
Functional and Allergenic Properties Assessment of Conalbumin (Ovotransferrin) after Oxidation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Human Sera
2.3. Preparation of Oxidation Samples
2.4. Electrophoretic Analysis
2.5. Structure Analysis
2.5.1. Intrinsic Fluorescence Measurements
2.5.2. Secondary Structure Analysis
2.6. Thermal Stability Analysis
2.7. Functional Properties Analysis
2.7.1. Emulsifying Properties
2.7.2. Foaming properties
2.8. Assessment of In Vitro Allergenicity on CA Oxidation Complexes
2.8.1. Western Blot
2.8.2. ELISA Analysis
2.8.3. Assays of KU812 Cells
2.9. Assay of Dendritic Cells (DCs)
2.10. In Vitro Digestion Stability Assay
2.11. Statistical Analysis
3. Results
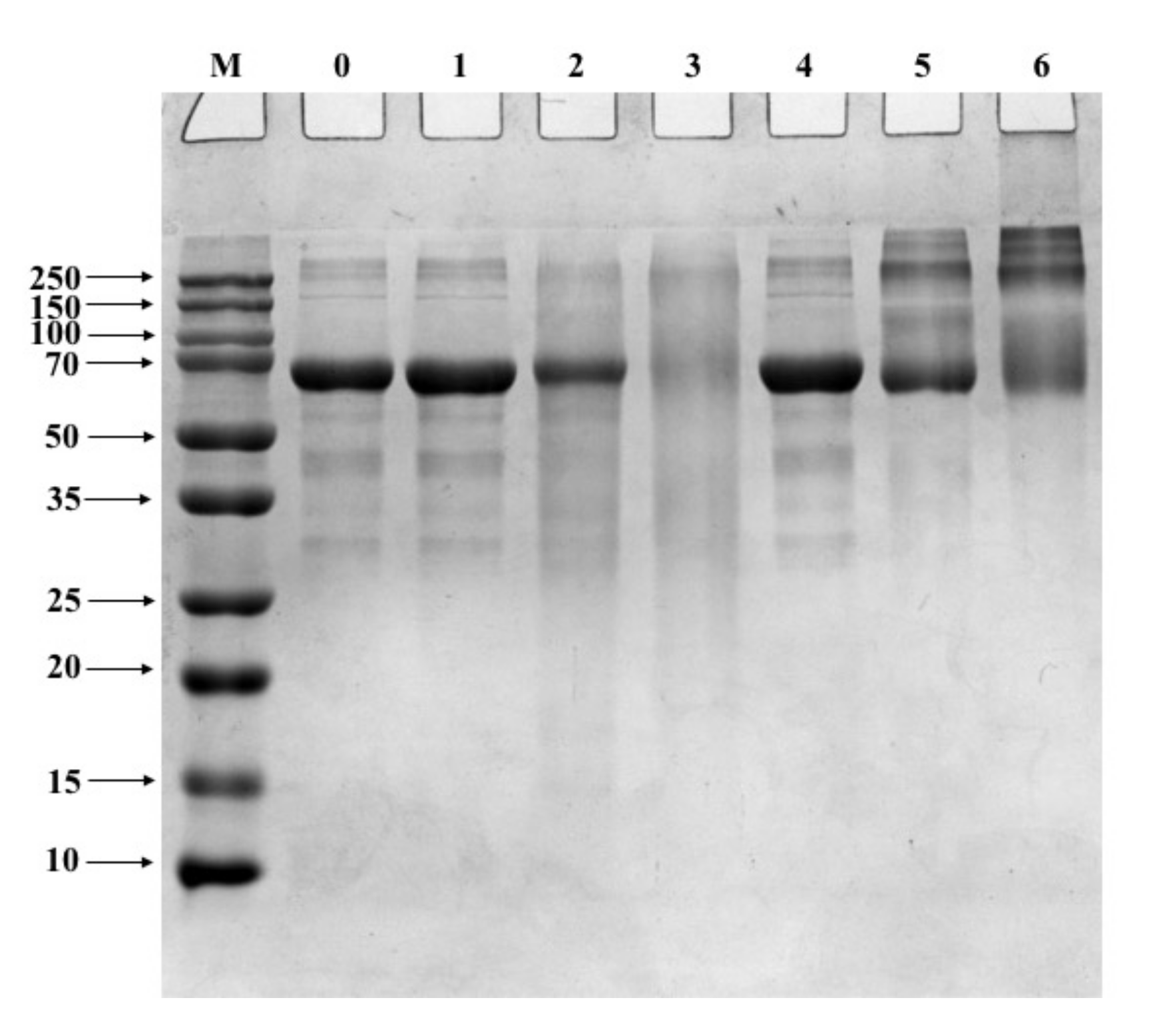
3.1. SDS-PAGE
3.2. Effect of Oxidation on CA Structure
3.3. Function Properties
3.4. Stability of CA after Oxidation in a Simulated Digestion In Vitro
3.5. Uptake by DCs
3.6. In Vitro Allergenicity Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiménez-Saiz, R.; Ruiz-Henestrosa, VMP; López-Fandiño, R.; Molina, E. In vitro digestibility and allergenicity of emulsified hen egg. Food Res. Int. 2012, 48, 404–409. [Google Scholar] [CrossRef] [Green Version]
- Abeyrathne, E.D.; Lee, H.Y.; Ahn, D.U. Egg white proteins and their potential use in food processing or as nutraceutical and pharmaceutical agents—A review. Poult. Sci. 2013, 92, 3292–3299. [Google Scholar] [CrossRef]
- Wang, J.; Chi, Y.; Cheng, Y.; Zhao, Y. Physicochemical properties, in vitro digestibility and antioxidant activity of dry-heated egg white protein. Food Chem. 2018, 246, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Vanga, S.K.; Wang, J.; Raghavan, V. Effects of Ultrasonic and Microwave Processing on Avidin Assay and Secondary Structures of Egg White Protein. Food Bioprocess Technol. 2018, 11, 1974–1984. [Google Scholar] [CrossRef]
- Yamada, H.; Nagae, T.; Watanabe, N. High-pressure protein crystallography of hen egg-white lysozyme. Acta. Crystallogr. D. Biol. Crystallogr. 2015, 71 Pt 4, 742–753. [Google Scholar] [CrossRef] [Green Version]
- Enomoto, H.; Nagae, S.; Hayashi, Y.; Li, C.P.; Aoki, T. Improvement of functional properties of egg white protein through glycation and phosphorylation by dry-heating. Asian Australas. J. Anim. Sci. 2009, 22, 591–597. [Google Scholar] [CrossRef]
- Savage, J.H.; Matsui, E.C.; Skripak, J.M.; Wood, R.A. The natural history of egg allergy. J. Allergy Clin. Immunol. 2007, 120, 1413–1417. [Google Scholar] [CrossRef]
- Anvari, S.; Miller, J.; Yeh, C.Y.; Davis, C.M. IgE-Mediated Food Allergy. Clin. Rev. Allergy Immunol. 2019, 57, 244–260. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Perez, T.I.; Zuidhof, M.J.; Renema, R.A.; Wu, J. Oxidative stability of omega-3 polyunsaturated fatty acids enriched eggs. J. Agric. Food Chem. 2013, 61, 11595–11602. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, C.; Kong, X.; Hua, Y. Oxidative modification of soy protein by peroxyl radicals. Food Chem. 2009, 116, 295–301. [Google Scholar] [CrossRef]
- Uchida, K. Aldehyde adducts generated during lipid peroxidation modification of proteins. Free Radic. Res. 2015, 49, 896–904. [Google Scholar] [CrossRef]
- Naveena, B.M.; Faustman, C.; Tatiyaborworntham, N.; Yin, S.; Ramanathan, R.; Mancini, R.A. Detection of 4-hydroxy-2-nonenal adducts of turkey and chicken myoglobins using mass spectrometry. Food Chem. 2010, 122, 836–840. [Google Scholar] [CrossRef]
- Lee, S.; Joo, S.T.; Alderton, A.L.; Hill, D.W.; Faustman, C. Oxymyoglobin and lipid oxidation in yellowfin tuna (Thunnus albacares) loins. J. Food Sci. 2003, 68, 1664–1668. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, S.; Ahn, D.U. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, Z.; Lin, H.; Du, S.; Zhu, Z. Effect of malondialdehyde treatment on the IgE binding capacity and conformational structure of shrimp tropomyosin. Food Chem. 2015, 175, 374–380. [Google Scholar] [CrossRef]
- Lv, L.; Lin, H.; Li, Z.; Yuan, F.; Gao, Q.; Ma, J. Effect of 4-hydroxy-2-nonenal treatment on the IgE binding capacity and structure of shrimp (Metapenaeus ensis) tropomyosin. Food Chem. 2016, 212, 313–322. [Google Scholar] [CrossRef]
- Abeyrathne, E.; Lee, H.; Ahn, D. Sequential separation of lysozyme, ovomucin, ovotransferrin, and ovalbumin from egg white. Poult. Sci. 2014, 93, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Kranthi, V.S.; Jin, W.; Vijaya, R. Impact of food processing on the structural and allergenic properties of egg white. Trends Food Sci. Technol. 2018, 78, 188–196. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Lv, L.; Lin, H.; Li, Z.; Wang, J.; Ahmed, I.; Chen, H. Changes of structure and IgE binding capacity of shrimp (Metapenaeus ensis) tropomyosin followed by acrolein treatment. Food Funct. 2017, 8, 1028–1036. [Google Scholar] [CrossRef]
- Liu, G.; Tu, Z.; Wang, H.; Zhang, L.; Huang, T.; Ma, D. Monitoring of the functional properties and unfolding change of Ovalbumin after DHPM treatment by HDX and FTICR MS. Food Chem. 2017, 227, 413–421. [Google Scholar] [CrossRef]
- Huang, X.; Tu, Z.; Wang, H.; Zhang, Q.; Shi, Y.; Xiao, H. Increase of ovalbumin glycation by the Maillard reaction after disruption of the disulfide bridge evaluated by liquid chromatography and high resolution mass spectrometry. J. Agric. Food Chem. 2013, 61, 2253–2262. [Google Scholar] [CrossRef]
- Liu, Y.; Ying, D.; Cai, Y.; Le, X. Improved antioxidant activity and physicochemical properties of curcumin by adding ovalbumin and its structural characterization. Food Hydrocoll. 2017, 72, 304–311. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Z.; Li, K.; Li, X.; Yang, A.; Tong, P.; Chen, H. Allergenicity assessment on thermally processed peanut influenced by extraction and assessment methods. Food Chem. 2019, 281, 130–139. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhang, T.; Velickovic, T.C.; Li, S.; Wu, X. Covalent conjugation with -epigallo-catechin 3-gallate and chlorogenic acid changes allergenicity and functional properties of ara h1 from peanut. Food Chem. 2020, 331, 127355. [Google Scholar] [CrossRef]
- Lv, L.; Lin, H.; Li, Z.; Nayak, B.; Ahmed, I. Structural changes of 2, 2′-azobi (2-amidinopropane) dihydrochloride (AAPH) treated shrimp tropomyosin decrease allergenicity. Food Chem. 2019, 274, 547–557. [Google Scholar] [CrossRef]
- Wu, W.; Wu, X.; Hua, Y. Structural modification of soy protein by the lipid peroxidation product acrolein. LWT Food Sci. Technol. 2010, 43, 133–140. [Google Scholar] [CrossRef]
- Li, Z.; Lin, H.; Cao, L.M.; Jamil, K. The influence of gamma irradiation on the allergenicity of shrimp (Penaeus vannamei). J. Food Eng. 2007, 79, 945–949. [Google Scholar] [CrossRef]
- Li, Z.; Lin, H.; Pawar, R.; Li, Z.X.; Li, M.H. Mapping IgE binding epitopes of major shrimp (Penaeus monodon) allergen with immunoinformatics tools. Food Chem. Toxicol. 2011, 49, 2954–2960. [Google Scholar] [CrossRef]
- Yuan, F.; Lv, L.; Li, Z.; Mi, N.; Chen, H.; Lin, H. Effect of transglutaminase catalyzed glycosylation on the allergenicity and conformational structure of shrimp (Metapenaeus ensis) tropomyosin. Food Chem. 2017, 219, 215–222. [Google Scholar] [CrossRef]
- Liu, G.; Xiong, Y.L.; Butterfield, D.A. Chemical, physical, and gel-forming properties of oxidized myofibrils and whey-and soyprotein isolates. J. Food Sci. 2000, 65, 811–818. [Google Scholar] [CrossRef]
- Sarker, M.; Rose, J.; McDonald, M.; Morrow, M.R.; Booth, V. Modifications to surfactant protein B structure and lipid interactions under respiratory distress conditions: Consequences of tryptophan oxidation. Biochemistry 2010, 50, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Narayan, M.; Welker, E.; Wedemeyer, W.J.; Scheraga, H.A. Oxidative folding of proteins. Accounts Chem. Res. 2000, 33, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Roos, Y.H.; Biliaderis, C.G.; Miao, S. Food emulsions as delivery systems for flavor compounds: A review. Crit. Rev. Food Sci. 2017, 57, 3173–3187. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhao, M.; Sun, W.; Zhao, G.; Ren, J. Effects of microfluidization treatment and transglutaminase cross-linking on physicochemical, functional, and conformational properties of peanut protein isolate. J. Agric. Food Chem. 2011, 59, 8886–8894. [Google Scholar] [CrossRef]
- Mikita, C.P.; Padlan, E.A. Why is there a greater incidence of allergy to the tropomyosin of certain animals than to that of others? Med. Hypotheses 2007, 69, 1070–1073. [Google Scholar] [CrossRef]
- Refsgaard, H.H.; Tsai, L.; Stadtman, E.R. Modifications of proteins by polyunsaturated fatty acid peroxidation products. Proc. Natl. Acad. Sci. USA 2000, 97, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Suprun, M.; Sicherer, S.H.; Wood, R.A.; Jones, S.M.; Leung, D.Y.M.; Burks, A.W.; Dunkin, D.; Witmer, M.; Grishina, G.; Getts, R.; et al. Mapping Sequential IgE-Binding Epitopes on Major and Minor Egg Allergens. Int. Arch. Allergy Immunol. 2022, 183, 249–261. [Google Scholar] [CrossRef]
- Passante, E.; Frankish, N. The RBL-2H3 cell line: Its provenance and suitability as a model for the mast cell. Inflamm. Res. 2009, 58, 737–745. [Google Scholar] [CrossRef]
- Zhu, J.; Deng, H.; Yang, A.; Wu, Z.; Li, X. Effect of microbial transglutaminase cross-linking on the quality characteristics and potential allergenicity of tofu. Food Funct. 2019, 10, 5485–5497. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Xu, H.; Wu, Y.; Li, X.; Zou, L.; Gao, J.; Chen, H. Alpha7-nicotinic acetylcholine receptors involve the imidacloprid-induced inhibition of IgE-mediated rat and human mast cell activation. RSC Adv. 2017, 7, 51896–51906. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Li, X.; Gao, J.; Chen, H. Characterization of the potential allergenicity of irradiated bovine α-lactalbumin in a BALB/c mouse model. Food Chem. Toxicol. 2016, 97, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Yoon, T.J.; Kim, H.H.; Han, Y.S.; Choi, H. Changes in the antigenicity and allergenicity of ovalbumin in chicken egg white by N-acetylglucosaminidase. Food Chem. 2017, 217, 342–345. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, L.; Ye, L.; Lin, X.; Li, L.; Chen, J.; Yue, W.; Wu, X. Functional and Allergenic Properties Assessment of Conalbumin (Ovotransferrin) after Oxidation. Foods 2022, 11, 2308. https://doi.org/10.3390/foods11152308
Lv L, Ye L, Lin X, Li L, Chen J, Yue W, Wu X. Functional and Allergenic Properties Assessment of Conalbumin (Ovotransferrin) after Oxidation. Foods. 2022; 11(15):2308. https://doi.org/10.3390/foods11152308
Chicago/Turabian StyleLv, Liangtao, Liying Ye, Xiao Lin, Liuying Li, Jiamin Chen, Wenqi Yue, and Xuli Wu. 2022. "Functional and Allergenic Properties Assessment of Conalbumin (Ovotransferrin) after Oxidation" Foods 11, no. 15: 2308. https://doi.org/10.3390/foods11152308
APA StyleLv, L., Ye, L., Lin, X., Li, L., Chen, J., Yue, W., & Wu, X. (2022). Functional and Allergenic Properties Assessment of Conalbumin (Ovotransferrin) after Oxidation. Foods, 11(15), 2308. https://doi.org/10.3390/foods11152308