
Production of Indigo by Recombinant Escherichia coli with Expression of Monooxygenase, Tryptophanase, and Molecular Chaperone


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids and Culture Conditions
2.2. DNA Manipulation Techniques
2.3. Vectors Construction for Gene Expression in E. coli
2.4. Indigo Production by E. coli Fermentation
2.5. Enzymatic Activity Assay
2.6. Measurement of Indole, Indigo, and Tryptophan
3. Results
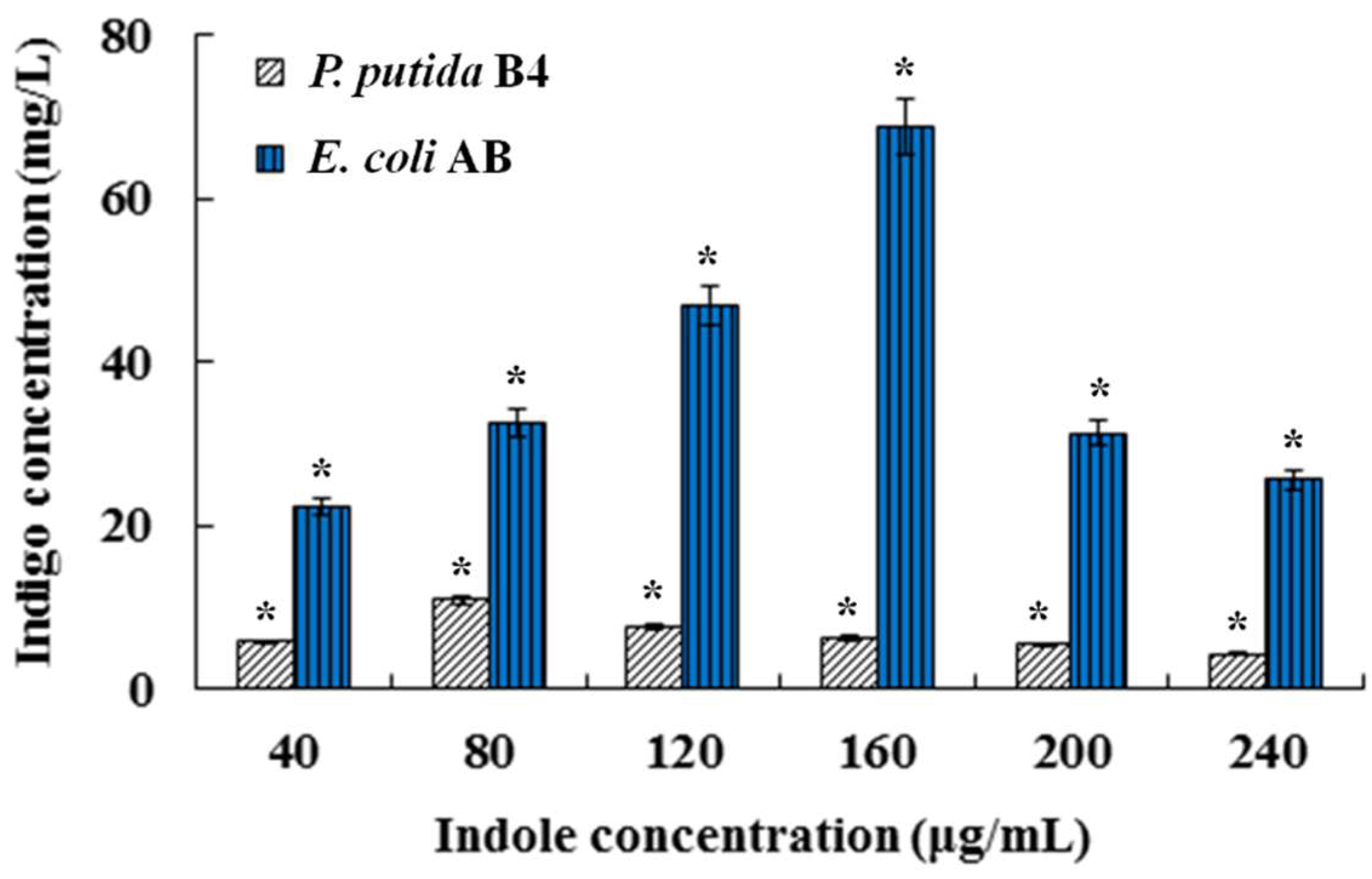
3.1. Expression of Styrene Monooxygenase Gene StyAB Generated Indigo Biosynthesis in E. coli
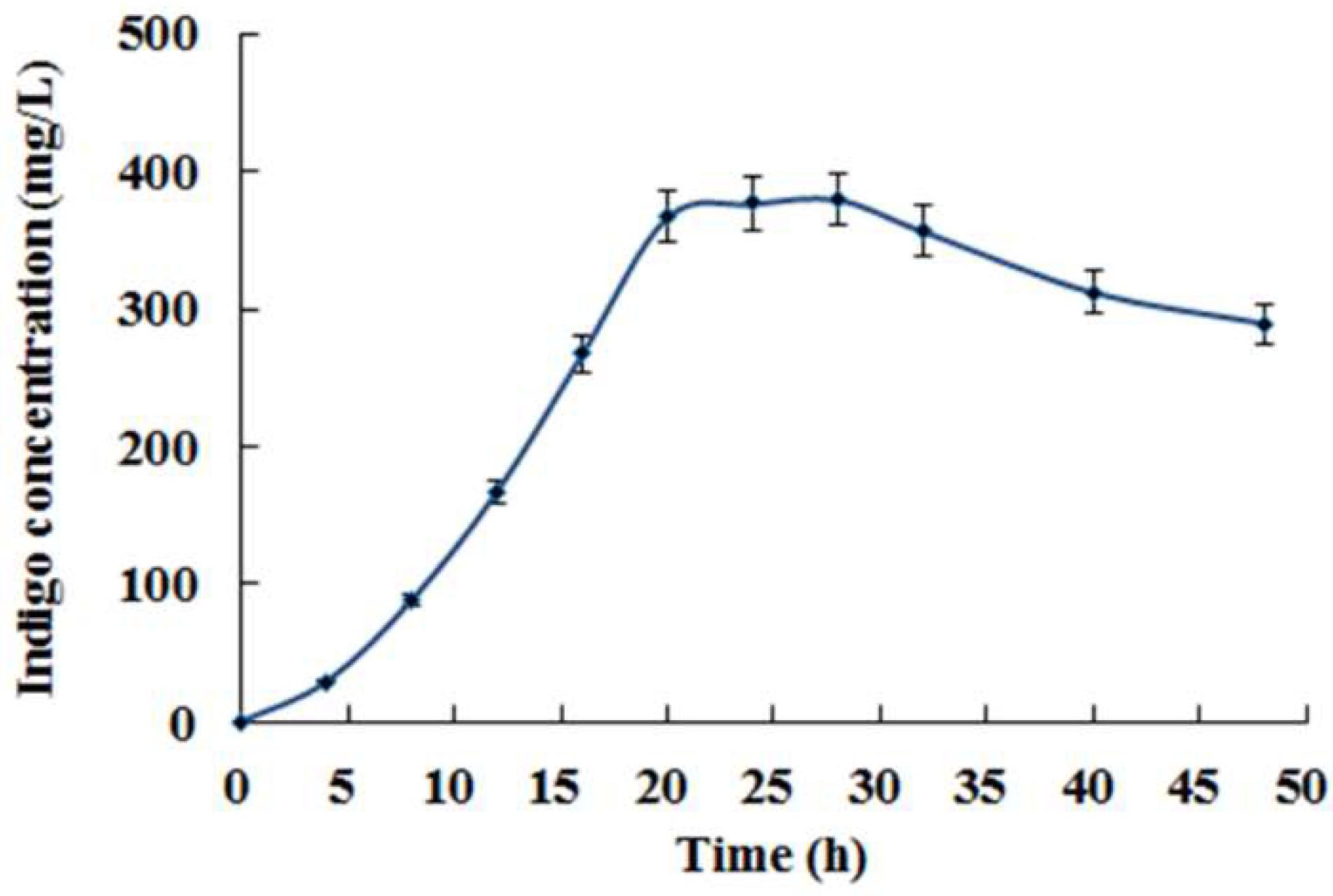
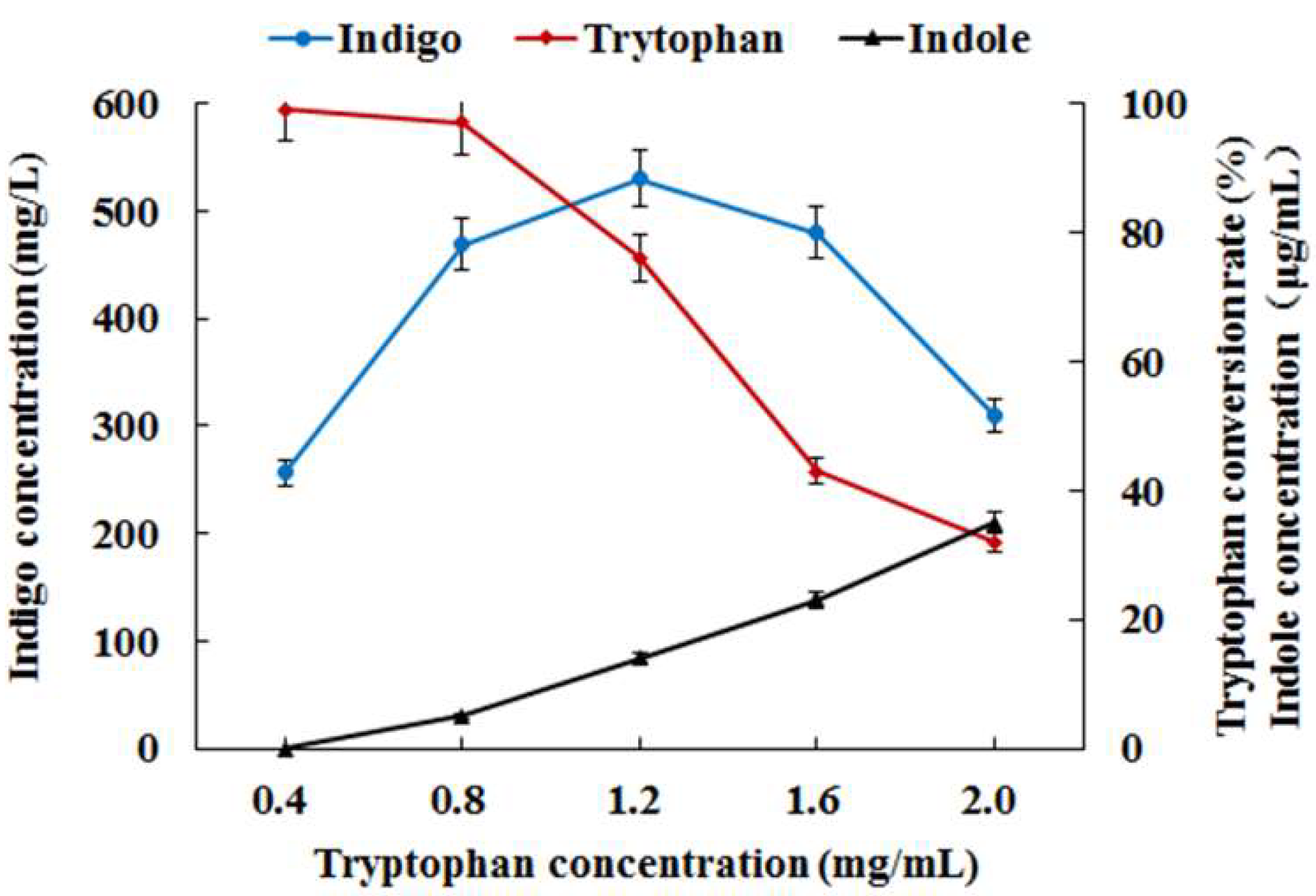
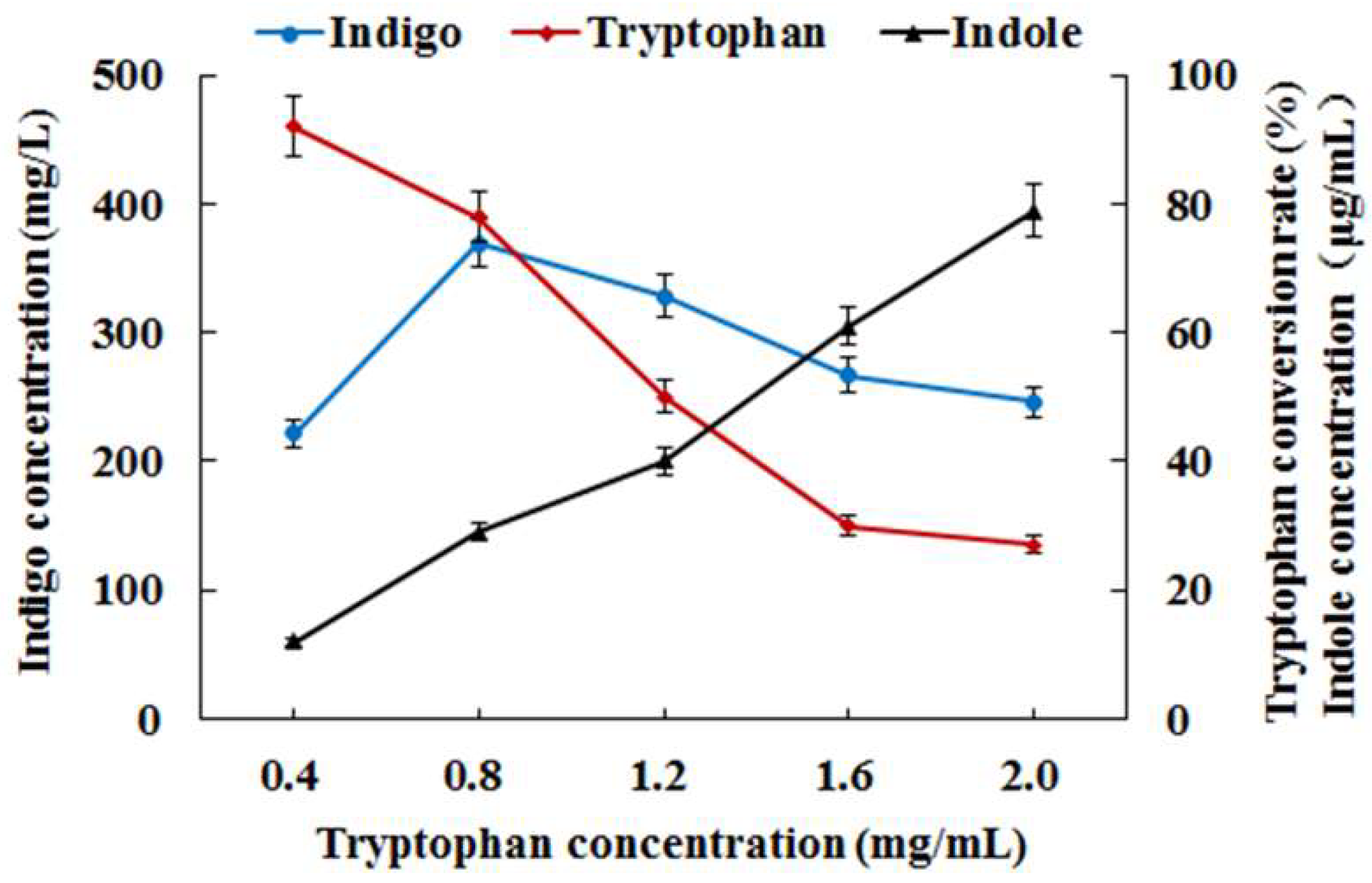
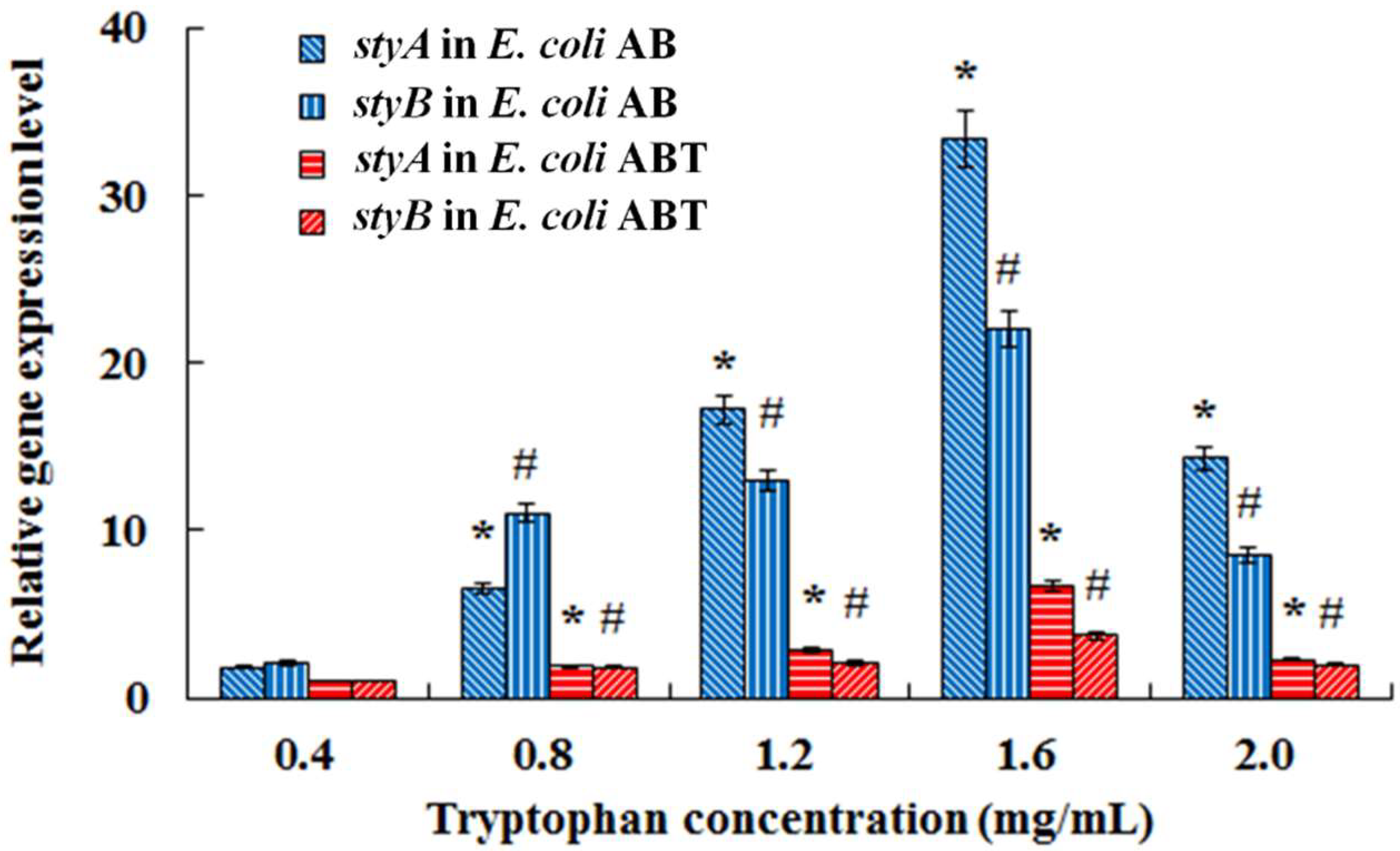
3.2. Co-Expression of Monooxygenase Gene StyAB and Tryptophanase Gene TnaA for Indigo Biosynthesis in E. coli
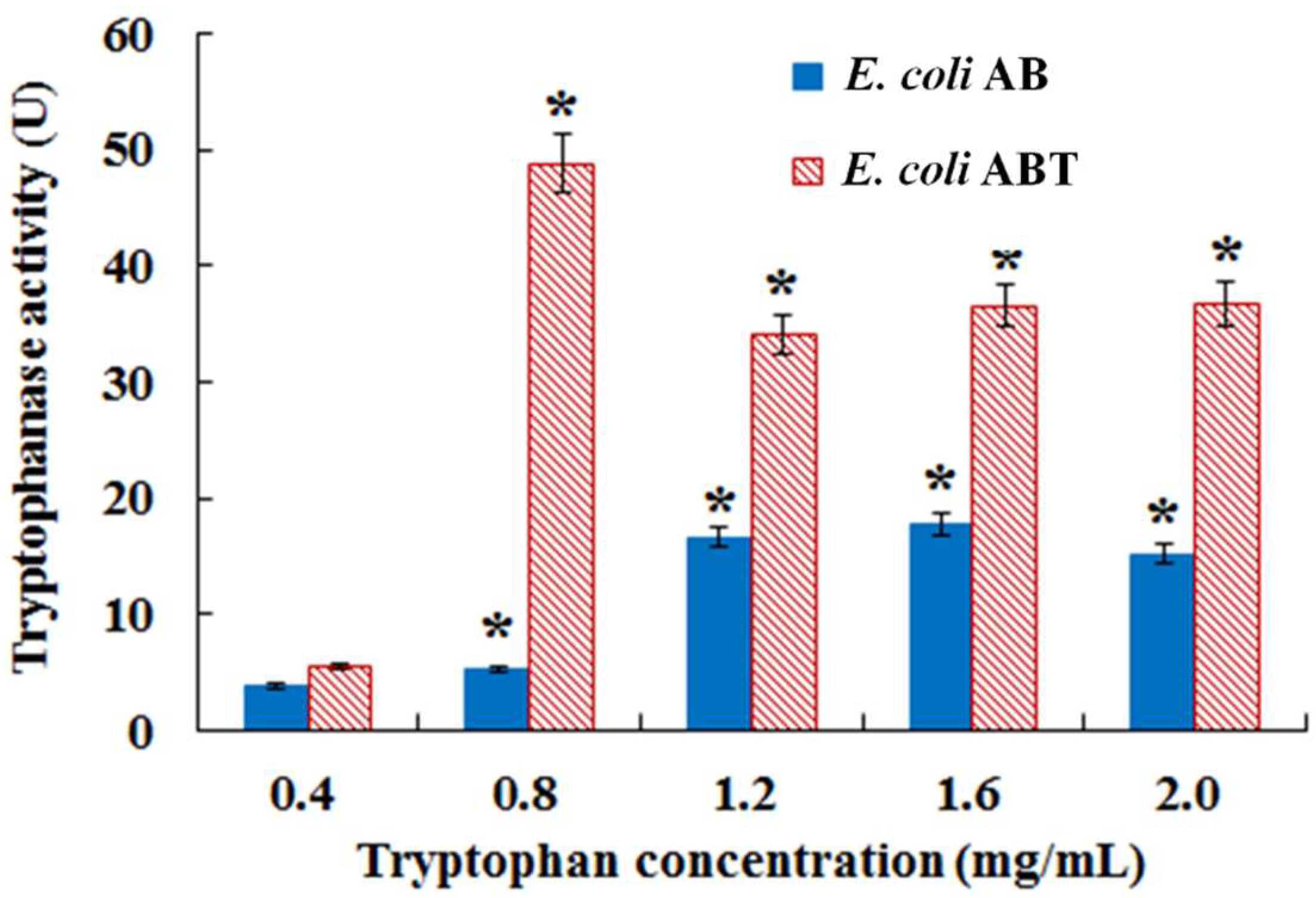
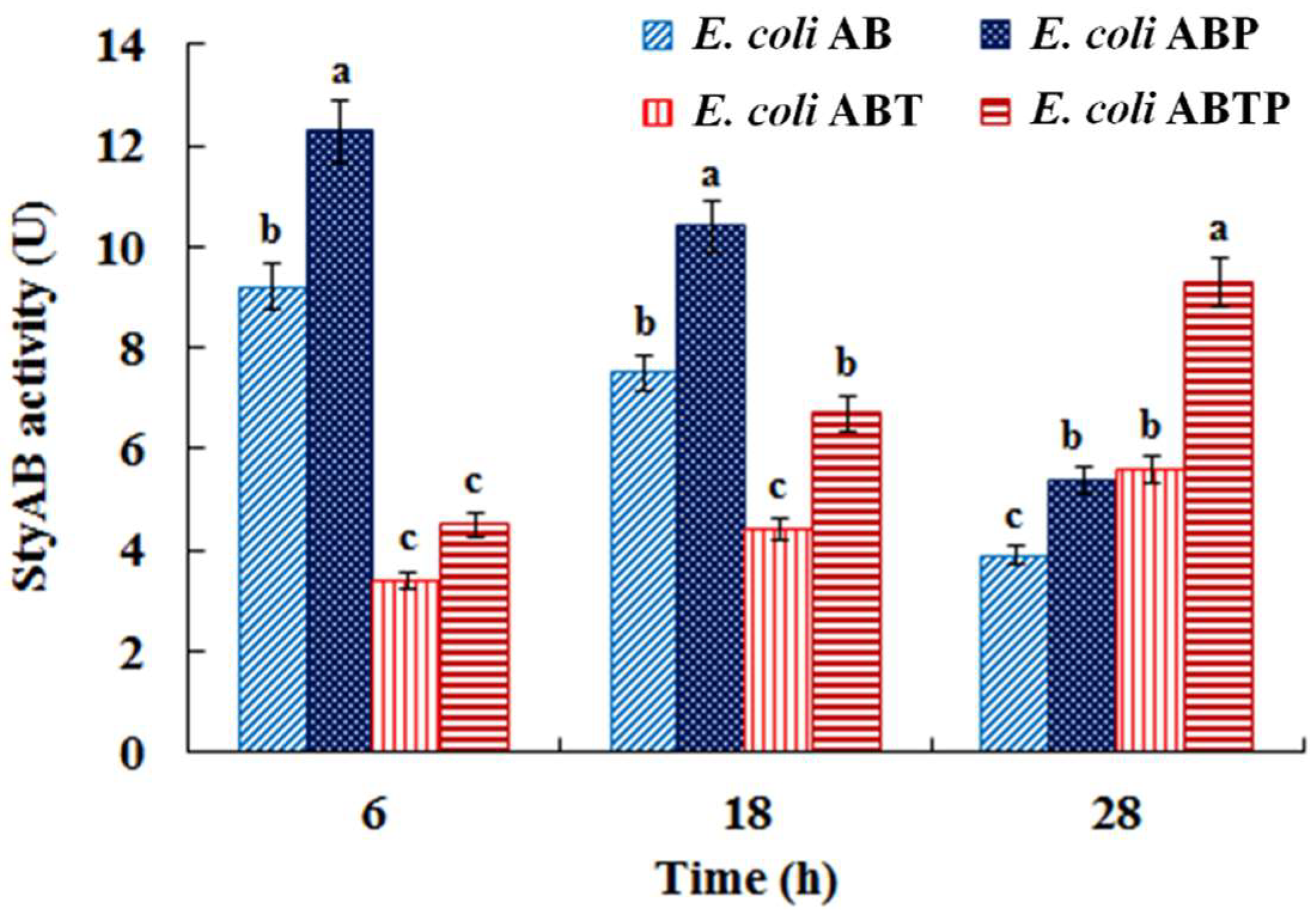
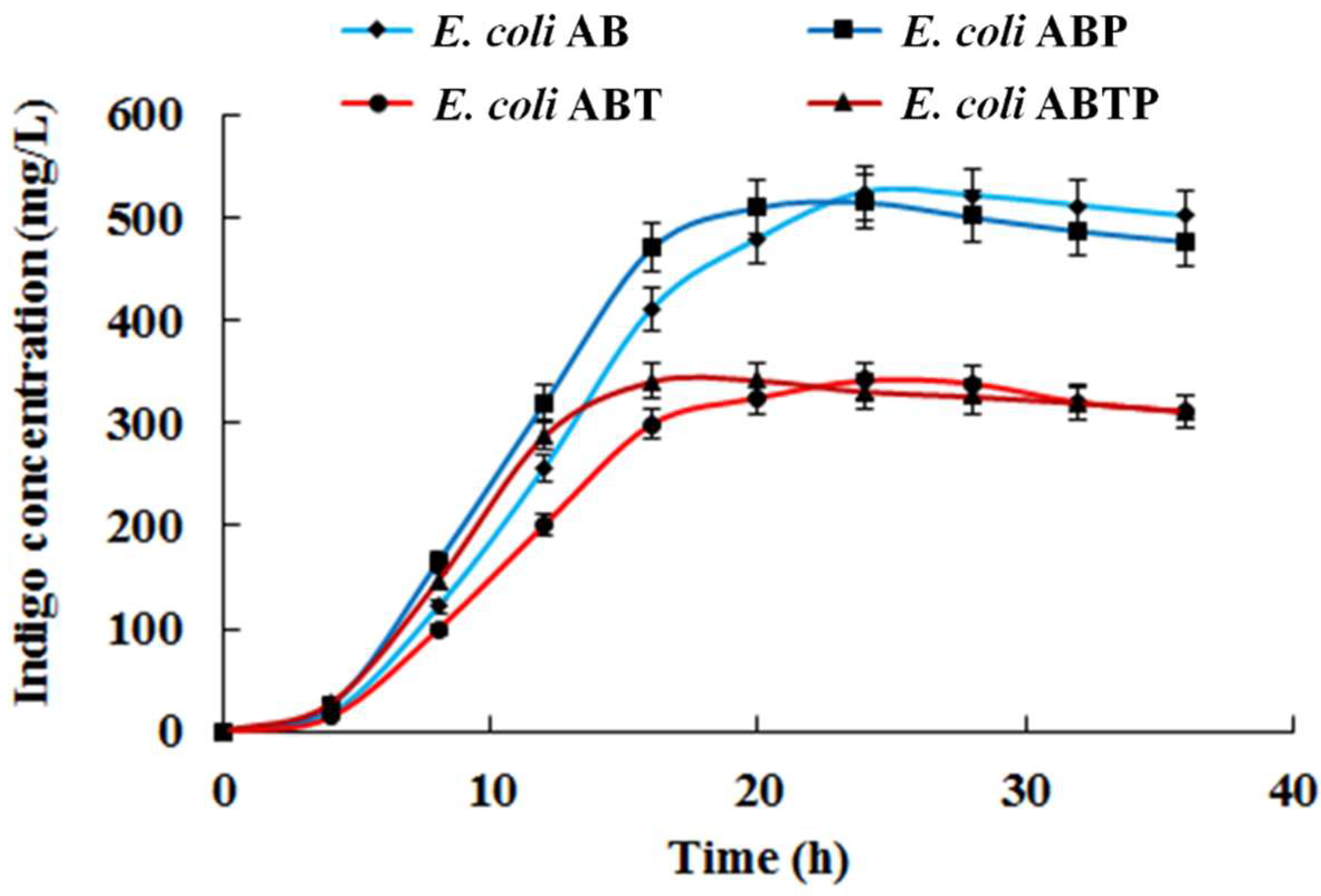
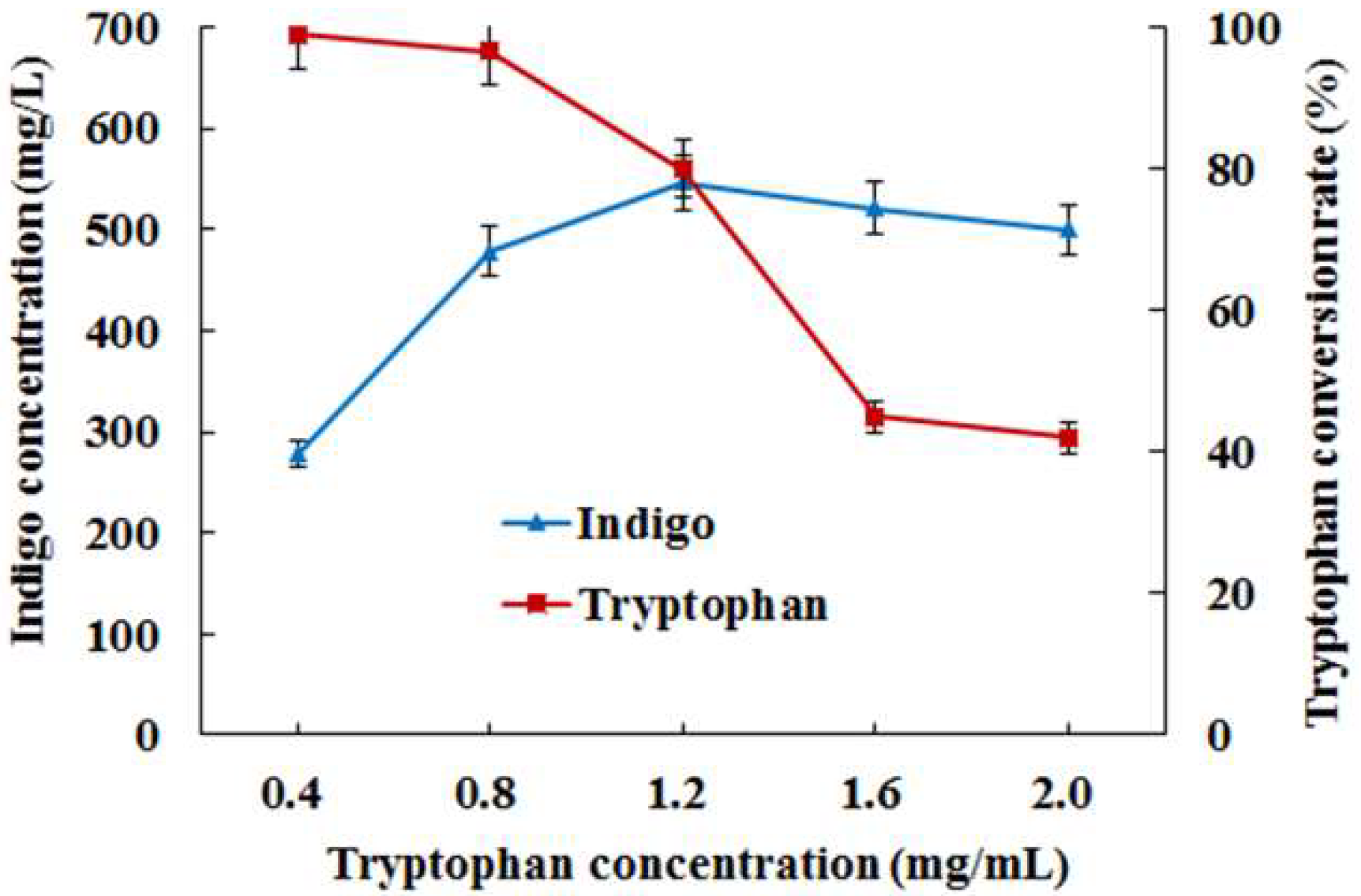
3.3. Introduction of Molecular Chaperone Enhanced the Activity of Monooxygenase StyAB and Indigo Biosynthesis in E. coli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Erkan, G.; Şengül, K.; Kaya, S. Dyeing of white and indigo dyed cotton fabrics with Mimosa tenuiflora extract. J. Saudi Chem. Soc. 2014, 18, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Han, G.H.; Bang, S.E.; Babu, B.K.; Chang, M.; Shin, H.-J.; Kim, S.W. Bio-indigo production in two different fermentation systems using recombinant Escherichia coli cells harboring a flavin-containing monooxygenase gene (fmo). Process Biochem. 2011, 46, 788–791. [Google Scholar] [CrossRef]
- Bechtold, T.; Turcanu, A.; Geissler, S.; Ganglberger, E. Process balance and product quality in the production of natural indigo from Polygonum tinctorium Ait. applying low-technology methods. Bioresour. Technol. 2002, 81, 171–177. [Google Scholar] [CrossRef]
- Pathak, H.; Madamwar, D. Biosynthesis of indigo dye by newly isolated naphthalene-degrading strain Pseudomonas sp. HOB1 and its application in dyeing cotton fabric. Appl. Biochem. Biotechnol. 2010, 160, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, B.; Samanta, S.K.; Jain, R.K. Indigo production by naphthalene-degrading bacteria. Lett. Appl. Microbiol. 2000, 31, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Doukyu, N.; Nakano, T.; Okuyama, Y.; Aono, R. Isolation of an Acinetobacter sp. ST-550 which produces a high level of indigo in a water-organic solvent two-phase system containing high levels of indole. Appl. Microbiol. Biotechnol. 2002, 58, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Gillam, E.M.; Aguinaldo, A.M.; Notley, L.M.; Kim, D.; Mundkowski, R.G.; Volkov, A.A.; Arnold, F.H.; Soucek, P.; DeVoss, J.J.; Guengerich, F.P. Formation of indigo by recombinant mammalian cytochrome P450. Biochem. Biophys. Res. Commun. 1999, 265, 469–472. [Google Scholar] [CrossRef]
- Mercadal, J.P.; Isaac, P.; Sineriz, F.; Ferrero, M.A. Indigo production by Pseudomonas sp. J26, a marine naphthalene-degrading strain. J. Basic Microbiol. 2010, 50, 290–293. [Google Scholar] [CrossRef]
- O’Connor, K.E.; Dobson, A.D.; Hartmans, S. Indigo formation by microorganisms expressing styrene monooxygenase activity. Appl. Environ. Microbiol. 1997, 63, 4287–4291. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.H.; Chen, H.P.; Huang, J.H.; Liu, T.T.; Lin, T.K.; Wang, S.J.; Tseng, C.H.; Shu, H.Y. Identification and characterization of an indigo-producing oxygenase involved in indole 3-acetic acid utilization by Acinetobacter baumannii. Antonie Van Leeuwenhoek 2012, 101, 881–890. [Google Scholar] [CrossRef]
- Qu, Y.; Ma, Q.; Zhang, X.; Zhou, H.; Li, X.; Zhou, J. Optimization of indigo production by a newly isolated Pseudomonas sp. QM. J. Basic Microbiol. 2012, 52, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Mermod, N.; Harayama, S.; Timmis, K.N. New route to bacterial production of indigo. Nat. Biotechnol. 1986, 4, 321–324. [Google Scholar] [CrossRef]
- Furuya, T.; Takahashi, S.; Ishii, Y.; Kino, K.; Kirimura, K. Cloning of a gene encoding flavin reductase coupling with dibenzothiophene monooxygenase through coexpression screening using indigo production as selective indication. Biochem. Biophys. Res. Commun. 2004, 313, 570–575. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Shi, S.; Zhou, H.; Ma, Q.; Li, X.; Zhang, X.; Zhou, J. Characterization of a novel phenol hydroxylase in indoles biotransformation from a strain Arthrobacter sp. W1. PLoS ONE 2012, 7, e44313. [Google Scholar] [CrossRef]
- Cheng, L.; Yin, S.; Chen, M.; Sun, B.; Hao, S.; Wang, C. Enhancing indigo production by over-expression of the styrene monooxygenase in Pseudomonas putida. Curr. Microbiol. 2016, 73, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Green, M.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.D.; O’Connor, K.E.; Dobson, A.D. Biochemistry, genetics and physiology of microbial styrene degradation. FEMS Microbiol. Rev. 2002, 26, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Garbe, T.R.; Kobayashi, M.; Yukawa, H. Indole-inducible proteins in bacteria suggest membrane and oxidant toxicity. Arch. Microbiol. 2000, 173, 78–82. [Google Scholar] [CrossRef]
- Wang, D.; Ding, X.; Rather, P.N. Indole can act as an extracellular signal in Escherichia coli. J. Bacteriol. 2001, 183, 4210–4216. [Google Scholar] [CrossRef] [Green Version]
- Snell, E.E. Tryptophanase: Structure, catalytic activities, and mechanism of action. Adv. Enzymol. Relat. Areas Mol. Biol. 1975, 42, 287–333. [Google Scholar] [CrossRef]
- Rydenfelt, M.; Garcia, H.G.; Cox, R.S.; Phillips, R. The influence of promoter architectures and regulatory motifs on gene expression in Escherichia coli. PLoS ONE 2014, 9, e114347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royo, J.L.; Moreno-Ruiz, E.; Cebolla, A.; Santero, E. Stable long-term indigo production by overexpression of dioxygenase genes using a chromosomal integrated cascade expression circuit. J. Biotechnol. 2005, 116, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, K.; Kanemori, M.; Kitagawa, M.; Yanagi, H.; Yura, T. Chaperone coexpression plasmids: Differential and synergistic roles of DnaK-DnaJ-GrpE and GroEL-GroES in assisting folding of an allergen of Japanese cedar pollen, Cryj2, in Escherichia coli. Appl. Environ. Microbiol. 1998, 64, 1694–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, K.; Hofstetter, K.; Röthlisberger, M.; Witholt, B.; Schmid, A. Biochemical characterization of StyAB from Pseudomonas sp. strain VLB120 as a two-component flavin-diffusible monooxygenase. J. Bacteriol. 2004, 186, 5292–5302. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Relevant Features | Source |
|---|---|---|
| Plasmids | ||
| pBK-CMV | Vector for gene expression in E. coli, KanR | Laboratory collection |
| pBK-AB | styAB gene cloned in pBK-CMV, KanR | This work |
| pBK-ABT | TnaA gene cloned in pBK-AB, KanR | This work |
| pGro7 | Chaperone plasmid harboring groES-groEL, CmR | Takara, Beijing, China |
| Strains | ||
| E. coli DH5α | Host for gene expression | TIANGEN, Beijing, China |
| E. coli AB | E. coli DH5α harboring pBK-AB | This work |
| E. coli ABT | E. coli DH5α harboring pBK-ABT | This work |
| E. coli ABTP | E. coli DH5α harboring pBK-ABT and pGro7 | This work |
| P. putida B4 | Donor of styAB gene | Laboratory collection |
| Gene | Primer | Sequence (5′–3′) |
|---|---|---|
| q RT PCR | ||
| styA | styA-F | GGCGAGCTGATTGAGATTC |
| styA-R | TTTTGCCGTTATTGAGGGT | |
| styB | styB-F | AAAAGATGTGGTGGTGGAT |
| styB-R | TGCTGAAGAATGCCGATAA | |
| 16S rRNA | 16S-F | CCACCTGGACTGATACT |
| 16S-R | GCACCTGTCTCAATGTT | |
| PCR | ||
| styAB | styAB-F | AACTGCAGATGAAAAAGCGTATCGGTATTG |
| styAB-R | CCCAAGCTTTCAATTCAGTGGCAACGGGTT | |
| TnaA | TnaA-F | CCCAAGCTTATGGAAAACTTTAAACATCTCCC |
| TnaA-R | CTAGTCTAGATTAAACTTCTTTAAGTTTTGCGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, L.; Yue, J.; Zhu, Y.; Yin, S. Production of Indigo by Recombinant Escherichia coli with Expression of Monooxygenase, Tryptophanase, and Molecular Chaperone. Foods 2022, 11, 2117. https://doi.org/10.3390/foods11142117
Du L, Yue J, Zhu Y, Yin S. Production of Indigo by Recombinant Escherichia coli with Expression of Monooxygenase, Tryptophanase, and Molecular Chaperone. Foods. 2022; 11(14):2117. https://doi.org/10.3390/foods11142117
Chicago/Turabian StyleDu, Lingyan, Jianming Yue, Yiying Zhu, and Sheng Yin. 2022. "Production of Indigo by Recombinant Escherichia coli with Expression of Monooxygenase, Tryptophanase, and Molecular Chaperone" Foods 11, no. 14: 2117. https://doi.org/10.3390/foods11142117
APA StyleDu, L., Yue, J., Zhu, Y., & Yin, S. (2022). Production of Indigo by Recombinant Escherichia coli with Expression of Monooxygenase, Tryptophanase, and Molecular Chaperone. Foods, 11(14), 2117. https://doi.org/10.3390/foods11142117