
A Novel Fermented Rapeseed Meal, Inoculated with Selected Protease-Assisting Screened B. subtilis YY-4 and L. plantarum 6026, Showed High Availability and Strong Antioxidant and Immunomodulation Potential Capacity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Method
2.2.1. Preparation of the FRSMP
2.2.2. Nutritional Compositions and Microstructure of the RSM before and after Fermentation
2.2.3. Digestibility of the DM and CP of the RSM before and after Fermentation
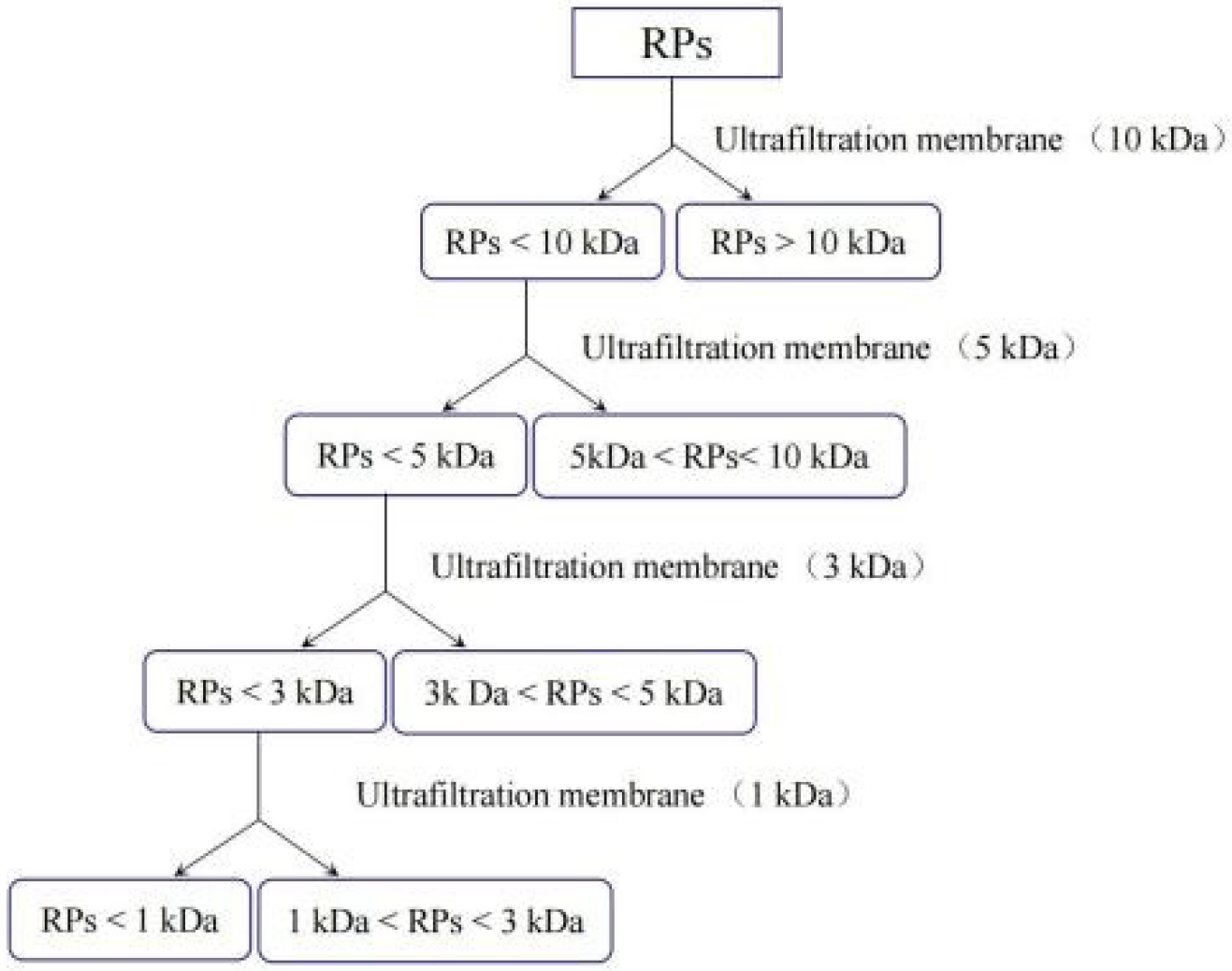
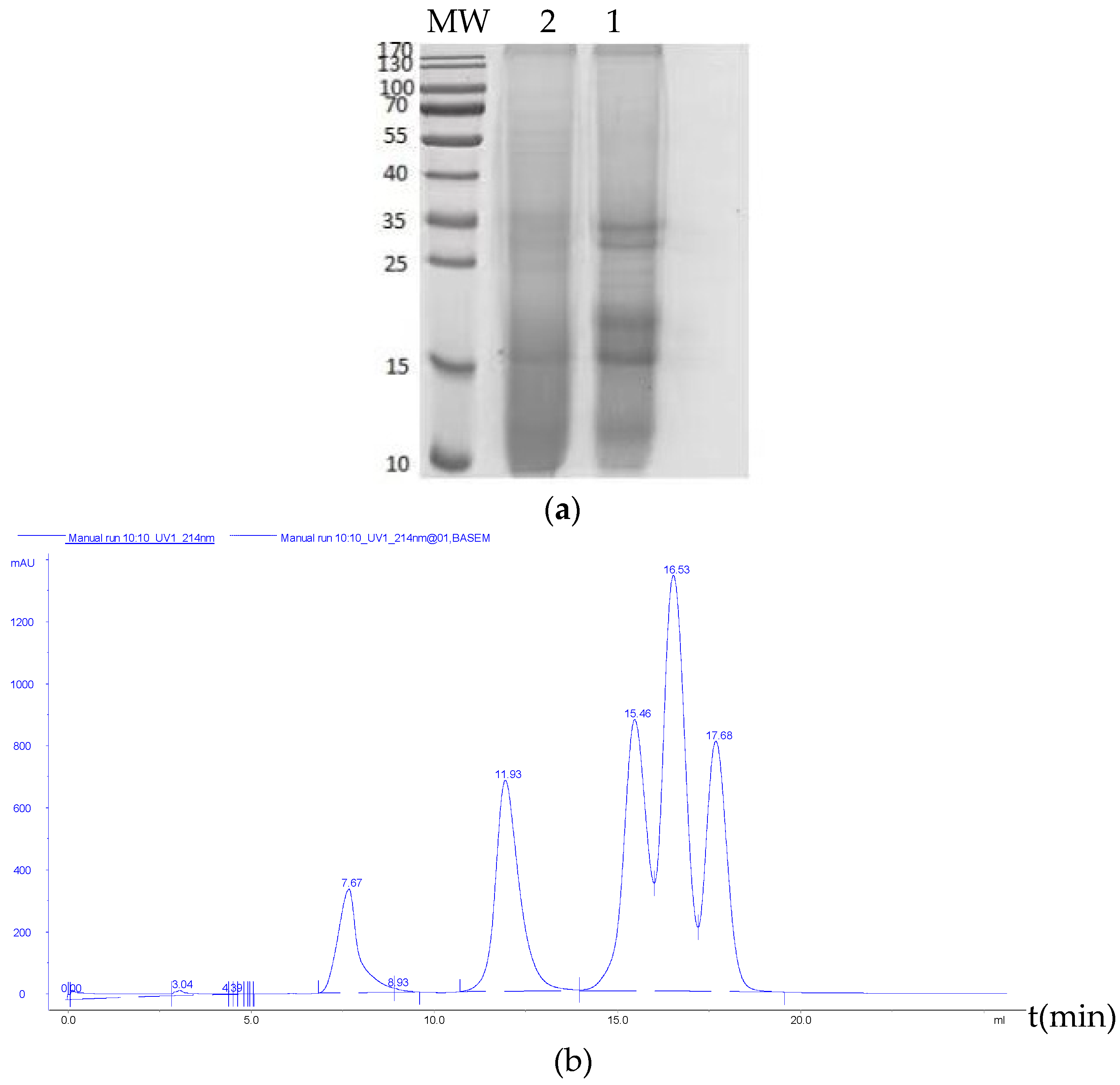
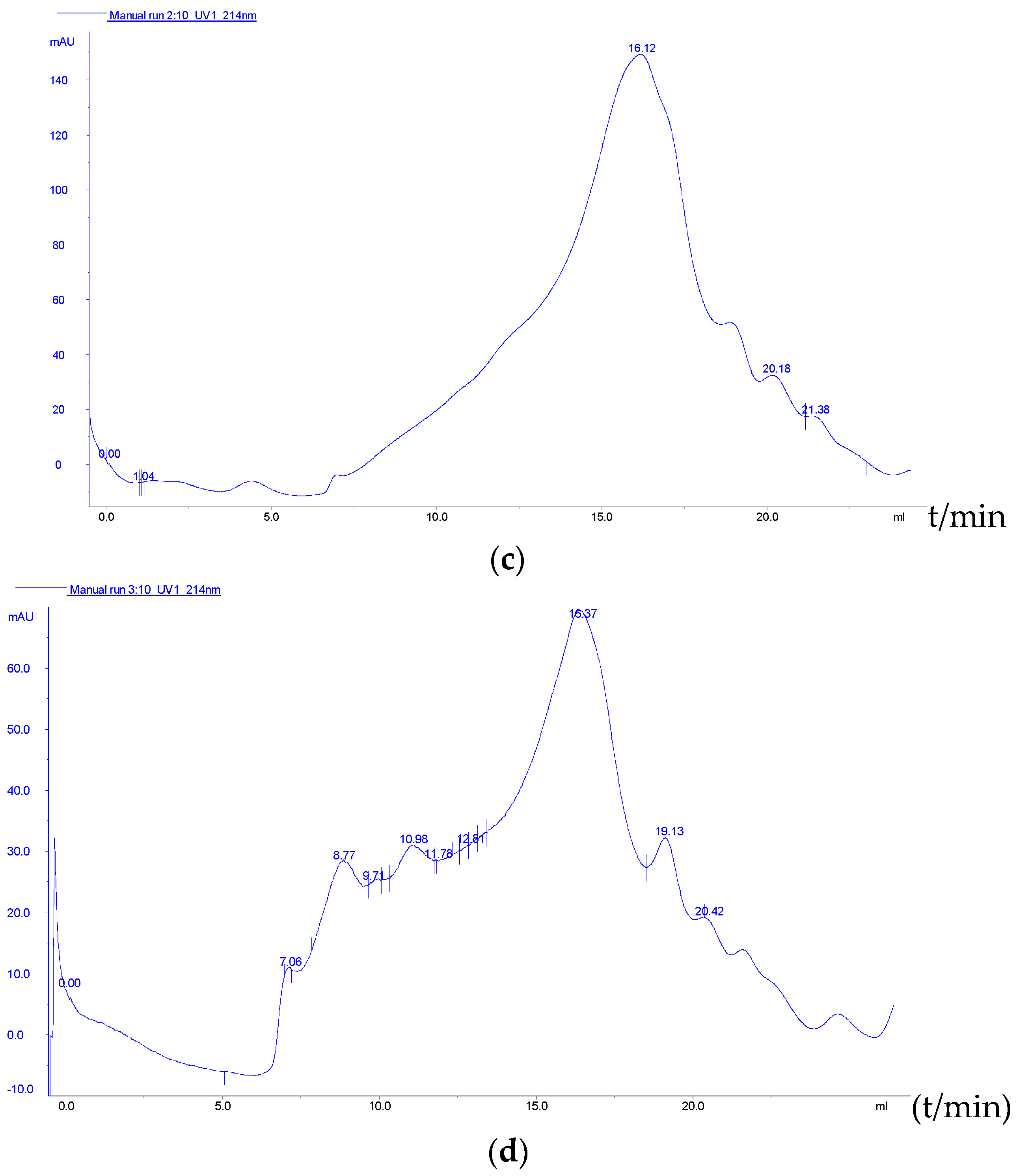
2.2.4. Molecular Weight Distribution and Free Amino Acid Profile of the RSM before and after Fermentation
2.2.5. Evaluation of the Antioxidant Activity of the RSM before and after Fermentation In Vitro
2.2.6. The Availability and Bioactivity In Vivo of the RSM before and after Fermentation
2.2.7. Statistical Analysis
3. Results and Analyses
3.1. Establishment of Novel Fermentation System
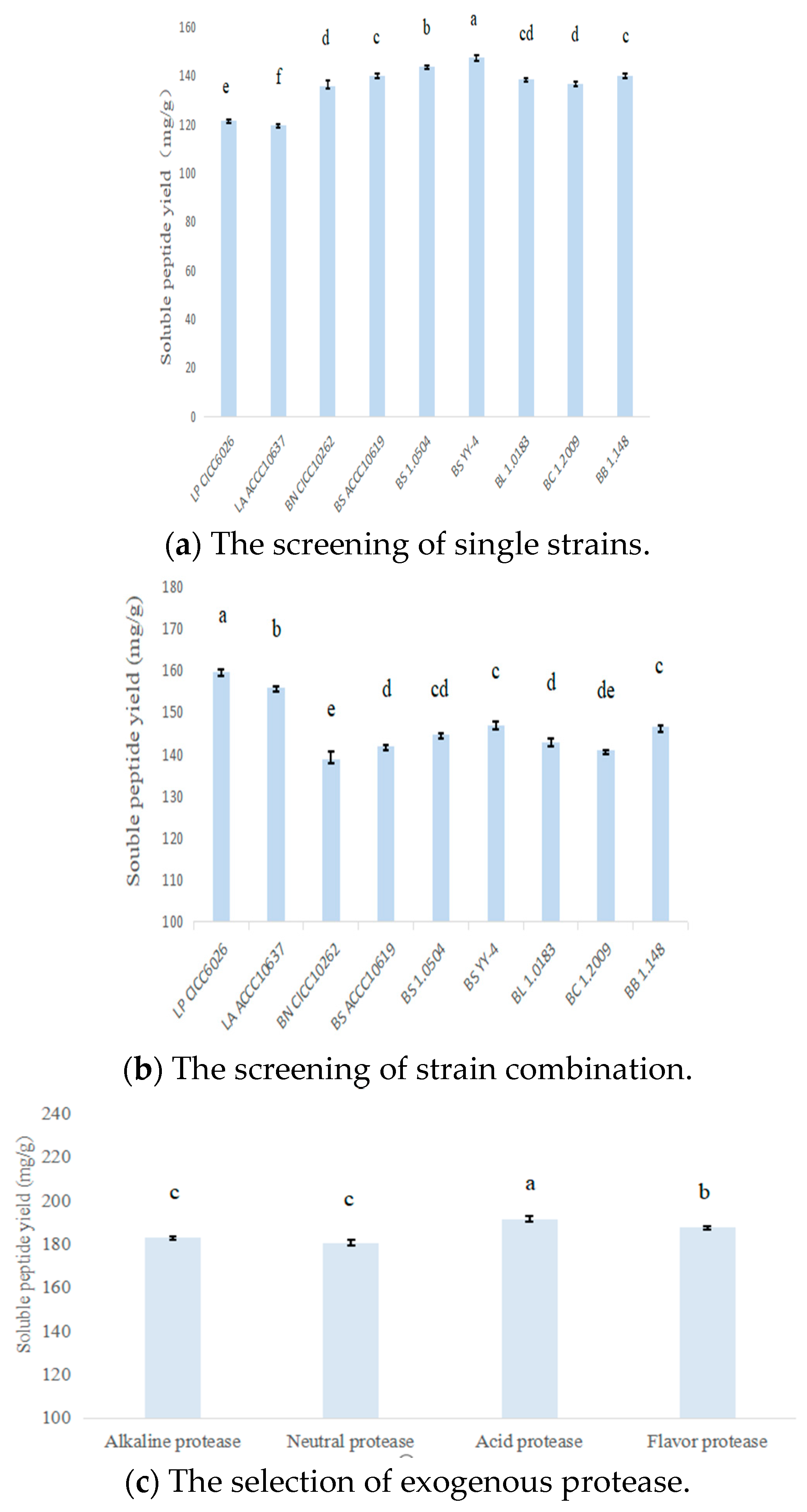
3.1.1. Screening of Strains and Exogenous Protease
3.1.2. Optimization of Fermentation Condition
3.2. Surface Morphology and Microstructure of the RSM before and after Fermentation
3.3. Chemical Compositions of the RSM before and after Fermentation
3.4. Molecular Weight Distribution and Free Amino Acid Profile of the RSM before and after Fermentation
3.5. Availability In Vitro and In Vivo of the RSM before and after Fermentation
3.6. Antioxidant Activity In Vitro of the RSM before and after Fermentation
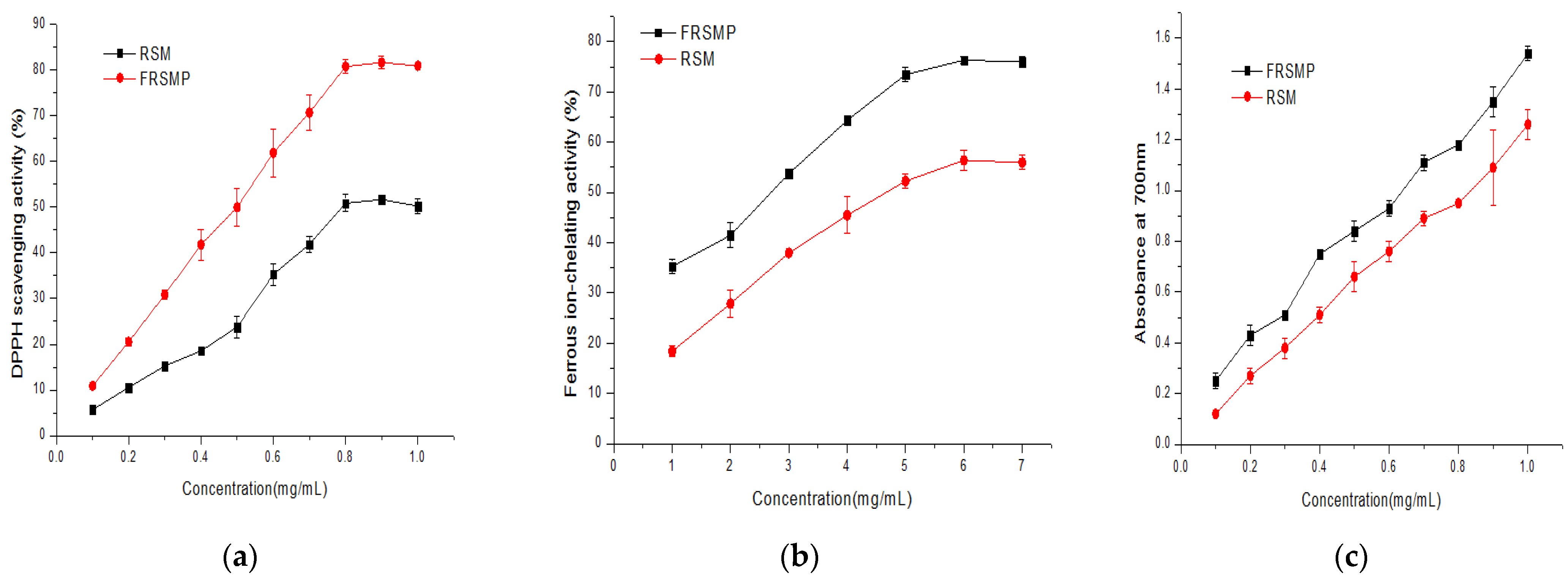
3.6.1. DPPH Scavenging Activity
3.6.2. Ferrous-Ion-Chelating Activity
3.6.3. Reducing Power
3.7. Immunomodulation Activity In Vivo of the RSM before and after Fermentation
4. Discussion
4.1. Discussion on the Novel Fermentation System
4.1.1. Discussion on the Novel Fermentation Method
4.1.2. Discussion on the Optimization of Fermentation Conditions
4.2. Discussion on the Changes of the Nutritional Values of the RSM after Fermentation
4.3. Discussion on the Change of the Molecular Weight Distribution and Free Amino Acid Composition of the RSM after Fermentation
4.4. Discussion on the Availability of the RSM after Fermentation
4.5. Discussion on the Antioxidant In Vitro of the RSM after Fermentation
4.6. Discussion on the Immunomodulation Activity In Vitro of the RSM after Fermentation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Citeau, M.; Regis, J.; Carr’e, P.; Fine, F. Value of hydroalcoholic treatment of rapeseed for oil extraction and protein enrichment. OCL 2016, 26, 1. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.S.; Dai, C.H.; Tang, Y.X.; Xing, Z.; Mintah, B.K.; Dabbour, M.; Ding, Q.Z.; He, R.H.; Ma, H.L. Thermophilic solid-state fermentation of rapeseed meal and analysis of microbial community diversity. LWT Food Sci. Technol. 2019, 116, 108520. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Yan, Z.B.; An, C.M.; Wang, D.; Liu, X.; Zhang, J.F.; Yang, W.L.; Deng, B.C.; Xie, J.Q. Rapeseed protein-derived antioxidant peptide RAP alleviates renal fibrosis through MAPK/NFκB signaling pathways in diabetic nephropathy. Drug Des. Dev. Ther. 2018, 12, 1255–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, I.M.; Carvalho, M.G.V.S.; Rocha, J.M.S. Increase of protein extraction yield from rapeseed meal through a pretreatment with phytase. J. Sci. Food Agr. 2017, 97, 2641–2646. [Google Scholar] [CrossRef]
- Zhang, S.B.; Wang, Z.; Xu, S.Y.; Gao, X.F. Purification and characterization of a radical scavenging peptide from rapeseed protein hydrolysated. J. Am. Oil Chem. Soc. 2009, 86, 959–966. [Google Scholar] [CrossRef]
- Xue, Z.; Yu, W.; Liu, Z.; Wu, M.; Kou, X.; Wang, J. Preparation and antioxidative properties of a rapeseed (Brassica napus) protein hydrolysate and three peptide fractions. J. Agr. Food Chem. 2009, 57, 5287–5293. [Google Scholar] [CrossRef]
- Jia, A.W.Q.; Rodriguez-Alonso, E.; Bianeis, M.; Keppler, J.K.; Goots, A.J. Assessing functional properties of rapeseed protein concentrate versus isolate for food applications. Innov. Food Sci. Emerg. Technol. 2021, 68, 102636. [Google Scholar] [CrossRef]
- Simitzis, P. Agro-industrial by-products and their bioactive compounds-anally against Oxidative stress and skin aging. Cosmetics 2018, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Terefe, N.S.; Augustin, M.A. Fermentation for tailoring the technological and health related functionality of food products. Crit. Rev. Food Sci. 2020, 60, 2887–2913. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biot. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [Green Version]
- Adeola, M.A.; Christophe, L.B.; Rodney, J.M.; Samson, O.A.; John, M.; Rong, H.; Abraham, G.; Rotimi, E.A. Antioxidant properties of Australian canola meal protein hydrolysates. Food Chem. 2014, 146, 500–506. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Hsiao, F.S.H.; Wen, C.M.; Wu, C.Y.; Dybus, A.; Yu, Y.H. Mixed fermentation of soybean meal by protease and probiotics and its effects on the growth performance and immune response in broilers. J. Appl. Anim. Res. 2019, 47, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.W.; Xia, P.B.; Zhang, L.L.; Hu, Y.; Xie, Q.H.; Xiang, H.Y. A novel fermented soybean, inoculated with selected Bacillus, Lactobacillus and Hansenula strains, showed strong antioxidant and anti-fatigue potential activity. Food Chem. 2020, 333, 127527. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Tech. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Wei, F.H.; Liu, X.L.; Yi, C.X.; Zhang, Y.G. Improvement of the nutritional value, sensory properties and bioavailability of rapeseed meal fermented with mixed microorganisms. LWT-Food Sci. Technol. 2019, 112, 108238. [Google Scholar] [CrossRef]
- Rong, H.; Ju, X.R.; Yuan, J.; Wang, L.F.T.; Girgih, A.; Aluko, R.E. Antioxidant activities of rapeseed peptides produced by solid state fermentation. Food Res. Int. 2012, 49, 432–438. [Google Scholar] [CrossRef]
- Li, S.L.; Jin, Z.Y.; Hua, D.J.; Yang, W.W.; Yan, Y.Y.; Nie, X.B.; Lin, J.; Zhang, Q.Y.; Gai, D.; Jia, Y.X.; et al. Effect of solid-state fermentation with Lactobacillus casei on the nutritional value, isoflavones, phenolic acids and antioxidant activity of whole soybean flour. LWT Food Sci. Tech. 2020, 125, 109264. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, X.; Xu, H.J.; Sun, Y.K.; Zhang, Y.G.; Wang, Y. Improvement of the nutritional, antioxidant and bioavailability properties of corn gluten-wheat bran mixture fermented with lactic acid bacteria and acid protease. LWT-Food Sci. Tech. 2021, 144, 111161. [Google Scholar] [CrossRef]
- Rezazadeh, F.; Kowsar, R.; Rafiee, H.; Riasi, A. Fermentation of soybean meal improves growth performance and immune response of abruptly weaned Holstein calves during cold weather. Anim. Feed Sci. Tech. 2019, 254, 114206. [Google Scholar] [CrossRef]
- Albaser, A.; Kazana, E.; Bennett, M.; Cebeci, F.; Luang-In, V.; Spanu, P.D.; Rossiter, J.T. Discovery of a Bacterial Glycoside Hydrolase Family 3 (GH3) β-Glucosidase with Myrosinase Activity from a Citrobacter Strain Isolated from Soil. J. Agri. Food Chem. 2016, 64, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar] [CrossRef]
- Dai, Y.; Tan, J.N.; Yang, R.J.; Yuan, Q.; Che, L.M.G.; Zhu, S.H. Optimization on the determination of glucosinolates in rapeseed with Anthrone colorimetry. J. Tradit. Chin. Med. 2014, 4, 60–61. [Google Scholar] [CrossRef]
- Wang, Y.C.; Xu, K.K.; Lu, F.; Wang, Y.N.; Ouyang, N.N.; Ma, H.L. Increasing peptide yield of soybean meal solid-state fermentation of ultrasound-treated Bacillus amyloliquefaciens. Innov. Food Sci. Emerg. 2021, 72, 102704. [Google Scholar] [CrossRef]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 38, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Laguna, O.; Barakat, A.; Alhamada, H.; Durand, E.; Baréa, B.; Fine, F.; Villeneuve, P.; Citeau, M.; Dauguet, S.; Lecomte, J. Production of proteins and phenolic compounds enriched fractions from rapeseed and sunflower meals by dry fractionation processes. Ind. Crops Prod. 2018, 118, 160–172. [Google Scholar] [CrossRef]
- Wheeler, E.L.; Ferrel, R.E. A method for phytic acid determination in wheat and wheat fractions. Cereal Chem. 1971, 48, 312–320. [Google Scholar]
- Wilfart, A.; Jaguelin-Peyraud, Y.; Simmins, H.; Noblet, J.; Van Milgen, J.; Montagne, L. Kinetics of enzymatic digestion of feeds as estimated by a stepwise in vitro method. Anim. Feed Sci. Technol. 2008, 141, 171–183. [Google Scholar] [CrossRef]
- Zheng, W.K.; Zhao, F.; Zhang, H.F. Effect of Dietary Type and Storage Condition of Intestinal juice on and character of Jejunal Juice in Ducks. Chin. J. Anim. Nutr. 2009, 21, 652–658. [Google Scholar] [CrossRef]
- Sun, H.; Tang, J.W.; Yao, X.H.; Wu, Y.F.; Wang, X.; Feng, J. Improvement of the nutritional quality of cottonseed meal by Bacillus subtilis and the addition of papain. Int. J. Agr. Biol. Eng. 2012, 14, 563–568. [Google Scholar] [CrossRef]
- Lin, F.; Chen, L.; Liang, R.; Zhang, Z.; Wang, J.; Cai, M.; Li, Y. Pilot-scale production of low molecular weight peptides from corn wet milling byproducts and the antihypertensive effects in vivo and in vitro. Food Chem. 2011, 124, 801–807. [Google Scholar] [CrossRef]
- Jiang, X.; Cui, Z.Q.; Wang, L.H.; Xu, H.J.; Zhang, Y.G. Production of bioactive peptides from corn gluten meal by solid-state fermentation with Bacillus subtilis MTCC5480 and evaluation of its antioxidant capacity in vivo. LWT-Food Sci. Technol. 2020, 8131, 109767. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, L.X.; Rui, X.; Li, W.; Chen, X.H.; Jiang, M.; Dong, M.S. Enhancement of the antioxidant capacity of soy whey by fermentation with Lactobacillus plantarum B1-6. J. Func. Foods. 2015, 12, 33–44. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT® 9.1.3 User’s Guide, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2009. [Google Scholar]
- Bao, J.; Zhang, X.; Zheng, J.H.; Ren, D.F.; Lu, J. Mixed fermentation of Spirulina platensis with Lactobacillus plantarum and Bacillus subtilis by random-centroid optimization. Food Chem. 2018, 264, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.J.; Yang, Z.B.; Yang, W.R.; Li, H.R.; Zhang, C.Y.; Jiang, S.Z.; Li, X.M. Conventional solid fermentation alters mycotoxin contents and microbial diversity analyzed by high-throughput sequencing of a Fusarium mycotoxin-contaminated diet. Can. J. Anim. Sci. 2018, 98, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Małgorzata, K.P.; Borowczyk, P.; Zaworska, A.; Nowak, W.; Frankiewicz, A.; Gulewicz, P. The Effect of Dry Yeast Fermentation on Chemical Composition and Protein Characteristics of Blue Lupin Seeds. Food Technol. Biotechnol. 2016, 54, 360–366. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, H.; Han, B.; Li, H.Y.; Liu, X.L. Improvement of nutritional value, molecular weight patterns (soluble peptides), free amino acid patterns, total phenolics and antioxidant activity of fermented extrusion pretreatment rapeseed meal with Bacillus subtilis YY-1 and Saccharomyces cerevisiae YY-2. LWT-Food Sci. Technol. 2022, 160, 113280. [Google Scholar] [CrossRef]
- Idris, A.S.O.; Pandey, A.; Rao, S.S.; Sukumaran, R.K. Cellulase production through solid-state tray fermentation, and its use for bioethanol from sorghum stover. Bioresour. Technol. 2017, 242, 265–271. [Google Scholar] [CrossRef]
- Xing, Z.; Hou, X.S.; Tang, Y.X.; He, R.H.; Mintah, B.K.; Dabbour, M.H. Monitoring of polypeptide content in the solid-state fermentation process of rapeseed meal using NIRS and chemometrics. J. Food Process Eng. 2018, 41, e12853. [Google Scholar] [CrossRef]
- Couto, S.R.; Sanroman, M.A. Application of solid-state fermentation to food Industry—A review. J. Food Eng. 2006, 76, 291–302. [Google Scholar] [CrossRef]
- Asensio-Grau, A.; Calvo-Lerma, J.; Heredia, A.; Andrés, A. In vitro digestion of salmon: Influence of processing and intestinal conditions on macronutrients digestibility. Food Chem. 2021, 342, 128387. [Google Scholar] [CrossRef]
- Olukomaiya, O.; Adiamo, O.Q.; Fernando, W.C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Effect of solid-state fermentation on proximate composition, anti-nutritional factor, microbiological and functional properties of lupin flour. Food Chem. 2020, 315. [Google Scholar] [CrossRef]
- Polanowska, K.; Grygier, A.; Kuligowski, M.; Rudzi´nska, M.; Nowak, N. Effect of Tempe fermentation by three different strains of Rhizopus oligosporus on nutritional characteristics of faba beans. LWT-Food Sci. Technol. 2020, 122, 109024. [Google Scholar] [CrossRef]
- Cho, K.M.; Lee, J.H.; Yun, H.D.; Ahn, B.Y.; Kim, H.; Seo, W.T. Changes of phytochemical constituents (isoflavones, flavanols, and phenolic acids) during cheonggukjang soybeans fermentation using potential probiotics Bacillus subtilis CS90. J. Food Compos Anal. 2011, 24, 402–410. [Google Scholar] [CrossRef]
- Liu, F.; Chen, Z.; Shao, J.; Wang, C.; Zhan, C. Effect of fermentation on the peptide content, phenolics and antioxidant activity of defatted wheat germ. Food Biosci. 2017, 20, 141–148. [Google Scholar] [CrossRef]
- Li, X.; Xing, Y.; Cao, L.; Xu, Q.; Li, S.; Wang, R.; Jiang, Z.; Che, Z.; Lin, H. Effects of six commercial Saccharomyces cerevisiae strains on phenolic attributes, antioxidant activity, and aroma of kiwifruit (Actinidia deliciosa cv.) wine. BioMed Res. Int. 2017, 2017, 2934743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista-Expósitoa, S.; Martínez-Villaluenga, C.; Montserrat, D.; Silván, J.M.; Frias, J.; Peñas, E. Combination of pH-controlled fermentation in mild acidic conditions and enzymatic hydrolysis by Savinase to improve metabolic health-promoting properties of lentil. J. Func. Foods 2018, 48, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.S.; Yu, X.J.; Qin, X.P.; Ma, H.L.; ElGasim, A.; Yagou, A.; Hu, J.L. Hydrolysis of rapeseed meal protein under simulated duodenum digestion: Kinetic modeling and antioxidant activity modeling and antioxidant activity. LWT Food Sci. Technol. 2016, 68, 523–531. [Google Scholar] [CrossRef]
- Roberts, P.R.; Burney, J.D.; Black, K.W.; Zaloga, G.P. Effect of chain length on absorption of biologically active peptides from the gastrointestinal tract. Digestion 1999, 60, 332–337. [Google Scholar] [CrossRef]
- Ajibola, C.F.; Fashakin, J.B.; Fagbemi, T.N.; Aluko, R.E. Effect of peptide size on antioxidant properties of African yam bean seed (Sphenostylis stenocarpa) protein hydrolysate fractions. Int. J. Mol. Sci. 2011, 12, 6685–6702. [Google Scholar] [CrossRef] [Green Version]
- Li, C.J.; Ma, J.W.; Zhao, L.; Zhang, Y.Y.; Yu, Y.H.; Shu, X.Y.; Li, J.; Jia, X.S. Molecular selective binding of basic amino acids by a water-soluble pillar arene. Chem. Comm. 2013, 49, 19–24. [Google Scholar] [CrossRef]
- Yin, H.M.; Zhong, Y.D.; Xia, S.K.; Hu, J.L.; Nie, S.P.; Xiong, T.; Xie, M.Y. Effects of fermentation with Lactobacillus plantarum NCU137 on nutritional, sensory and stability properties of Coix (Coix lachryma-jobi L.) seed. Food Chem. 2020, 314, 126037. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.W.; Wang, R.F.; Hao, X.R.; Duan, Y.X.; Meng, Z.Q.; An, X.P.; Qi, J.W. Dietary fermented soybean meal inclusion improves growth performance and ileal barrier function of the weaned piglets challenged by enterotoxigenic Escherichia coli K88. Anim. Feed Sci. Technol. 2020, 268, 114596. [Google Scholar] [CrossRef]
- Wen, C.T.; Zhang, J.X.; Zhang, H.H.; Duan, Y.Q.; Ma, H.L. Plant protein-derived antioxidant peptides: Isolation, identification, mechanism of action and application in food systems: A review. Trends Food Sci Tech. 2020, 105, 308–322. [Google Scholar] [CrossRef]
- Yu, L.; Li, R.; Liu, W.; Zhou, Y.; Li, Y.; Qin, Y.; Chen, Y.H.; Xu, Y. Protective effects of wheat peptides against ethanol-induced gastric mucosal lesions in rats: Vasodilation and anti-inflammation. Nutrients 2020, 12, 2355. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.; Ryu, B.; Ngo, D.; Kim, S. Peptide isolated from Japanese flounder skin gelatin protects against cellular oxidative damage. J. Agr. Food Chem. 2012, 60, 9112–9119. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agr. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Ruann, J.S.C.; Hélia, H.S. Advantages of an acid protease from Aspergillus oryzae over commercial preparations for production of whey protein hydrolysates with antioxidant activities. Biocatal. Agr. Biotech. 2014, 3, 58–65. [Google Scholar] [CrossRef]
- Pan, M.; Jiang, T.S.; Pan, J.L. Antioxidant Activities of Rapeseed Protein Hydrolysates. Food Bioprocess Tech. 2011, 4, 1144–1152. [Google Scholar] [CrossRef]
- Wang, D.; Wang, L.J.; Zhu, F.X.; Zhu, J.Y.; Chen, X.D.; Zou, L.; Saito, M.; Li, L.T. In vitro and in vivo studies on the antioxidant activities of the aqueous extracts of Douchi (a traditional Chinese salt-fermented soybean food). Food Chem. 2008, 107, 1421–1428. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxid. Med. Cell Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.Z.; Zeng, X.G.; Ding, X.L. Effects of replacing soybean meal with fermented rapeseed meal on performance, serum biochemical variables and intestinal morphology of broilers. Asian Austral. J. Anim. 2012, 25, 1734–1741. [Google Scholar] [CrossRef]
- Tang, J.W.; Sun, H.; Yao, X.H.; Wu, Y.F.; Wang, X.; Feng, J. Effects of replacement of soybean meal by fermented cottonseed meal on growth performance, serum biochemical parameters and immune function of yellow-feathered broilers. Asian Austral. J. Anim. 2012, 25, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.K.; Shang, Q.H.; Wang, Q.Q.; Hu, J.X.; Piao, X.S. Comparative effects of enzymolytic soybean meal and antibiotics in diets on growth performance, antioxidant capacity, immunity, and intestinal barrier function in weaned pigs. Anim. Feed Sci. Technol. 2019, 248, 47–58. [Google Scholar] [CrossRef]
- Luti, S.; Mazzoli, L.; Ramazzotti, M.; Galli, V.; Venturi, M.; Marino, G.; Lehmann, M.; Guerrini, S.; Granchi, L.; Paoli, P.; et al. Antioxidant and anti-inflammatory properties of sourdoughs containing selected Lactobacilli strains are retained in breads. Food Chem. 2020, 322, 126710. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Control Diet | Experiment Diet |
|---|---|---|
| Compositions (g/kg, as fed basis) Soybean meal | 180 | 180 |
| RSM | 50 | − |
| FERSM | − | 50 |
| Corn | 390 | 390 |
| Wheat | 310 | 310 |
| Fish meal | 30 | 30 |
| Stone | 15 | 15 |
| Salt | 5 | 5 |
| Oil | 10 | 10 |
| Premix 1 | 10 | 10 |
| Nutrient level (%) | ||
| CP | 19.22 | 19.359 |
| Energy | 3.796 | 3.796 |
| Fiber | 2.88 | 2.565 |
| Fat | 3.49 | 3.495 |
| Ca | 0.274 | 0.275 |
| p | 0.482 | 0.486 |
| Methionine | 0.322 | 0.322 |
| Lysine | 0.891 | 0.896 |
| Threonine Trp | 0.694 0.219 | 0.694 0.22 |
| Composition | RSM | FRSMP | SEM | p |
|---|---|---|---|---|
| Glucosinolate (µmol/g DM) | 75.17 a | 6.15 b | 15.44 | 0.000 |
| Crude protein(mg/g DM) | 360 b | 405 a | 11.3 | 0.017 |
| NDF (mg/g DM) | 226 a | 140 b | 19.5 | 0.000 |
| Lactic acid (g/kg DM) | 30.50 b | 110.30 a | 24.43 | 0.000 |
| Soluble peptides (mg/g DM) | 53.33 b | 231.05 a | 30.85 | 0.000 |
| Total phenolic content (µg/g DM) | 2.18 | 2.45 | 0.039 | 0.579 |
| Phytic acid (µg/gDM) | 2.68 a | 0.25 b | 0.544 | 0.000 |
| Peptide fraction (kDa) (mg/g DM) | ||||
| <1 kDa | 5.39 b | 80.10 a | 16.70 | 0.000 |
| 1–3 kDa | 0.44 b | 2.14 a | 0.38 | 0.000 |
| 3–5 kDa | 2.85 b | 32.99 a | 6.74 | 0.000 |
| 5–10 kDa | 2.59 b | 56.51 a | 12.06 | 0.000 |
| >10 kDa | 42.08 a | 19.74 b | 5.00 | 0.000 |
| Composition | RSM | FRSMP | SEM | p |
|---|---|---|---|---|
| Glycine | 26.03 b | 79.02 a | 11.87 | 0.000 |
| Alanine | 396.95 a | 318.18 b | 17.65 | 0.000 |
| Serine | 98.31 a | 28.05 b | 15.73 | 0.000 |
| Proline | 529.27 b | 673.09 a | 32.29 | 0.000 |
| Valine | 182.34 a | 389.77 b | 46.42 | 0.000 |
| Threonine | 107.84 a | 90.69 b | 4.21 | 0.012 |
| Cysteine | 6.33 b | 17.83 a | 2.61 | 0.000 |
| Isoleucine | 79.34 a | 281.1 b | 45.14 | 0.000 |
| Asparagine | 219.76 a | 31.04 b | 42.21 | 0.000 |
| Aspartic acid | 402.52 a | 167.63 b | 52.61 | 0.000 |
| Glutamine | 37.13 a | 26.46 b | 2.52 | 0.004 |
| Glutamate | 708.95 b | 2052.08 a | 300.57 | 0.000 |
| Methionine | 3.73 b | 454.33 a | 100.79 | 0.000 |
| Histidine | 21.99 | 24.64 | 0.78 | 0.083 |
| Phenylalanine | 99.25 b | 627.23 a | 118.15 | 0.000 |
| Arginine | 318.19 a | 111.00 b | 46.35 | 0.000 |
| Tryptophan | 85.64 b | 171.22 a | 19.16 | 0.000 |
| Lysine | 154.15 b | 1261.30 a | 247.71 | 0.000 |
| Tyrosine | 50.05 b | 500.66 a | 100.78 | 0.000 |
| Leucine | 103.02 b | 608.99 a | 113.14 | 0.000 |
| Total amino acids | 3630.80 a | 7914.31 b | 958.35 | 0.000 |
| EAA | 815.32 b | 3884.63 a | 686.53 | 0.000 |
| EAA/NEAA | 0.29 b | 0.97 a | 0.059 | 0.000 |
| Item | RSM | FRSMP | SEM | p |
|---|---|---|---|---|
| DMD (g/kg) | 700 b | 779 a | 17.92 | 0.000 |
| CPD (g/kg) | 861 b | 880 a | 4.67 | 0.012 |
| DMI (g/d) | 18.06 a | 18.76 a | 0.28 | 0.088 |
| ADG (g/d) | 7.56 b | 8.46 a | 0.24 | 0.034 |
| FCR | 2.39 | 2.22 | 0.05 | 0.115 |
| Serum | ||||
| GH (ng/mL) | 5.19 | 6.15 | 0.24 | 0.260 |
| IGF-1 (ng/mL) | 167.11 a | 204.75 b | 5.86 | 0.011 |
| T3 (ng/mL) | 0.46 b | 0.60 a | 0.03 | 0.014 |
| T4 (ng/mL) | 45.39 b | 66.16 a | 4.49 | 0.004 |
| Tissue | ||||
| IGF-1 (ng/mg) | 10.82 b | 17.02 a | 0.89 | 0.000 |
| Item | RSM | FRSMP | SEM | p |
|---|---|---|---|---|
| Serum | ||||
| IgA (g/L) | 0.49 | 0.56 | 0.02 | 0.087 |
| IgM (g/L) | 0.42 b | 0.55 a | 0.02 | 0.000 |
| IgG (g/L) | 5.45 b | 6.33 a | 0.16 | 0.028 |
| IL-1β (pg/mL) | 38.67 a | 34.32 b | 1.32 | 0.089 |
| IL-6 (pg/mL) | 227.93 a | 155.68 b | 8.50 | 0.000 |
| Tissue | ||||
| SigA (µg/mg) | 0.54 b | 0.68 a | 0.02 | 0.039 |
| IL-1β (pg/mg) | 3.32 | 2.96 | 0.09 | 0.316 |
| IL-6 (pg/mg) | 22.65 a | 16.99 b | 0.68 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Sun, H.; Liu, X. A Novel Fermented Rapeseed Meal, Inoculated with Selected Protease-Assisting Screened B. subtilis YY-4 and L. plantarum 6026, Showed High Availability and Strong Antioxidant and Immunomodulation Potential Capacity. Foods 2022, 11, 2118. https://doi.org/10.3390/foods11142118
Wang Y, Sun H, Liu X. A Novel Fermented Rapeseed Meal, Inoculated with Selected Protease-Assisting Screened B. subtilis YY-4 and L. plantarum 6026, Showed High Availability and Strong Antioxidant and Immunomodulation Potential Capacity. Foods. 2022; 11(14):2118. https://doi.org/10.3390/foods11142118
Chicago/Turabian StyleWang, Yan, Hao Sun, and Xiaolan Liu. 2022. "A Novel Fermented Rapeseed Meal, Inoculated with Selected Protease-Assisting Screened B. subtilis YY-4 and L. plantarum 6026, Showed High Availability and Strong Antioxidant and Immunomodulation Potential Capacity" Foods 11, no. 14: 2118. https://doi.org/10.3390/foods11142118