffalo milk fat.
In general, the FA profiles of cow and buffalo milk fat were, with the exception of butyric, palmitic and linoleic acids, similar in composition. Pegolo et al. [
68] reported that
buffalo and cow species have comparable average milk FA although others have reported greater variations [
69,
70]. The lipid composition of milk responds to changes in diet in a more pronounced way compared to other macro-constituents such as proteins, which are determined largely by genetics. FA profile can be manipulated through direct addition (supplementation) of FAs in the diet, alteration of rumen conditions, or via biotransformation, in the rumen, of dietary FAs to other (long-chain) FA species. Pasture feeding increases the concentration of certain milk FAs, mainly C18:0 C18:1 C18:3 and CLA, and decreases saturated FAs from C10:0 to C16:0 [
46,
54]. Mixed-ration (concentrated) feeds have been reported to yield higher levels of C16:0 and lower levels of nutritionally beneficial FAs [
5]. The health implications of milk fat consumption have, for many years, generated much controversy and confusion, resulting in commonly held negative perceptions. More recently, meta-analytical studies have redressed this imbalance and have contributed to a gradual shift in scientific opinion that considers the contribution of SFAs to human health to be less detrimental than previously thought. This topic has been the subject of recent reviews by Mohan et al. [
61] and Lordan and Zabetakis [
71].
3.4.1. Fatty Acid Positional Distribution
During digestion, fats undergo enzymatic hydrolysis by pancreatic lipases, which cleave FAs preferentially from the sn-1 and sn-3 positions, i.e., the outer positions of the triglyceride molecule, leaving the central sn-2 FA attached to the glycerol molecule. These sn-2 monoglycerides are freely absorbed regardless of the remaining FA type. The rate of absorption of free FAs (FFAs) emanating from the sn-1 and sn-3 positions depends on FA chain length with longer chain FAs (C12:0 to C18:0) less readily absorbed than shorter chain FAs (C6:0 to C10:0). As such positional esterification of FAs is important in terms of nutrition and energy provision to the neonate. Human milk has an unusually high proportion of C16:0 at the sn-2 position (60% or more), which facilitates absorption and digestion, and has led to recent interest in the provision of infant formulae (IF) with this triglyceride structure.
Vegetable oils, traditionally used in IF, have much lower levels of palmitic acid at
sn-2. Comparison of the
sn-2 content of the milk fats examined in this study (
n = 1;
Table 7) demonstrate that cow cream had the highest level of palmitic acid (41.5%) with sheep the lowest at 29.5%. Others have reported similar values [
72,
73] for cow milk fat with rather less data available for other species [
67,
74,
75]. Blasi et al. [
76] reported %
sn-2 values for C16:0 of 44.1, 35.7, 27.2 and 40.9 for cow, goat, sheep and buffalo milk, respectively. Although comparatively lower than human milk with respect to
sn-2 C16:0 levels cow milk still represents a valuable source and is a promising substrate for lipase-induced structural modification of milk fat triglycerides for inclusion into IF products. In the case of
sn-1 and
sn-3 FA positioning, the results of the present study are similar to those published by Blasi et al. [
76] and summarised by MacGibbon and Taylor [
62] and Mohan et al. [
61]. Studies suggest that the regiospecific binding of FA is likely to be genetically based [
77]. The influence of the positional distribution of FAs on both nutrition and functional properties (melting and crystallisation) of lipid triglycerides remains poorly understood.
3.4.2. Phospholipids
The polar lipids of milk are the main constituents of the MFGM; the tri-layer film that stabilises milk fat globules against coalescence. The composition of the MFGM ranges widely, with polar lipids accounting for 30–75% and proteins making up 25–75% [
78]. Polar lipids make up only 0.4–1% of total milk lipids. Phospholipids (PL) are a sub-class of polar lipids comprised of glycerophospholipids and sphingolipids, which have well-established nutraceutical properties [
79]. The main polar lipids of milk are phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylinositol (PI), phosphatidylserine (PS) and sphingomyelin (SM). In the present study, goat milk fat had the highest total % weight of PL, with buffalo the lowest (
n = 1;
Table 7). The average fat globule size is generally lower in small ruminant species, compared to cow (goat < sheep < cow), and has a greater proportion of PL (as seen in the content of goat milk fat), a requirement for the stabilisation of relatively higher fat globule surface areas. In contrast, buffalo cream has a higher fat content than the other species examined but also has larger fat globules, and accordingly has a lower proportion of polar lipids relative to total fat [
69]. PE accounted for the highest proportion of PLs, with the exception of cow cream in which PC was marginally higher (31.4 vs. 28.6% of total PL). PI and PS values were the lowest overall. It should be noted that studies indicate that variability in the proportions of PLs in the MFGM can stem from the methods used for PL extraction rather than inter-species variation [
80].









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}