
The Beneficial Effect of Coarse Cereals on Chronic Diseases through Regulating Gut Microbiota


Abstract
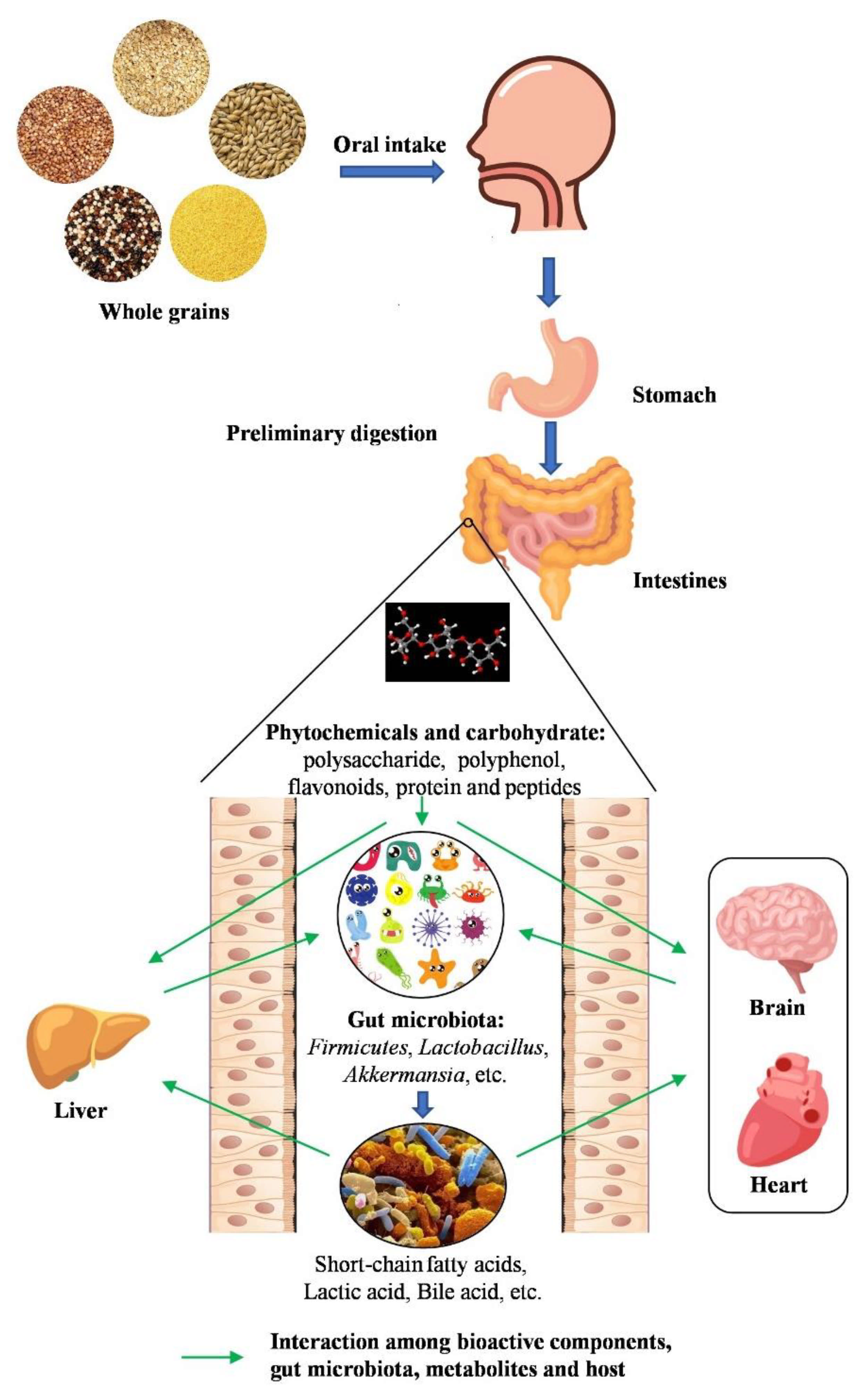
:1. Introduction
2. Effects on Gastrointestinal Diseases
2.1. Inflammatory Bowel Disease
2.2. Colorectal Cancer

{kind=link}
| Cereal | Component | Pathological Type | Study Characteristics | Pathological Parameters | Changes in Gut Microbiota | Reference |
|---|---|---|---|---|---|---|
| barley | leaf | colitis | female C57Bl/6J mice | IL-4 ↑, IL-10 ↑, TNF-α ↓, stool frequency ↑, gut transit time ↓, inosine ↑, guanosine ↑, glucose ↓, lactic acid ↑, body Weight ↑ | Proteobacteria ↓, Enterobacteriaceae ↑, Firmicutes ↑, Bacteroidetes ↓, Lactobacillus ↑ | [29] |
| insoluble fiber | colitis | female C57BL/6J mice | IL-6 ↓, TNF-α ↓, IL-1β ↓, IL-4 ↓, IL-10 ↓, SCFAs ↑, secondary bile acids ↑, body Weight ↑ | Akkermansia ↓, Parasutterella ↑, Erysipelatoclostridium ↑, Alistipes ↑, Verrucomicrobia ↓ | [28] | |
| buckwheat | whole grain | gastritis | SPF male C57BL/6 mice | IL-6 ↓, IL-1β ↓, IL-18 ↓, TH17 ↓, TGF-β ↓, IL-17A ↓, ILA ↑ | Unclassified_Lachnospiraceae ↓, Un-classified_Clostridiales ↓, Unclassified_Rikenellaceae ↓, Oscil-lospira ↓, Unclassified_S24-7 ↑, Lactobacillus ↑ | [33] |
| millet | polyphenol | colorectal cancer | male C57BL/6J mice | COX-2 ↓and EMR1 ↓, PCNA cells ↓, caspase 3 ↑ | Firmicutes ↑, Bacteroidetes ↓, Prevotella ↓, Corprobacillus ↑, Parabacteroides ↑, AF12 ↑, Coprococcus ↑, Oscillospira ↑, Ruminococcus ↑, Prevotella ↓, Desulfovibrio ↓ | [38] |
| tryptophan and fiber | colorectal cancer | SPF male BALB/c mice | IL-6 ↓, IL-17 ↓, MPO ↓, MCP-1 ↓, serum C-P ↓, IFN-γ ↓, LSP ↓, COX-2 ↓, iNOS ↓, FOXP3 ↑, IL-22 ↑, ZO-1 ↑, IL-10 ↑, occludin ↑, Bcl-2 ↑, PCNA ↑, VEGF ↑, AHR ↑, SCFAs ↑ | Allobaculum ↑, Bifidobacterium ↑, Bacteroidales_S24-7 ↑, Alistipes ↓, | [36] | |
| whole grain | acute ulcerative colitis | male C57BL/6 mice | IL-6 ↓, claudin2 ↓, ZO-1 ↑, occludin ↑, claudin 2↓ | Muribaculaceae ↑ | [34] | |
| oat | bran | inflammatory | pig | IL-8 ↓, colonic IL-8 ↓, NF-κB ↓, TNF-α ↓ | Catenibacterium ↑, Peptococcus ↓, Prevotella ↑, Butyricicoccus ↑, Catenibacterium ↑, Coprococcus ↓, Desulfovibrio ↓ | [39] |
| β-glucan | enteritis | male SD rats | IL-12 ↓, IL-1α ↓, β, IL-6 ↓, IL-10 ↑, TNF-alpha ↑, lactic acid ↑, propionic acid ↑, hydroxybutyric acid ↓ | - | [25] | |
| fiber | necrotic enteritis | broiler chickens | succinic acid ↑, acetic acid ↓, propionic acid ↓, valeric acid ↓ | Perfringens ↑, Lactobacillus ↓, Salmonellae ↓, | [27] | |
| quinoa | whole grain | colonic colitis | male C57BL/6 mice | IL-6 ↓, IL-1β ↓, IFN-γ ↑ | Proteobacteria ↓, Escherichia/Shigella ↓, Peptoclostridium↓, Bacteroidetes ↑, Verrucomicrobia ↑ | [32] |
| sorghum | whole grain | colorectal cancer | humanfemale C57BL/6 mice | - | Bacteroides ↑, Fusobacterium ↑, Dorea, Porphyromonas ↑, Pseudomonas ↓, Prevotella ↓, Acinetobacter ↓, Catenibacterium ↓ | [37] |
3. Effects on Cardiovascular Diseases
3.1. Hyperlipidemia
3.2. Hypertension
3.3. Nonalcoholic Fatty Liver Disease
3.4. Atherosclerosis
| Cereal | Component | Pathological Type | Study Characteristics | Pathological Parameters | Changes in Gut Microbiota | Reference |
|---|---|---|---|---|---|---|
| buckwheat | fermented milk | dyslipidemia | males C57BL/6 rats | weight ↓, LPS ↑, TNF-α ↑, IL-6 ↑, SOD ↓, T-AOC ↓, CAT ↓, MAD ↑, acetic ↑, propionate ↑, total acid ↑ | Firmicutes ↓, Bacteroidetes ↑, norank-f-Bacteroidales-S24-7-group ↓, unclassified-f-Lachnospiraceae ↓, Blautia ↓, Enterobacter ↑, Akkermansia ↑ | [56] |
| whole-grain-based food | nonalcoholic fatty liver disease | male Kunming mice | SCFAs ↑, bile acid ↑, weight ↓, TC ↓, TG ↓, LDL-C ↓, LPS ↓, ALT ↓, AST ↓ | Blautia ↑, Lactobacillus ↑, Streptococcus ↑, unclassified_Rikenellacea ↑, Desulfovibrio ↓, Bacteroidales_S24-7 ↓, Allobaculum ↓ | [71] | |
| hypercholesterolemia | male gold hamsters | TC ↓, LDL-cholesterol ↓, CE ↓, TG ↓, BA ↑, acetate ↑, propionate ↑, butyrate ↑, SCFAs ↑ | Erysipelotrichaceae ↓, Ruminococcaceae ↓, Lactobac-illaceae ↓, Lachnospiraceae ↓, Eubacteriaceae ↑, Bacteroide-tes ↓, Ruminococcus-1 ↓, Ruminococcus-2 ↓, Ru-minococcuceae-UGG-014 ↓ | [60] | ||
| fermented common buckwheat | non-alcoholic fatty liver disease | SPF male Kunming mice | weight ↓, TG ↓, TC ↓, LDL-C ↓, AST ↓, ALT ↓, TBA ↓, NEFA ↓, HDL-C ↑ | Ruminiclostridium ↑, Lacobacillus, Alistipes ↑, Bacteroidales S24-7 group ↓, Clostridiales XIII ↓ | [76] | |
| fermented black tartary buckwheat | hyperlipidemia | male SD rats | tyrosine ↑, lysine ↑, total flavonoids ↑, total polyphenols ↑, quercetin ↑, kaempferol ↑, rutin ↓ | Lactobacillus ↑, Faecalibaculum ↑, Allobaculum ↑, Romboutsia ↓ | [84] | |
| d-fagomine | hypertension | male WKY rats | uric acid ↑ | Bacteroidetes ↑, enterobacteriales ↑, E. colipopulations ↑ | [66] | |
| protein | dyslipidemia | SPF Male C57BL/6 mice | TC ↓, TG ↓, total bile acids ↑, SCFAs ↑, LPS ↓, TNF-α ↓, IL-6 ↓ | Lactobacillus ↑, Bifidobacterium ↑, Enterococcus ↑, Escherichia coli ↓ | [54] | |
| millet | arabinoxylan | lipid derangements, endotoxemia | male mice (Swiss albino, LACA strain) | total cholesterol ↓, HDL-C ↓, LDL-C ↓, VLDL-C ↓, NEFA ↓, FASN ↓, ACC ↓, SREBP-1c ↓, HMGCOA-R ↓, HLST ↑, TNF-α ↓, IL-1β ↓, IL-6 ↓, CRP ↓ | Bacteroidales S24-7 ↑, Blautia sp. ↓, Lactobacillus sp. ↓, Bifidobacterium sp. ↓, Roseburia ↓, Bacteroidetes ↓ | [85] |
| millet shell polyphenols | atherosclerosis | male ApoE−/−mice | LSP ↓, TNF-α ↓, IL-1β ↓, occludin ↑, zona occludens-1 ↑, claudin1 ↑ | Verrucomicrobia ↓, Actinobacteria ↓, Oscillospira ↓, Ruminococcus ↓, Bacteroidetes ↑, Allobaculum ↑ | [81] | |
| vitexin | brain oxidative stress and inflammation | male C57BL/6 N mice | SOD ↑, CAT ↑, GPx ↑, MDA ↓, Keap1 ↓, TNF-α ↓, IL-1β ↓, IL-6 ↓, IL-10 ↓ | Firmicutes ↓, Verrucomicrobiota ↑, Lachnospiraceae ↑, Escherichia-shigella ↑ | [86] | |
| oat | fiber | hypertension | human | SCFAs ↑ | Spirillum ↑, Bifidobacterium ↑, Trichosporium ↑ | [65] |
| fiber | atherosclerosis | male LDLR−/−mice | GFAP ↓, IBα1 ↓, SCFAs ↑, zonula occludens-1 ↑, occludin ↑, LSP ↓ | Actinobacteria ↑, Peptostreptococcaceae ↑, Coriobacteriaceae ↑, Eisenbergiella ↑, Romboutsia ↑, Rikenellaceae ↓, Anaerotruncus ↓, Parabacteroides ↓ | [83] | |
| flavonoids | hyperlipidemia | male C57BL/6N mice | weight ↓, PPARα ↑, CPT-1 ↑, CYP7A1 ↑, FXR ↑, TGR5 ↑, NTCP ↑, BSTP ↑, SREBP-1c ↓, FAS ↓, ASBT ↓ | Akkermansia ↑, Lachnoclostridium ↓, Blautia ↓, Colidextribacter ↓, Desulfovibrio ↓ | [47] | |
| β-glucan | cardiovascular disease | apo-E−/−mice | total cholesterol ↓ | Verrucomicrobia ↓, Proteobacteria ↑, Peptostreptococcaceae ↓, Christenellaceae ↓, Desulfovibrionaceae ↑, Bacteroidaceae ↑, Helicobacter ↑, Akkermansia ↑, Ruminococcus spp. ↑ | [87] | |
| whole grain | cholesterol | human | TC ↓, LDL-C ↓ | Bifidobacteria ↑, lactobacilli ↑ | [62] | |
| whole grain | cholesterol and diabetes | male C57BL/6J mice | weight ↓, insulin ↓, plasma total cholesterol ↓ | Lachnospiraceae ↑ | [61] | |
| oat ethanol extracts | lipid metabolic disorder | male SD rats | the total SCFAs ↑, acetate ↑, propionate ↑, SREBP-1C ↓, FAS ↓, HMGCR ↓, TG ↑, TC ↑, LDL-C ↑, HDL-C ↑ | Bifidobacterium ↑, Akkermansia ↑, Rombutsia ↓, Fusicatenibacter ↓, Holdemanella ↓, Turicibacter ↓ | [48] | |
| quinoa | soluble polysaccharide | hyperlipidemia | male SD rats | TG ↓, LDL-C ↓, MDA ↓, ASTl ↓, TC ↓, ALT ↓, HDL-C ↑, SCAFs ↑ | the ratio of Firmicutes ↓, Bacteroides ↓, Proteobacteria ↓, Desulfovibrio ↓, Allobaculum ↓ | [50] |
| protein | antihypertensive | SHRs and WKY rats | Allobaculum ↑, Turicibacter ↑, Staphylococcus ↑ | [67] |
4. Effects on Obesity
5. Effects on Diabetes
6. Prebiotic Effect
7. Effect on Other Chronic Diseases
7.1. Metabolic Syndrome
7.2. Neurodegenerative Disorders
7.3. Malnutrition
8. Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Fu, J.; Zhang, Y.; Hu, Y.C.; Zhao, G.; Tang, Y.; Zou, L. Concise review: Coarse cereals exert multiple beneficial effects on human health. Food Chem. 2020, 325, 126761. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.B.; Wang, Y.Y.; Shabani, K.I.; Qin, X.L.; Liu, X. Comparison of structural features of reconstituted doughs affected by starches from different cereals and other botanical sources. J. Cereal Sci. 2020, 93, 102937. [Google Scholar] [CrossRef]
- Lie, L.; Brown, L.; Forrester, T.E.; Plange-Rhule, J.; Bovet, P.; Lambert, E.V.; Layden, B.T.; Luke, A.; Dugas, L.R. The Association of Dietary Fiber Intake with Cardiometabolic Risk in Four Countries across the Epidemiologic Transition. Nutrients 2018, 10, 628. [Google Scholar] [CrossRef] [Green Version]
- Idehen, E.; Tang, Y.; Sang, S.M. Bioactive phytochemicals in barley. J. Food Drug Anal. 2017, 25, 148–161. [Google Scholar] [CrossRef] [Green Version]
- Llaverias, G.; Escola-Gil, J.C.; Lerma, E.; Julve, J.; Pons, C.; Cabre, A.; Cofan, M.; Ros, E.; Sanchez-Quesada, J.L.; Blanco-Vaca, F. Phytosterols inhibit the tumor growth and lipoprotein oxidizability induced by a high-fat diet in mice with inherited breast cancer. J. Nutr. Biochem. 2013, 24, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Hehemann, J.H.; Correc, G.; Barbeyron, T.; Helbert, W.; Czjzek, M.; Michel, G. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 2010, 464, 908–912. [Google Scholar] [CrossRef] [PubMed]
- De Vos, W.M.; de Vos, E.A.J. Role of the intestinal microbiome in health and disease: From correlation to causation. Nutr. Rev. 2012, 70, S45–S56. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Devkota, S.; Wang, Y.W.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10(-/-) mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henao-Mejia, J.; Elinav, E.; Jin, C.C.; Hao, L.M.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Costabile, A.; Klinder, A.; Fava, F.; Napolitano, A.; Fogliano, V.; Leonard, C.; Gibson, G.R.; Tuohy, K.M. Whole-grain wheat breakfast cereal has a prebiotic effect on the human gut microbiota: A double-blind, placebo-controlled, crossover study. Br. J. Nutr. 2008, 99, 110–120. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Probert, H.M.; Van Loo, J.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Xing, X.; Zhen, H. Enzyme deactivation treatments did not decrease the beneficial role of oat food in intestinal microbiota and short-chain fatty acids: An in vivo study. J. Sci. Food Agric. 2013, 93, 504–508. [Google Scholar] [CrossRef]
- Berger, K.; Falck, P.; Linninge, C.; Nilsson, U.; Axling, U.; Grey, C.; Stalbrand, H.; Karlsson, E.N.; Nyman, M.; Holm, C.; et al. Cereal Byproducts Have Prebiotic Potential in Mice Fed a High-Fat Diet. J. Agric. Food Chem. 2014, 62, 8169–8178. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharm. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.B.; Pere-Trepat, E.; Montoliu, I.; Martin, F.P.J.; Collino, S.; Moco, S.; Godin, J.P.; Cleroux, M.; Guy, P.A.; Breton, I.; et al. A Whole-Grain-Rich Diet Reduces Urinary Excretion of Markers of Protein Catabolism and Gut Microbiota Metabolism in Healthy Men after One Week. J. Nutr. 2013, 143, 766–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.Y.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A.; et al. Gut microbiome composition is linked to whole grain-induced immunological improvements. ISME J. 2013, 7, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Tome, S.; Hernandez-Ledesma, B.; Chaparro, M.; Indiano-Romacho, P.; Bernardo, D.; Gisbert, J.P. Role of food proteins and bioactive peptides in inflammatory bowel disease. Trends Food Sci. Technol. 2019, 88, 194–206. [Google Scholar] [CrossRef]
- Gassull, M.A. Review article: The intestinal lumen as a therapeutic target in inflammatory bowel disease. Aliment. Pharm. Ther. 2006, 24, 90–95. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Larussa, T.; Imeneo, M.; Luzza, F. Potential role of nutraceutical compounds in inflammatory bowel disease. World J. Gastroenterol. 2017, 23, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Albenberg, L.; Compher, C.; Baldassano, R.; Piccoli, D.; Lewis, J.D.; Wu, G.D. Diet in the Pathogenesis and Treatment of Inflammatory Bowel Diseases. Gastroenterology 2015, 148, 1087–1106. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.Y.; Zhao, J.J.; AL-Ansi, W.; Wang, J.; Xue, L.M.; Liu, J.X.; Wang, Y.; Fan, M.C.; Qian, H.F.; Li, Y.; et al. Oat beta-glucan alleviates DSS-induced colitis via regulating gut microbiota metabolism in mice. Food Funct. 2021, 12, 8976–8993. [Google Scholar] [CrossRef] [PubMed]
- Wilczak, J.; Blaszczyk, K.; Kamola, D.; Gajewska, M.; Harasym, J.P.; Jalosinska, M.; Gudej, S.; Suchecka, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. The effect of low or high molecular weight oat beta-glucans on the inflammatory and oxidative stress status in the colon of rats with LPS-induced enteritis. Food Funct. 2015, 6, 590–603. [Google Scholar] [CrossRef]
- Menon, R.; Gonzalez, T.; Ferruzzi, M.; Jackson, E.; Winderl, D.; Watson, J. Oats-From Farm to Fork. Adv. Food Nutr. Res. 2016, 77, 1–55. [Google Scholar] [CrossRef]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.B. Effect of oat hulls as a free choice feeding on broiler performance, short chain fatty acids and microflora under a mild necrotic enteritis challenge. Anim. Nutr. 2018, 4, 65–72. [Google Scholar] [CrossRef]
- Tian, M.L.; Li, D.T.; Ma, C.; Feng, Y.; Hu, X.S.; Chen, F. Barley Leaf Insoluble Dietary Fiber Alleviated Dextran Sulfate Sodium-Induced Mice Colitis by Modulating Gut Microbiota. Nutrients 2021, 13, 846. [Google Scholar] [CrossRef] [PubMed]
- Li, D.T.; Feng, Y.; Tian, M.L.; Ji, J.F.; Hu, X.S.; Chen, F. Gut microbiota-derived inosine from dietary barley leaf supplementation attenuates colitis through PPAR gamma signaling activation. Microbiome 2021, 9, 1–22. [Google Scholar] [CrossRef]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [Green Version]
- Rose, D.J. Impact of whole grains on the gut microbiota: The next frontier for oats? Br. J. Nutr. 2014, 112, S44–S49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhang, Y.; Qiu, B.; Fan, S.J.; Ding, H.F.; Liu, Z.H. Quinoa whole grain diet compromises the changes of gut microbiota and colonic colitis induced by dextran Sulfate sodium in C57BL/6 mice. Sci. Rep. 2018, 8, 14916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, W.; Wang, X.; Ding, C.; Liu, J.Z.; Li, Y.; Li, W.J.; Sun, Y.D. High-Salt Diet-Induced Gastritis in C57BL/6 Mice is Associated with Microbial Dysbiosis and Alleviated by a Buckwheat Diet. Mol. Nutr. Food Res. 2020, 64, e1900965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, W.; Zhang, D.; Yang, Y.; Wang, X.; Li, L. Fermented and Germinated Processing Improved the Protective Effects of Foxtail Millet Whole Grain Against Dextran Sulfate Sodium-Induced Acute Ulcerative Colitis and Gut Microbiota Dysbiosis in C57BL/6 Mice. Front. Nutr. 2021, 8, 694936. [Google Scholar] [CrossRef]
- Nadeem, M.S.; Kumar, V.; Al-Abbasi, F.A.; Kamal, M.A.; Anwar, F. Risk of colorectal cancer in inflammatory bowel diseases. Semin. Cancer Biol. 2020, 64, 51–60. [Google Scholar] [CrossRef]
- Zhang, B.W.; Xu, Y.C.; Liu, S.; Lv, H.; Hu, Y.Z.; Wang, Y.Y.; Li, Z.; Wang, J.; Ji, X.M.; Ma, H.; et al. Dietary Supplementation of Foxtail Millet Ameliorates Colitis-Associated Colorectal Cancer in Mice via Activation of Gut Receptors and Suppression of the STAT3 Pathway. Nutrients 2020, 12, 2367. [Google Scholar] [CrossRef]
- Yang, J.; McDowell, A.; Kim, E.K.; Seo, H.; Lee, W.H.; Moon, C.M.; Kym, S.M.; Lee, D.H.; Park, Y.S.; Jee, Y.K.; et al. Development of a colorectal cancer diagnostic model and dietary risk assessment through gut microbiome analysis. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.P.; Shan, S.H.; Zhang, C.; Shi, J.Y.; Li, H.Q.; Li, Z.Y. Inhibitory Effects of Bound Polyphenol from Foxtail Millet Bran on Colitis-Associated Carcinogenesis by the Restoration of Gut Microbiota in a Mice Model. J. Agric. Food Chem. 2020, 68, 3506–3517. [Google Scholar] [CrossRef]
- He, B.B.; Bai, Y.; Jiang, L.L.; Wang, W.; Li, T.T.; Liu, P.; Tao, S.Y.; Zhao, J.C.; Han, D.D.; Wang, J.J. Effects of Oat Bran on Nutrient Digestibility, Intestinal Microbiota, and Inflammatory Responses in the Hindgut of Growing Pigs. Int. J. Mol. Sci. 2018, 19, 2407. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Miyoshi, T.; Yunoki, K.; Ito, H. Postprandial hyperlipidemia as a potential residual risk factor. J. Cardiol. 2016, 67, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Ling, Y.; Shi, Z.; Yang, X.L.; Cai, Z.W.; Wang, L.X.; Wu, X.M.; Ye, A.Q.; Jiang, J.P. Hypolipidemic effect of pure total flavonoids from peel of Citrus (PTFC) on hamsters of hyperlipidemia and its potential mechanism. Exp. Gerontol. 2020, 130, 110786. [Google Scholar] [CrossRef] [PubMed]
- Magni, P.; Macchi, C.; Morlotti, B.; Sirtori, C.R.; Ruscica, M. Risk identification and possible countermeasures for muscle adverse effects during statin therapy. Eur. J. Intern. Med. 2015, 26, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Osto, M.; Geurts, L.; Everard, A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes 2012, 3, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.P.; Tang, K.R.; Deng, Y.J.; Chen, R.S.; Liang, S.; Xie, H.J.; He, Y.F.; Chen, Y.N.; Yang, Q.H. Effects of shenling baizhu powder herbal formula on intestinal microbiota in high-fat diet-induced NAFLD rats. Biomed. Pharm. 2018, 102, 1025–1036. [Google Scholar] [CrossRef]
- Han, F.; Wang, Y.; Han, Y.Y.; Zhao, J.X.; Han, F.L.; Song, G.; Jiang, P.; Miao, H.J. Effects of Whole-Grain Rice and Wheat on Composition of Gut Microbiota and Short-Chain Fatty Acids in Rats. J. Agric. Food Chem. 2018, 66, 6326–6335. [Google Scholar] [CrossRef]
- Marshall, S.; Petocz, P.; Duve, E.; Abbott, K.; Cassettari, T.; Blumfield, M.; Fayet-Moore, F. The Effect of Replacing Refined Grains with Whole Grains on Cardiovascular Risk Factors: A Systematic Review and Meta-Analysis of Randomized Controlled Trials with GRADE Clinical Recommendation. J. Acad. Nutr. Diet 2020, 120, 1859–1883.e31. [Google Scholar] [CrossRef]
- Duan, R.Q.; Guan, X.; Huang, K.; Zhang, Y.; Li, S.; Xia, J.A.; Shen, M. Flavonoids from Whole-Grain Oat Alleviated High-Fat Diet-Induced Hyperlipidemia via Regulating Bile Acid Metabolism and Gut Microbiota in Mice. J. Agric. Food Chem. 2021, 69, 7629–7640. [Google Scholar] [CrossRef]
- Huang, K.; Yu, W.W.; Li, S.; Guan, X.; Liu, J.; Song, H.D.; Liu, D.D.; Duan, R.Q. Effect of embryo-remaining oat rice on the lipid profile and intestinal microbiota in high-fat diet fed rats. Food Res. Int. 2020, 129, 108816. [Google Scholar] [CrossRef]
- Zhang, L.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Smith, P.T.; Ananthan, R.; Longvah, T. Antioxidant and immunomodulatory activities of polysaccharides from the roots of Sanguisorba officinalis. Int. J. Biol. Macromol. 2012, 51, 1057–1062. [Google Scholar] [CrossRef]
- Cao, Y.N.; Zou, L.; Li, W.; Song, Y.; Zhao, G.; Hu, Y.C. Dietary quinoa (Chenopodium quinoa Willd.) polysaccharides ameliorate high-fat diet-induced hyperlipidemia and modulate gut microbiota. Int. J. Biol. Macromol. 2020, 163, 55–65. [Google Scholar] [CrossRef]
- Fotschki, B.; Juskiewicz, J.; Jurgonski, A.; Amarowicz, R.; Opyd, P.; Bez, J.; Muranyi, I.; Petersen, I.L.; Llopis, M.L. Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats. Nutrients 2020, 12, 2781. [Google Scholar] [CrossRef]
- Riviere, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]
- Marinelli, L.; Martin-Gallausiaux, C.; Bourhis, J.M.; Beguet-Crespel, F.; Blottiere, H.M.; Lapaque, N. Identification of the novel role of butyrate as AhR ligand in human intestinal epithelial cells. Sci. Rep.-UK 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.L.; Yan, B.B.; Xiao, Y.; Zhou, Y.M.; Liu, T.Y. Tartary buckwheat protein prevented dyslipidemia in high-fat diet-fed mice associated with gut microbiota changes. Food Chem. Toxicol. 2018, 119, 296–301. [Google Scholar] [CrossRef]
- Rivera-Espinoza, Y.; Gallardo-Navarro, Y. Non-dairy probiotic products. Food Microbiol. 2010, 27, 1–11. [Google Scholar] [CrossRef]
- Zhou, Y.; Jiang, Q.; Zhao, S.; Yan, B.; Zhou, X. Impact of Buckwheat Fermented Milk Combined with High-Fat Diet on Rats’ Gut Microbiota and Short-Chain Fatty Acids. J. Food Sci. 2019, 84, 3833–3842. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.; Itoh, K. Intestinal colonization by a Lachnospiraceae bacterium contributes to the development of diabetes in obese mice. Microbes Environ. 2014, 29, 427–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J.W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, S.A.; Kamil, A.; Fleige, L.; Gahan, C.G.M. The Cholesterol-Lowering Effect of Oats and Oat Beta Glucan: Modes of Action and Potential Role of Bile Acids and the Microbiome. Front. Nutr. 2019, 6, 171. [Google Scholar] [CrossRef]
- Sun, N.X.; Tong, L.T.; Liang, T.T.; Wang, L.L.; Liu, L.Y.; Zhou, X.R.; Zhou, S.M. Effect of Oat and Tartary Buckwheat-Based Food on Cholesterol—Lowering and Gut Microbiota in Hypercholesterolemic Hamsters. J. Oleo Sci. 2019, 68, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.L.; Hergert, N.; Rompato, G.; Lefevre, M. Whole grain oats improve insulin sensitivity and plasma cholesterol profile and modify gut microbiota composition in C57BL/6J mice. J. Nutr. 2015, 145, 222–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, M.L.; Tzounis, X.; Tuohy, K.M.; Lovegrove, J.A. Hypocholesterolemic and Prebiotic Effects of a Whole-Grain Oat-Based Granola Breakfast Cereal in a Cardio-Metabolic “At Risk” Population. Front. Microbiol. 2016, 7, 1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Chen, C.A.; Tsai, J.S.; Chen, G.W. Preparation and Identification of Novel Antihypertensive Peptides from the In Vitro Gastrointestinal Digestion of Marine Cobia Skin Hydrolysates. Nutrients 2019, 11, 1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Himmelfarb, D.C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: Executive Summary: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Hypertension 2018, 71, 1269–1324. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Cui, L.; Qi, J.; Ojo, O.; Du, X.; Liu, Y.; Wang, X. The effect of dietary fiber (oat bran) supplement on blood pressure in patients with essential hypertension: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2458–2470. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Hereu, M.; Atienza, L.; Amezqueta, S.; Casas, J.; Munoz, S.; Medina, I.; Miralles-Perez, B.; Romeu, M.; Torres, J.L. The Buckwheat Iminosugar d-Fagomine Attenuates Sucrose-Induced Steatosis and Hypertension in Rats. Mol. Nutr. Food Res. 2020, 64, e1900564. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Hao, Y.; Fan, X.; Richel, A.; Everaert, N.; Yang, X.; Ren, G. Administration with Quinoa Protein Reduces the Blood Pressure in Spontaneously Hypertensive Rats and Modifies the Fecal Microbiota. Nutrients 2021, 13, 2446. [Google Scholar] [CrossRef]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Sakanoi, Y.; Yamamoto, K.; Ota, T.; Seki, K.; Imai, M.; Ota, R.; Asayama, Y.; Nakashima, A.; Suzuki, K.; Tsuduki, T.; et al. Simultaneous Intake of Euglena Gracilis and Vegetables Synergistically Exerts an Anti-Inflammatory Effect and Attenuates Visceral Fat Accumulation by Affecting Gut Microbiota in Mice. Nutrients 2018, 10, 1417. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Guo, W.L.; Li, Q.Y.; Xu, J.X.; Cao, Y.J.; Liu, B.; Yu, X.D.; Rao, P.F.; Ni, L.; Lv, X.C. The protective mechanism of Lactobacillus plantarum FZU3013 against non-alcoholic fatty liver associated with hyperlipidemia in mice fed a high-fat diet. Food Funct. 2020, 11, 3316–3331. [Google Scholar] [CrossRef]
- Huang, Z.R.; Deng, J.C.; Li, Q.Y.; Cao, Y.J.; Lin, Y.C.; Bai, W.D.; Liu, B.; Rao, P.F.; Ni, L.; Lv, X.C. Protective Mechanism of Common Buckwheat (Fagopyrum esculentum Moench.) against Nonalcoholic Fatty Liver Disease Associated with Dyslipidemia in Mice Fed a High-Fat and High-Cholesterol Diet. J. Agric. Food Chem. 2020, 68, 6530–6543. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; Kim, J.S.; Kang, Y.M.; Lim, J.H.; Kim, Y.M.; Lee, M.S.; Jeong, M.H.; Ahn, C.B.; Je, J.Y. Antioxidant activity and gamma-aminobutyric acid (GABA) content in sea tangle fermented by Lactobacillus brevis BJ20 isolated from traditional fermented foods. Food Chem. 2010, 122, 271–276. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, Z.; Guo, H.; He, D.; Zhao, H.; Wang, Z.; Zhang, W.; Liao, L.; Zhang, C.; Ni, L. The modulatory effect of infusions of green tea, oolong tea, and black tea on gut microbiota in high-fat-induced obese mice. Food Funct. 2016, 7, 4869–4879. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, K.L.; Wood, D.L.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bei, Q.; Liu, Y.; Wang, L.; Chen, G.; Wu, Z.Q. Improving free, conjugated, and bound phenolic fractions in fermented oats (Avena sativa L.) with Monascus anka and their antioxidant activity. J. Funct. Foods 2017, 32, 185–194. [Google Scholar] [CrossRef]
- Huang, Z.R.; Chen, M.; Guo, W.L.; Li, T.T.; Liu, B.; Bai, W.D.; Ai, L.Z.; Rao, P.F.; Ni, L.; Lv, X.C. Monascus purpureus-fermented common buckwheat protects against dyslipidemia and non-alcoholic fatty liver disease through the regulation of liver metabolome and intestinal microbiome. Food Res. Int. 2020, 136, 109511. [Google Scholar] [CrossRef]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef]
- Weber, C.; Noels, H. Atherosclerosis: Current pathogenesis and therapeutic options. Nat. Med. 2011, 17, 1410–1422. [Google Scholar] [CrossRef]
- Kasikara, C.; Doran, A.C.; Cai, B.S.; Tabas, I. The role of non-resolving inflammation in atherosclerosis. J. Clin. Investig. 2018, 128, 2713–2723. [Google Scholar] [CrossRef] [Green Version]
- Lutgens, E.; Atzler, D.; Doring, Y.; Duchene, J.; Steffens, S.; Weber, C. Immunotherapy for cardiovascular disease. Eur. Heart J. 2019, 40, 3937–3946. [Google Scholar] [CrossRef]
- Liu, F.M.; Shan, S.H.; Li, H.Q.; Shi, J.Y.; Hao, R.L.; Yang, R.P.; Li, Z.Y. Millet shell polyphenols prevent atherosclerosis by protecting the gut barrier and remodeling the gut microbiota in ApoE(-/-) mice. Food Funct. 2021, 12, 7298–7309. [Google Scholar] [CrossRef]
- De Oliveira, J.; Hort, M.A.; Moreira, E.L.G.; Glaser, V.; Ribeiro-do-Valle, R.M.; Prediger, R.D.; Farina, M.; Latini, A.; de Bem, A.F. Positive Correlation between Elevated Plasma Cholesterol Levels and Cognitive Impairments in Ldl Receptor Knockout Mice: Relevance of Cortico-Cerebral Mitochondrial Dysfunction and Oxidative Stress. Neuroscience 2011, 197, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Song, R.; Li, Y.; Zhang, W.; Wan, Z.; Wang, Y.; Zhang, H.; Han, S. Effects of Oat Fiber Intervention on Cognitive Behavior in LDLR(-/-) Mice Modeling Atherosclerosis by Targeting the Microbiome-Gut-Brain Axis. J. Agric. Food Chem. 2020, 68, 14480–14491. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.H.; Wu, S.S.; Xia, Y.; Huang, J.Z.; Ye, J.F.; Xuan, Z.N.; Li, P.; Du, B. Probiotic-fermented black tartary buckwheat alleviates hyperlipidemia and gut microbiota dysbiosis in rats fed with a high-fat diet. Food Funct. 2021, 12, 6045–6057. [Google Scholar] [CrossRef]
- Sarma, S.M.; Singh, D.P.; Singh, P.; Khare, P.; Mangal, P.; Singh, S.; Bijalwan, V.; Kaur, J.; Mantri, S.; Boparai, R.K.; et al. Finger millet arabinoxylan protects mice from high-fat diet induced lipid derangements, inflammation, endotoxemia and gut bacterial dysbiosis. Int. J. Biol. Macromol. 2018, 106, 994–1003. [Google Scholar] [CrossRef]
- Li, S.; Liang, T.; Zhang, Y.; Huang, K.; Yang, S.Y.; Lv, H.Y.; Chen, Y.; Zhang, C.H.; Guan, X. Vitexin alleviates high-fat diet induced brain oxidative stress and inflammation via anti-oxidant, anti-inflammatory and gut microbiota modulating properties. Free Radic. Biol. Med. 2021, 171, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.M.; London, L.E.E.; Bjorndahl, T.C.; Mandal, R.; Murphy, K.; Fitzgerald, G.F.; Shanahan, F.; Ross, R.P.; Wishart, D.S.; Caplice, N.M.; et al. Microbiome and metabolome modifying effects of several cardiovascular disease interventions in apo-E-/- mice. Microbiome 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, H.; Lim, J.; Lee, H.; Yoon, S.; Shin, S.S.; Yoon, M. The lemon balm extract ALS-L1023 inhibits obesity and nonalcoholic fatty liver disease in female ovariectomized mice. Food Chem. Toxicol. 2017, 106 Pt A, 292–305. [Google Scholar] [CrossRef]
- Xiao, X.; Clark, J.M.; Park, Y. Potential contribution of insecticide exposure and development of obesity and type 2 diabetes. Food Chem. Toxicol. 2017, 105, 456–474. [Google Scholar] [CrossRef]
- Gasparotto, J.; Chaves, P.R.; da Boit Martinello, K.; Silva Oliveira, L.F.; Gelain, D.P.; Fonseca Moreira, J.C. Obesity associated with coal ash inhalation triggers systemic inflammation and oxidative damage in the hippocampus of rats. Food Chem. Toxicol. 2019, 133, 110766. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, D.; Ke, W.; Liang, D.; Hu, X.; Chen, F. Resveratrol-induced gut microbiota reduces obesity in high-fat diet-fed mice. Int. J. Obes. 2020, 44, 213–225. [Google Scholar] [CrossRef]
- Ji, Y.; Ma, N.; Zhang, J.M.; Wang, H.T.; Tao, T.Y.; Pei, F.; Hu, Q.H. Dietary intake of mixture coarse cereals prevents obesity by altering the gut microbiota in high-fat diet fed mice. Food Chem. Toxicol. 2021, 147, 111901. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Mills, D.; Noratto, G. Molecular exploration of fecal microbiome in quinoa-supplemented obese mice. FEMS Microbiol. Ecol. 2016, 92, fiw089. [Google Scholar] [CrossRef] [Green Version]
- Presley, L.L.; Wei, B.; Braun, J.; Borneman, J. Bacteria Associated with Immunoregulatory Cells in Mice. Appl. Environ. Microbol. 2010, 76, 936–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breton, J.; Massart, S.; Vandamme, P.; De Brandt, E.; Pot, B.; Foligne, B. Ecotoxicology inside the gut: Impact of heavy metals on the mouse microbiome. BMC Pharm. Toxicol. 2013, 14, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mazcorro, J.F.; Mills, D.A.; Murphy, K.; Noratto, G. Effect of barley supplementation on the fecal microbiota, caecal biochemistry, and key biomarkers of obesity and inflammation in obese db/db mice. Eur. J. Nutr. 2018, 57, 2513–2528. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Ivanov, I.; Mills, D.A.; Noratto, G. Influence of whole-wheat consumption on fecal microbial community structure of obese diabetic mice. PeerJ 2016, 4, e1702. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.X.; Wang, T.X.; Sun, C.; Wang, J.; Sun, B.G. Whole barley prevents obesity and dyslipidemia without the involvement of the gut microbiota in germ free C57BL/6J obese mice. Food Funct. 2019, 10, 7498–7508. [Google Scholar] [CrossRef]
- Zhong, Y.; Nyman, M.; Fak, F. Modulation of gut microbiota in rats fed high-fat diets by processing whole-grain barley to barley malt. Mol. Nutr. Food Res. 2015, 59, 2066–2076. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Song, J.; Li, J.; Wang, H.; Zhang, Y.; Suo, H. A synbiotic consisting of Lactobacillus plantarum S58 and hull-less barley beta-glucan ameliorates lipid accumulation in mice fed with a high-fat diet by activating AMPK signaling and modulating the gut microbiota. Carbohydr. Polym. 2020, 243, 116398. [Google Scholar] [CrossRef] [PubMed]
- Przybylski, P.; Pyta, K.; Ratajczak-Sitarz, M.; Katrusiak, A.; Brzezinski, B. Structure of a New Schiff Base of Gossypol with Ethyl 4-Amino-1-piperidine Carboxylate in the Solid and in the Solution. Pol. J. Chem. 2009, 83, 747–759. [Google Scholar]
- Hereu, M.; Ramos-Romero, S.; Garcia-Gonzalez, N.; Amezqueta, S.; Torres, J.L. Eubiotic effect of buckwheat D-fagomine in healthy rats. J. Funct. Foods 2018, 50, 120–126. [Google Scholar] [CrossRef]
- Poudyal, H.; Panchal, S.K.; Diwan, V.; Brown, L. Omega-3 fatty acids and metabolic syndrome: Effects and emerging mechanisms of action. Prog. Lipid Res. 2011, 50, 372–387. [Google Scholar] [CrossRef]
- Mujico, J.R.; Baccan, G.C.; Gheorghe, A.; Diaz, L.E.; Marcos, A. Changes in gut microbiota due to supplemented fatty acids in diet-induced obese mice. Br. J. Nutr. 2013, 110, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Robertson, R.C.; Oriach, C.S.; Murphy, K.; Moloney, G.M.; Cryan, J.F.; Dinan, T.G.; Ross, R.P.; Stanton, C. Omega-3 polyunsaturated fatty acids critically regulate behaviour and gut microbiota development in adolescence and adulthood. Brain Behav. Immun. 2017, 59, 21–37. [Google Scholar] [CrossRef]
- Hereu, M.; Ramos-Romero, S.; Marin-Valls, R.; Amezqueta, S.; Miralles-Perez, B.; Romeu, M.; Mendez, L.; Medina, I.; Torres, J.L. Combined Buckwheat D-Fagomine and Fish Omega-3 PUFAs Stabilize the Populations of Gut Prevotella and Bacteroides While Reducing Weight Gain in Rats. Nutrients 2019, 11, 2606. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Zhang, Q.; Zhang, Y.; Yao, Z.; Song, P.; Wei, L.; Zhao, G.; Yan, Z. Effect of tartary buckwheat, rutin, and quercetin on lipid metabolism in rats during high dietary fat intake. Food Sci. Nutr. 2020, 8, 199–213. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, S.; Jiang, Y.; Wei, Y.; Zhou, X. Regulatory Function of Buckwheat-Resistant Starch Supplementation on Lipid Profile and Gut Microbiota in Mice Fed with a High-Fat Diet. J. Food Sci. 2019, 84, 2674–2681. [Google Scholar] [CrossRef]
- Zhou, Y.; Wei, Y.; Yan, B.; Zhao, S.; Zhou, X. Regulation of tartary buckwheat-resistant starch on intestinal microflora in mice fed with high-fat diet. Food Sci. Nutr. 2020, 8, 3243–3251. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.M.; Khare, P.; Jagtap, S.; Singh, D.P.; Baboota, R.K.; Podili, K.; Boparai, R.K.; Kaur, J.; Bhutani, K.K.; Bishnoi, M.; et al. Kodo millet whole grain and bran supplementation prevents high-fat diet induced derangements in a lipid profile, inflammatory status and gut bacteria in mice. Food Funct. 2017, 8, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Awika, J.M.; McDonough, C.M.; Rooney, L.W. Decorticating sorghum to concentrate healthy phytochemicals. J. Agric. Food Chem. 2005, 53, 6230–6234. [Google Scholar] [CrossRef]
- Ashley, D.; Marasini, D.; Brownmiller, C.; Lee, J.A.; Carbonero, F.; Lee, S.O. Impact of Grain Sorghum Polyphenols on Microbiota of Normal Weight and Overweight/Obese Subjects during In Vitro Fecal Fermentation. Nutrients 2019, 11, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Yang, C.; Wai, S.T.C.; Zhang, Y.; Portillo, M.P.; Paoli, P.; Wu, Y.; San Cheang, W.; Liu, B.; Carpene, C.; et al. Regulation of glucose metabolism by bioactive phytochemicals for the management of type 2 diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2019, 59, 830–847. [Google Scholar] [CrossRef]
- Della Pepa, G.; Vetrani, C.; Vitale, M.; Riccardi, G. Wholegrain Intake and Risk of Type 2 Diabetes: Evidence from Epidemiological and Intervention Studies. Nutrients 2018, 10, 1288. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Wang, L.X.; Chen, Z.L.; Hou, D.A.Z.; Xue, Y.; Diao, X.M.; Shen, Q. Foxtail Millet Improves Blood Glucose Metabolism in Diabetic Rats through PI3K/AKT and NF-kappa B Signaling Pathways Mediated by Gut Microbiota. Nutrients 2021, 13, 1837. [Google Scholar] [CrossRef]
- Fu, Y.X.; Yin, R.Y.; Guo, E.H.; Cheng, R.H.; Diao, X.M.; Xue, Y.; Shen, Q. Protein Isolates from Raw and Cooked Foxtail Millet Attenuate Development of Type 2 Diabetes in Streptozotocin-Induced Diabetic Mice. Mol. Nutr. Food Res. 2021, 65, 2000365. [Google Scholar] [CrossRef]
- Pascale, A.; Marchesi, N.; Govoni, S.; Coppola, A.; Gazzaruso, C. The role of gut microbiota in obesity, diabetes mellitus, and effect of metformin: New insights into old diseases. Curr. Opin. Pharm. 2019, 49, 1–5. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Gallego, C.; Collado, M.C.; Perez, G.; Ilo, T.; Jaakkola, U.M.; Bernal, M.J.; Periago, M.J.; Frias, R.; Ros, G.; Salminen, S. Resembling breast milk: Influence of polyamine-supplemented formula on neonatal BALB/cOlaHsd mouse microbiota. Br. J. Nutr. 2014, 111, 1050–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vojinovic, D.; Radjabzadeh, D.; Kurilshikov, A.; Amin, N.; Wijmenga, C.; Franke, L.; Ikram, M.A.; Uitterlinden, A.G.; Zhernakova, A.; Fu, J.; et al. Relationship between gut microbiota and circulating metabolites in population-based cohorts. Nat. Commun. 2019, 10, 5813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenberg, C.; Savage, D.B. An amino acid profile to predict diabetes? Nat. Med. 2011, 17, 418–420. [Google Scholar] [CrossRef]
- Chang, D.H.; Rhee, M.S.; Ahn, S.; Bang, B.H.; Oh, J.E.; Lee, H.K.; Kim, B.C. Faecalibaculum rodentium gen. nov. sp. nov. isolated from the faeces of a laboratory mouse. Antonie Van Leeuwenhoek 2015, 108, 1309–1318. [Google Scholar] [CrossRef]
- Fu, Y.X.; Yin, R.Y.; Liu, Z.Y.; Niu, Y.; Guo, E.H.; Cheng, R.H.; Diao, X.M.; Xue, Y.; Shen, Q. Hypoglycemic Effect of Prolamin from Cooked Foxtail Millet (Setaria italic) on Streptozotocin-Induced Diabetic Mice. Nutrients 2020, 12, 3452. [Google Scholar] [CrossRef]
- Zheng, J.P.; Zhu, L.; Hu, B.F.; Zou, X.J.; Hu, H.M.; Zhang, Z.G.; Jiang, N.; Ma, J.; Yang, H.B.; Liu, H.T. 1-Deoxynojirimycin improves high fat diet-induced nonalcoholic steatohepatitis by restoring gut dysbiosis. J. Nutr. Biochem. 2019, 71, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.M.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; Francois, P.; de Vos, W.M.; et al. Responses of Gut Microbiota and Glucose and Lipid Metabolism to Prebiotics in Genetic Obese and Diet-Induced Leptin-Resistant Mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lian, F.M.; Zhao, L.H.; Zhao, Y.F.; Chen, X.Y.; Zhang, X.; Guo, Y.; Zhang, C.H.; Zhou, Q.; Xue, Z.S.; et al. Structural modulation of gut microbiota during alleviation of type 2 diabetes with a Chinese herbal formula. ISME J. 2015, 9, 552–562. [Google Scholar] [CrossRef]
- Zhou, W.; Sailani, M.R.; Contrepois, K.; Zhou, Y.; Ahadi, S.; Leopold, S.R.; Zhang, M.J.; Rao, V.; Avina, M.; Mishra, T.; et al. Longitudinal multi-omics of host-microbe dynamics in prediabetes. Nature 2019, 569, 663–671. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Q.; Liu, M.; Zhang, X.; He, F.; Wang, G. Akkermansia muciniphila can reduce the damage of gluco/lipotoxicity, oxidative stress and inflammation, and normalize intestine microbiota in streptozotocin-induced diabetic rats. Pathog. Dis. 2018, 76, fty028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhao, Y.F.; Xu, J.; Xue, Z.S.; Zhang, M.H.; Pang, X.Y.; Zhang, X.J.; Zhao, L.P. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci. Rep. 2015, 5, 14405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Romero, S.; Hereu, M.; Atienza, L.; Casas, J.; Taltavull, N.; Romeu, M.; Amezqueta, S.; Dasilva, G.; Medina, I.; Torres, J.L. Functional Effects of the Buckwheat Iminosugar D-Fagomine on Rats with Diet-Induced Prediabetes. Mol. Nutr. Food Res. 2018, 62, e1800373. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef]
- Kristek, A.; Wiese, M.; Heuer, P.; Kosik, O.; Schar, M.Y.; Soycan, G.; Alsharif, S.; Kuhnle, G.G.C.; Walton, G.; Spencer, J.P.E. Oat bran, but not its isolated bioactive beta-glucans or polyphenols, have a bifidogenic effect in an in vitro fermentation model of the gut microbiota. Br. J. Nutr. 2019, 121, 549–559. [Google Scholar] [CrossRef]
- Gullon, B.; Gullon, P.; Tavaria, F.K.; Yanez, R. Assessment of the prebiotic effect of quinoa and amaranth in the human intestinal ecosystem. Food Funct. 2016, 7, 3782–3788. [Google Scholar] [CrossRef]
- Bekedam, E.K.; Roos, E.; Schols, H.A.; van Boekel, M.A.J.S.; Smit, G. Low molecular weight melanoidins in coffee brew. J. Agric. Food Chem. 2008, 56, 4060–4067. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Bellesia, A. The gastro-intestinal tract as the major site of biological action of dietary melanoidins. Amino Acids 2015, 47, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Aljahdali, N.; Gadonna-Widehem, P.; Anton, P.M.; Carbonero, F. Gut Microbiota Modulation by Dietary Barley Malt Melanoidins. Nutrients 2020, 12, 241. [Google Scholar] [CrossRef] [Green Version]
- Voreades, N.; Kozil, A.; Weir, T.L. Diet and the development of the human intestinal microbiome. Front. Microbiol. 2014, 5, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velikonja, A.; Lipoglavsek, L.; Zorec, M.; Orel, R.; Avgustin, G. Alterations in gut microbiota composition and metabolic parameters after dietary intervention with barley beta glucans in patients with high risk for metabolic syndrome development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef]
- Deaver, J.A.; Eum, S.Y.; Toborek, M. Circadian Disruption Changes Gut Microbiome Taxa and Functional Gene Composition. Front. Microbiol. 2018, 9, 737. [Google Scholar] [CrossRef]
- Cheng, W.Y.; Lam, K.L.; Li, X.J.; Kong, A.P.S.; Cheung, P.C.K. Circadian disruption-induced metabolic syndrome in mice is ameliorated by oat beta-glucan mediated by gut microbiota. Carbohydr. Polym. 2021, 267, 118216. [Google Scholar] [CrossRef]
- Dahlhausen, K.E.; Doroud, L.; Firl, A.J.; Polkinghorne, A.; Eisen, J.A. Characterization of shifts of koala (Phascolarctos cinereus) intestinal microbial communities associated with antibiotic treatment. PeerJ 2018, 6, e4452. [Google Scholar] [CrossRef]
- Sun, L.J.; Ma, L.J.; Ma, Y.B.; Zhang, F.M.; Zhao, C.H.; Nie, Y.Z. Insights into the role of gut microbiota in obesity: Pathogenesis, mechanisms, and therapeutic perspectives. Protein Cell 2018, 9, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.Y.; Li, J.; Wei, C.H.; Luo, T.; Deng, Z.Y.; Fan, Y.W.; Zheng, L.F. A polysaccharide from Fagopyrum esculentum Moench bee pollen alleviates microbiota dysbiosis to improve intestinal barrier function in antibiotic-treated mice. Food Funct. 2020, 11, 10519–10533. [Google Scholar] [CrossRef]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J.; et al. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef] [Green Version]
- Bruce-Keller, A.J.; Salbaum, J.M.; Luo, M.; Blanchard, E.T.; Taylor, C.M.; Welsh, D.A.; Berthoud, H.R. Obese-type gut microbiota induce neurobehavioral changes in the absence of obesity. Biol. Psychiatry 2015, 77, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.L.; Yu, Y.H.; Lin, D.H.; Zheng, P.; Zhang, P.; Hu, M.M.; Wang, Q.; Pan, W.; Yang, X.Y.; Hu, T.; et al. beta-glucan attenuates cognitive impairment via the gut-brain axis in diet-induced obese mice. Microbiome 2020, 8, 1–21. [Google Scholar] [CrossRef]
- Saji, N.; Niida, S.; Murotani, K.; Hisada, T.; Tsuduki, T.; Sugimoto, T.; Kimura, A.; Toba, K.; Sakurai, T. Analysis of the relationship between the gut microbiome and dementia: A cross-sectional study conducted in Japan. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.F.; Yu, Y.H.; Beck, E.J.; South, T.; Li, Y.L.; Batterham, M.J.; Tapsell, L.C.; Chen, J.Z. Diet high in oat beta-glucan activates the gut-hypothalamic (PYY3-36-NPY) axis and increases satiety in diet-induced obesity in mice. Mol. Nutr. Food Res. 2011, 55, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Mattson, M.P. The impact of dietary energy intake on cognitive aging. Front. Aging Neurosci. 2010, 2. [Google Scholar] [CrossRef]
- Black, R.E.; Victora, C.G.; Walker, S.P.; Maternal and Child Nutrition Study Group. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 2013, 382, 396. [Google Scholar] [CrossRef]
- Li, X.J.; Hui, Y.; Leng, B.F.; Ren, J.L.; Song, Y.N.; Che, L.Q.; Peng, X.; Huang, B.J.; Liu, S.L.; Li, L.; et al. Millet-based supplement restored gut microbial diversity of acute malnourished pigs. PLoS ONE 2021, 16, e0250423. [Google Scholar] [CrossRef]
- Gehrig, J.L.; Venkatesh, S.; Chang, H.W.; Hibberd, M.C.; Kung, V.L.; Cheng, J.Y.; Chen, R.Y.; Subramanian, S.; Cowardin, C.A.; Meier, M.F.; et al. Effects of microbiota-directed foods in gnotobiotic animals and undernourished children. Science 2019, 365, eaau4732. [Google Scholar] [CrossRef] [Green Version]
- Umu, O.C.O.; Fauske, A.K.; Akesson, C.P.; de Nanclares, M.P.; Sorby, R.; Press, C.M.; Overland, M.; Sorum, H. Gut microbiota profiling in Norwegian weaner pigs reveals potentially beneficial effects of a high-fiber rapeseed diet. PLoS ONE 2018, 13, e0209439. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, G.; Fan, X.; Teng, C.; Li, Y.; Everaert, N.; Blecker, C. The Beneficial Effect of Coarse Cereals on Chronic Diseases through Regulating Gut Microbiota. Foods 2021, 10, 2891. https://doi.org/10.3390/foods10112891
Ren G, Fan X, Teng C, Li Y, Everaert N, Blecker C. The Beneficial Effect of Coarse Cereals on Chronic Diseases through Regulating Gut Microbiota. Foods. 2021; 10(11):2891. https://doi.org/10.3390/foods10112891
Chicago/Turabian StyleRen, Guixing, Xin Fan, Cong Teng, Yajie Li, Nadia Everaert, and Christophe Blecker. 2021. "The Beneficial Effect of Coarse Cereals on Chronic Diseases through Regulating Gut Microbiota" Foods 10, no. 11: 2891. https://doi.org/10.3390/foods10112891
APA StyleRen, G., Fan, X., Teng, C., Li, Y., Everaert, N., & Blecker, C. (2021). The Beneficial Effect of Coarse Cereals on Chronic Diseases through Regulating Gut Microbiota. Foods, 10(11), 2891. https://doi.org/10.3390/foods10112891