
Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
2.2. Measurement of Browning Index, Firmness, and Extractable Juice
2.3. Measurement of , Generation Rate and H2O2 Content
2.4. Measurement of 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) and Radical Scavenging Capacity
2.5. Determination of SOD and CAT Activities
2.6. Measurement of Parameters Related to the AsA-GSH Cycle
2.7. Real-Time Quantitative PCR (RT-qPCR) Analysis
2.8. Statistical Analysis
3. Results
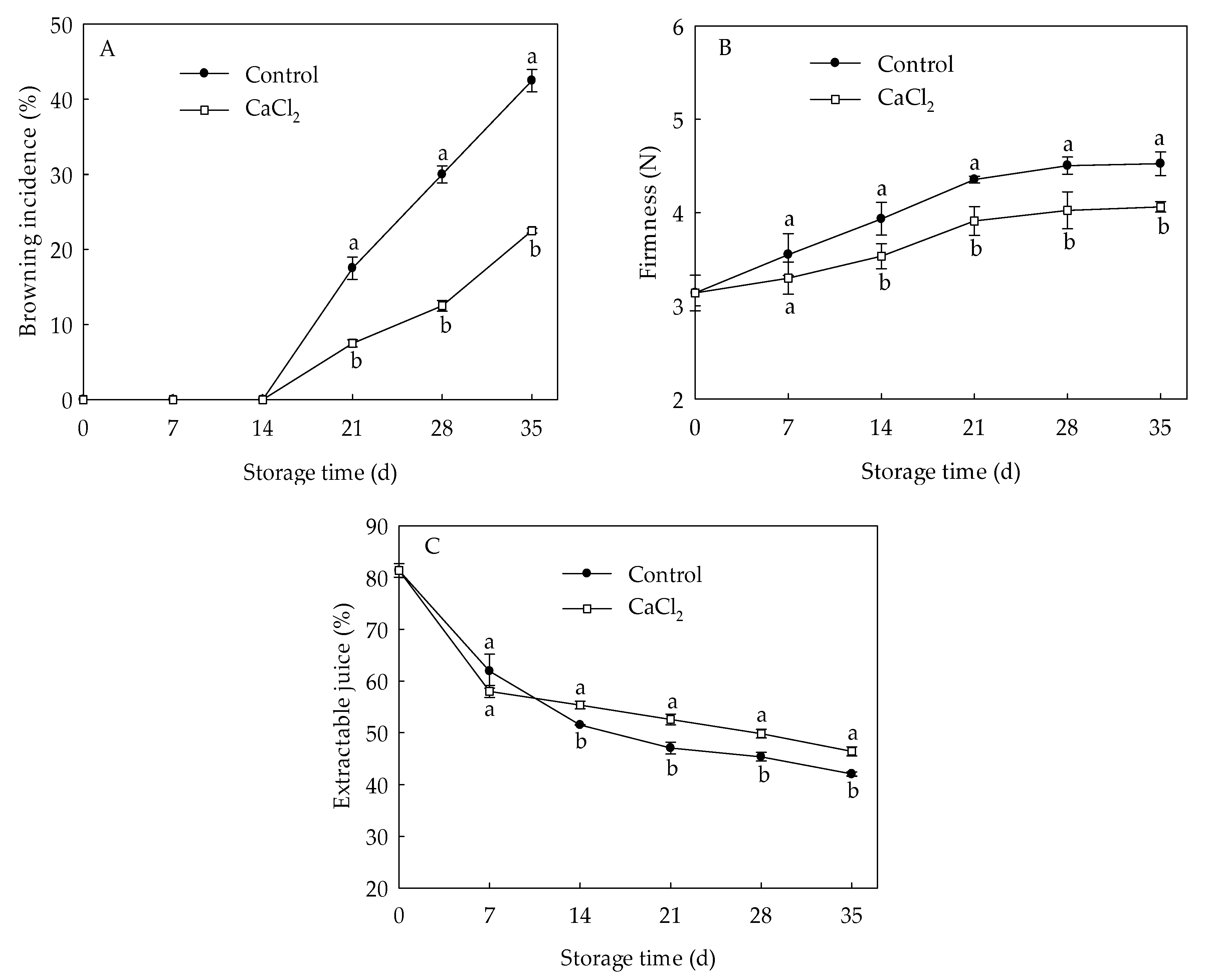
3.1. Effects of CaCl2 Treatment on Internal Browning Index, Fruit Firmness, Extractable Juice of Loquat Fruit
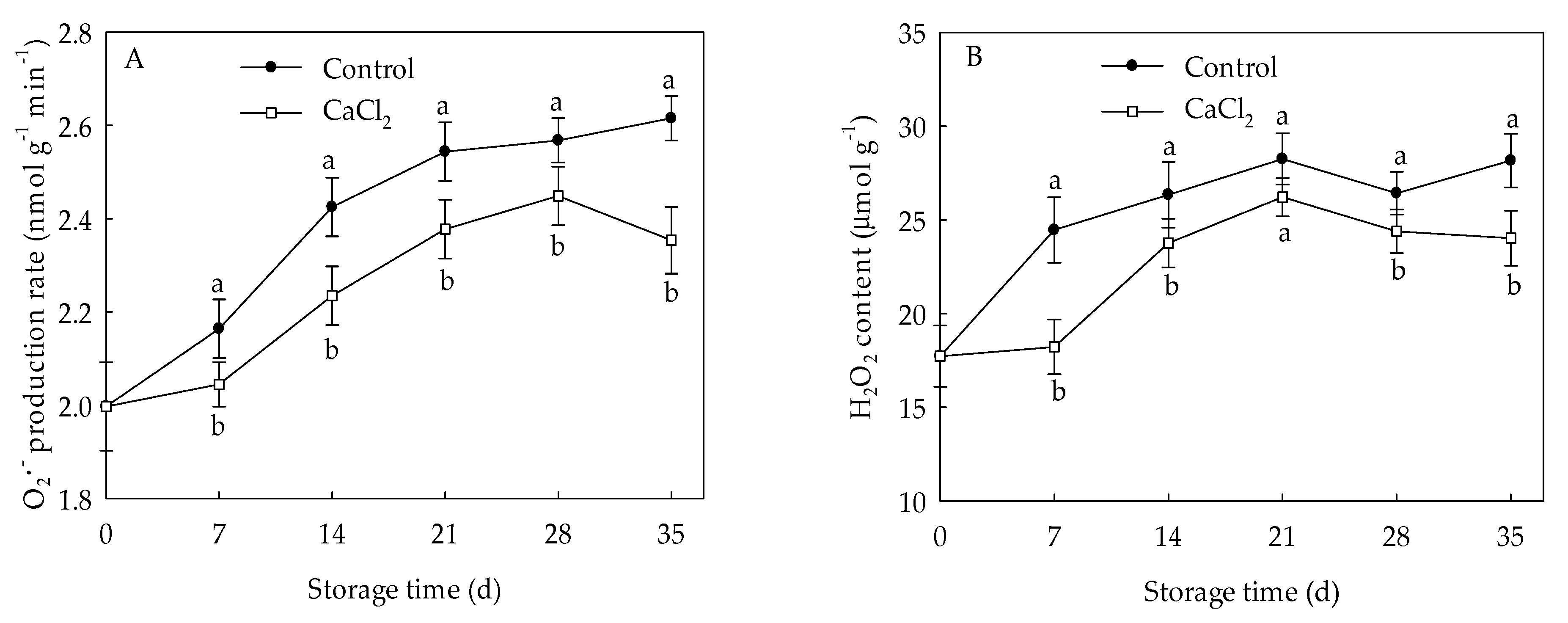
3.2. Effects of CaCl2 Treatment on Generation Rate and H2O2 Content of Loquat Fruit
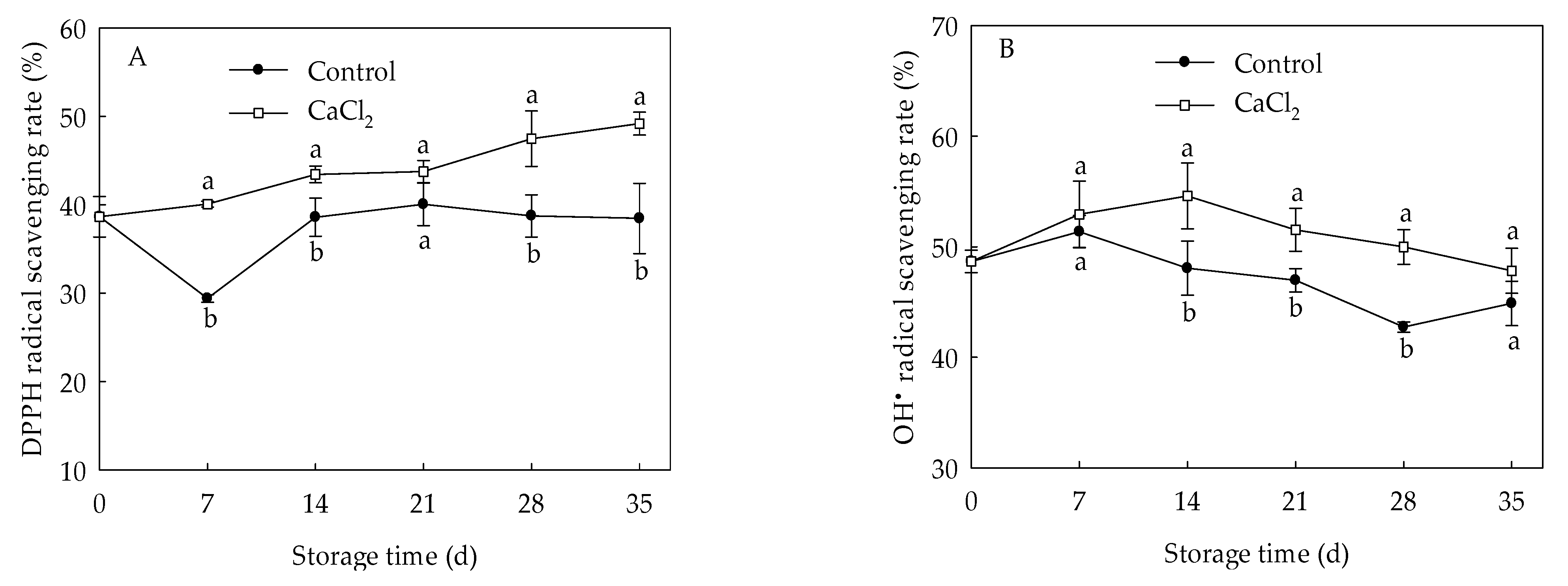
3.3. Effects of CaCl2 Treatment on DPPH and Radical Scavenging Capacity of Loquat Fruit
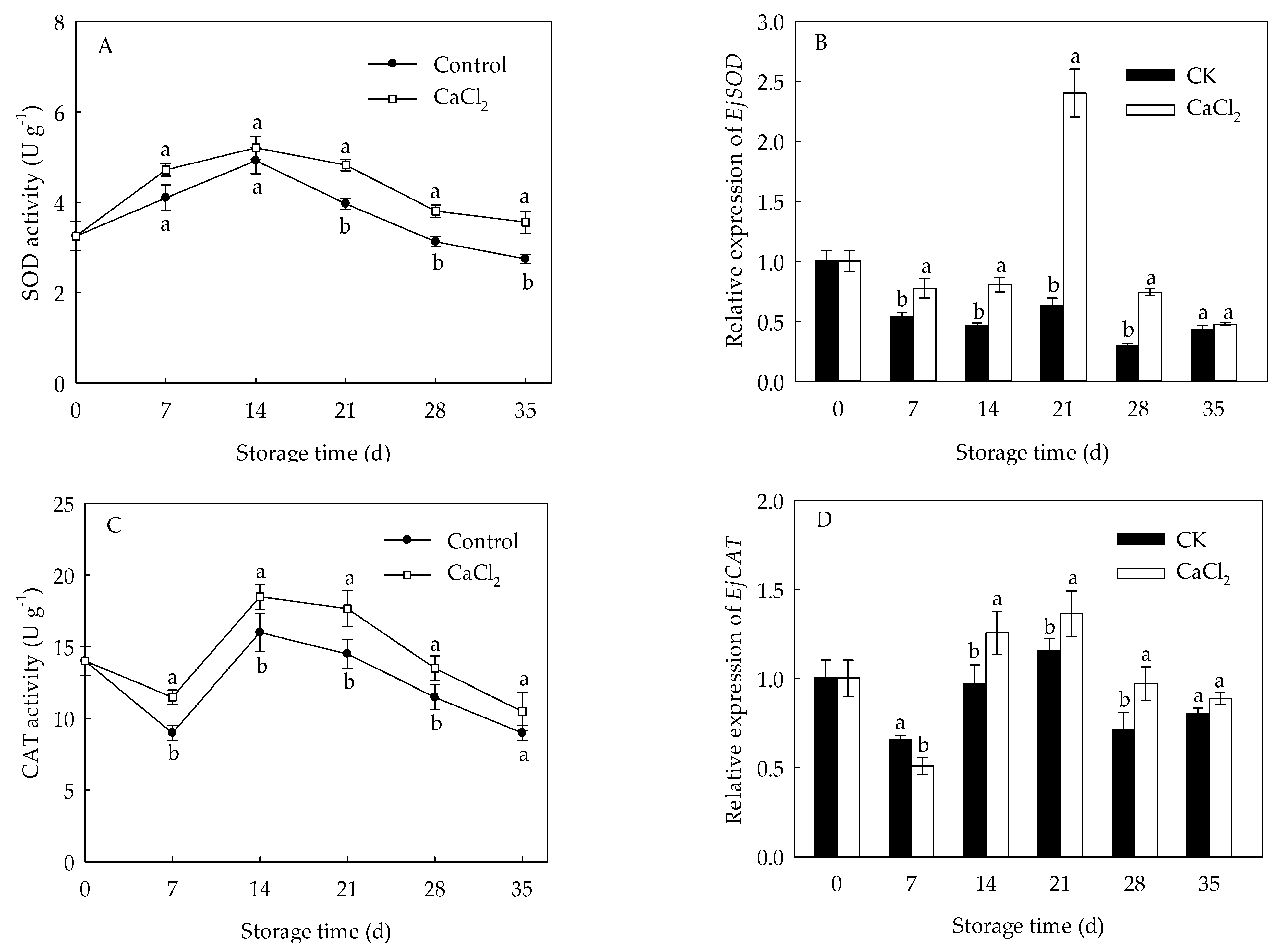
3.4. Effects of CaCl2 Treatment on CAT, SOD Activities and EjCAT, EjSOD Expressions of Loquat Fruit
3.5. Effects of CaCl2 Treatment on Contents of AsA, GSH, GSSG and GSH/GSSG Ratio of Loquat Fruit
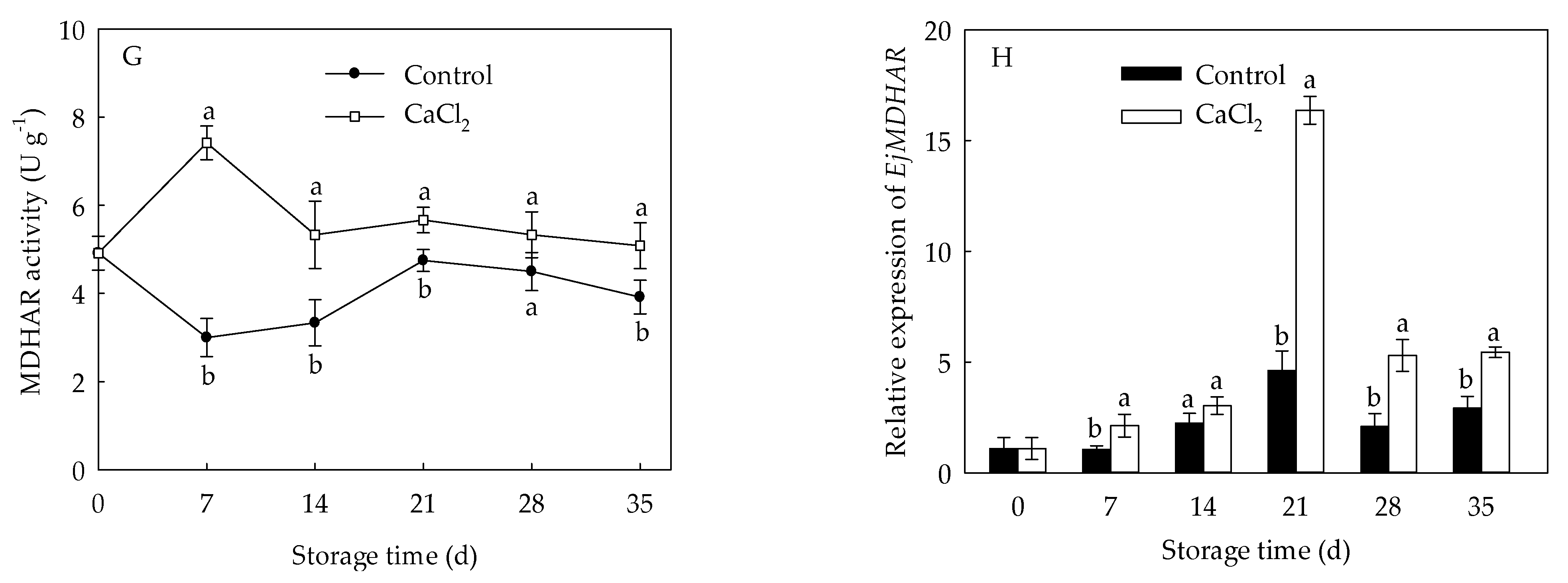
3.6. Effects of CaCl2 Treatment on Enzyme Activity and Gene Expression of APX, GR, DHAR, and MDHAR of Loquat Fruit
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, Y.H.; Li, S.Y.; Xi, Y.F. Changes of cell wall substances in relation to flesh woodiness in cold-stored loquat fruits. Acta Phytophysiol. Sin. 2000, 26, 306–310. [Google Scholar] [CrossRef]
- Cai, C.; Xu, C.J.; Li, X.; Ferguson, I.B.; Chen, K.S. Accumulation of lignin in relation to change in activities of lignification enzymes in loquat fruit flesh after harvest. Postharvest Boil. Technol. 2006, 40, 163–169. [Google Scholar] [CrossRef]
- Dreyer, A.; Dietz, K.J. Reactive Oxygen species and the redox-regulatory network in cold stress acclimation. Antioxidants 2018, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chan, Z.L. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ye, L.; Wang, Y.; Zhou, X.; Yang, J.; Wang, J.; Cao, K.; Zou, Z. Melatonin increases the chilling tolerance of chloroplast in cucumber seedlings by regulating photosynthetic electron flux and the ascorbate-glutathione cycle. Front. Plant Sci. 2016, 7, 1814. [Google Scholar] [CrossRef] [Green Version]
- Song, L.L.; Wang, J.H.; Shafi, M.; Liu, Y.; Wang, J.; Wu, J.S.; Wu, A.M. Hypobaric treatment effects on chilling injury, mitochondrial dysfunction, and the ascorbate–glutathione (AsA-GSH) cycle in postharvest peach fruit. J. Agric. Food Chem. 2016, 64, 4665–4674. [Google Scholar] [CrossRef]
- Liu, J.L.; Sun, J.H.; Pan, Y.G.; Yun, Z.; Zhang, Z.K.; Guoxiang Jiang, G.X.; Jiang, Y.M. Endogenous melatonin generation plays a positive role in chilling tolerance in relation to redox homeostasis in litchi fruit during refrigeration. Postharvest Boil. Technol. 2021, 178, 111554. [Google Scholar] [CrossRef]
- Yao, M.; Ge, W.; Zhou, Q.; Zhou, X.; Luo, M.; Zhao, Y.; Wei, B.; Ji, S. Exogenous glutathione alleviates chilling injury in postharvest bell pepper by modulating the ascorbate-glutathione (AsA-GSH) cycle. Food Chem. 2021, 352, 129458. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Yang, Z.; Cai, Y.; Zheng, Y. Fatty acid composition and antioxidant system in relation to susceptibility of loquat fruit to chilling injury. Food Chem. 2011, 127, 1777–1783. [Google Scholar] [CrossRef]
- Cai, Y.; Cao, S.; Yang, Z.; Zheng, Y. MeJA regulates enzymes involved in ascorbic acid and glutathione metabolism and improves chilling tolerance in loquat fruit. Postharvest Boil. Technol. 2011, 59, 324–326. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef] [Green Version]
- Ranty, B.; Aldon, D.; Cotelle, V.; Galaud, J.P.; Thuleau, P.; Mazars, C. Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 2016, 7, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium signaling-mediated plant response to cold stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.; Wu, C.; Luo, C.; Wei, M.; Qu, S.; Wang, S. Overexpression of MdCPK1a gene, a calcium dependent protein kinase in apple, increase tobacco cold tolerance via scavenging ROS accumulation. PLoS ONE 2020, 15, e0242139. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, Q.; Zhou, X.; Wei, B.; Ji, S. Calcium treatment alleviates pericarp browning of ‘nanguo’ pears by regulating the GABA shunt after cold storage. Front. Plant Sci. 2020, 11, 580986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, J.; Zhou, B.; Li, G.; Liu, Y.; Xia, X.L.; Xiao, Z.G.; Fei, L.; Ji, S.J. Calcium inhibited peel browning by regulating enzymes in membrane metabolism of ‘Nanguo’ pears during post-ripeness after refrigerated storage. Sci. Hortic. 2019, 244, 15–21. [Google Scholar] [CrossRef]
- Wei, D.; Zhao, X. Calcium maintained higher quality and enhanced resistance against chilling stress by regulating enzymes in reactive oxygen and biofilm metabolism of chinese winter jujube fruit. J. Food Biochem. 2020, 44, e13161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Ma, M.J.; Ye, B.; Liu, L.; Ji, S.J. Calcium ion improves cold resistance of green peppers (Capsicum annuum L.) by regulating the activity of protective enzymes and membrane lipid composition. Sci. Hortic. 2021, 277, 109789. [Google Scholar] [CrossRef]
- Li, Z.Y.; Wang, L.; Xie, B.; Hu, S.Q.; Zheng, Y.H.; Jin, P. Effects of exogenous calcium and calcium chelant on cold tolerance of postharvest loquat fruit. Sci. Hortic. 2020, 269, 109391. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, P.; Huang, Y.P.; Shan, T.M.; Wang, L.; Li, Y.Y.; Zheng, Y.H. Effect of hot water combined with glycine betaine alleviates chilling injury in cold-stored loquat fruit. Postharvest Boil. Technol. 2016, 118, 141–147. [Google Scholar] [CrossRef]
- Wang, L.; Shao, S.; Madebo, M.P.; Hou, Y.; Zheng, Y.; Jin, P. Effect of nano-SiO2 packing on postharvest quality and antioxidant capacity of loquat fruit under ambient temperature storage. Food Chem. 2020, 315, 126295. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.H.; Dhawan, P.; Math, P. A simple procedure for the isolation of high quality RNA from ripening banana fruit. Plant Mol. Biol. Rep. 2000, 18, 109–115. [Google Scholar] [CrossRef]
- Zeng, J.K.; Li, X.; Zhang, J.; Ge, H.; Yin, X.R.; Chen, K.S. Regulation of loquat fruit low temperature response and lignification involves interaction of heat shock factors and genes associated with lignin biosynthesis. Plant Cell Environ. 2016, 39, 1780–1789. [Google Scholar] [CrossRef]
- Youryon, P.; Supapvanich, S.; Kongtrakool, P.; Wongs-Aree, C. Calcium chloride and calcium gluconate peduncle infiltrations alleviate the internal browning of queen pineapple in refrigerated storage. Hortic. Environ. Biotechnol. 2018, 59, 205–213. [Google Scholar] [CrossRef]
- Habibi, F.; Ramezanian, A.; Guillén, F.; Martínez-Romero, D.; Serrano, M.; Valero, D. Susceptibility of blood orange cultivars to chilling injury based on antioxidant system, physiological and biochemical responses at different storage temperatures. Foods 2020, 9, 1609. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zheng, Y.; Wang, K.; Peng, J.; Rui, H. Methyl jasmonate reduces chilling injury and enhances antioxidant enzyme activity in postharvest loquat fruit. Food Chem. 2009, 115, 1458–1463. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sun, H.; Luo, M.L.; Zhou, X.; Zhou, Q.; Ji, S.J. Exogenous glycine betaine treatment alleviates low temperature-induced pericarp browning of ‘nanguo’ pears by regulating antioxidant enzymes and proline metabolism. Food Chem. 2019, 306, 125626. [Google Scholar] [CrossRef]
- Hao, J.; Li, X.; Xu, G.; Huo, Y.; Yang, H. Exogenous progesterone treatment alleviates chilling injury in postharvest banana fruit associated with induction of alternative oxidase and antioxidant defense. Food Chem. 2019, 286, 329–337. [Google Scholar] [CrossRef]
- Tan, X.L.; Zhao, Y.T.; Shan, W.; Kuang, J.F.; Chen, J.Y. Melatonin delays leaf senescence of postharvest chinese flowering cabbage through ros homeostasis. Food Res. Int. 2020, 138, 109790. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ye, T.; Ba, O.Z.; Liu, X.; Chan, Z. Comparative proteomic and metabolomic analyses reveal mechanisms of improved cold stress tolerance in bermudagrass (cynodon dactylon (L.) Pers.) by exogenous calcium. J. Integr. Plant Biol. 2014, 56, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Y.; Wang, Y.T.; Pan, X.B.; Xi, Z.M. Amelioration of cold-induced oxidative stress by exogenous 24-epibrassinolide treatment in grapevine seedlings: Toward regulating the ascorbate–glutathione cycle. Sci. Hortic. 2019, 244, 379–387. [Google Scholar] [CrossRef]
- Huan, C.; Han, S.A.; Jiang, L.; An, X.J.; Yu, M.L.; Xu, Y.; Ma, R.J.; Yu, Z.F. Postharvest hot air and hot water treatments affect the antioxidant system in peach fruit during refrigerated storage. Postharvest Boil. Technol. 2017, 126, 1–14. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.; Li, Z.; Zheng, Y.; Jin, P. Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis. Foods 2021, 10, 1662. https://doi.org/10.3390/foods10071662
Hou Y, Li Z, Zheng Y, Jin P. Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis. Foods. 2021; 10(7):1662. https://doi.org/10.3390/foods10071662
Chicago/Turabian StyleHou, Yuanyuan, Ziying Li, Yonghua Zheng, and Peng Jin. 2021. "Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis" Foods 10, no. 7: 1662. https://doi.org/10.3390/foods10071662
APA StyleHou, Y., Li, Z., Zheng, Y., & Jin, P. (2021). Effects of CaCl2 Treatment Alleviates Chilling Injury of Loquat Fruit (Eribotrya japonica) by Modulating ROS Homeostasis. Foods, 10(7), 1662. https://doi.org/10.3390/foods10071662