
Wheat Amylase Trypsin Inhibitors Aggravate Intestinal Inflammation Associated with Celiac Disease Mediated by Gliadin in BALB/c Mice

 ,
,  , , and
, , and
Abstract
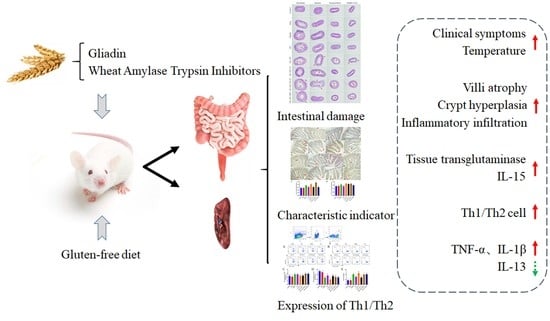
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animals and Diets
2.3. Animal Experiment
2.4. Histological Observation
2.5. Immunohistochemistry of Jejunum Tissue Sections
2.6. Splenocyte Culture In Vitro and Cytokine Analysis
2.7. Th1 and Th2 Cell Subsets of Splenocytes
2.8. Statistical Analysis
3. Results
3.1. Weight, Water Consumption, Clinical Symptoms and Body Temperature of Mice
3.2. Histology of Intestine Tissue Section
3.3. Immunohistochemistry of The Sections of Jejunum Tissue
3.4. Differentiation and Homeostasis of Th1/Th2
3.5. The Levels of Splenocyte Cytokines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hazard, B.; Trafford, K.; Lovegrove, A.; Griffiths, S.; Uauy, C.; Shewry, P. Strategies to improve wheat for human health. Nat. Food 2020, 1, 475–480. [Google Scholar] [CrossRef]
- De Sousa, T.; Ribeiro, M.; Sabença, C.; Igrejas, G. The 10,000-Year Success Story of Wheat! Foods 2021, 10, 2124. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef]
- Sabença, C.; Ribeiro, M.; Sousa, T.D.; Poeta, P.; Bagulho, A.S.; Igrejas, G. Wheat/Gluten-Related Disorders and Gluten-Free Diet Misconceptions: A Review. Foods 2021, 10, 1765. [Google Scholar] [CrossRef]
- Sander, I.; Rihs, H.-P.; Doekes, G.; Quirce, S.; Krop, E.; Rozynek, P.; van Kampen, V.; Merget, R.; Meurer, U.; Brüning, T.; et al. Component-resolved diagnosis of baker’s allergy based on specific IgE to recombinant wheat flour proteins. J. Allergy Clin. Immunol. 2015, 135, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Pronin, D.; Börner, A.; Anne Scherf, K. Old and modern wheat (Triticum aestivum L.) cultivars and their potential to elicit celiac disease. Food Chem. 2020, 339, 127952. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, Y.; Yuan, J.; Yuan, J.; Wang, Z.; Gao, J.; Chen, H. Characterization of AFA01 Capable of Degrading Gluten and Celiac-Immunotoxic Peptides. Foods 2021, 10, 1725. [Google Scholar] [CrossRef]
- Martucciello, S.; Sposito, S.; Esposito, C.; Paolella, G.; Caputo, I. Interplay between Type 2 Transglutaminase (TG2), Gliadin Peptide 31-43 and Anti-TG2 Antibodies in Celiac Disease. Int. J. Mol. Sci. 2020, 21, 3673. [Google Scholar] [CrossRef]
- Zhou, C.; Gao, F.; Gao, J.; Yuan, J.; Lu, J.; Sun, Z.; Xu, M.; Engel, J.; Hui, W.; Gilissen, L.; et al. Prevalence of coeliac disease in Northwest China: Heterogeneity across Northern Silk road ethnic populations. Aliment. Pharm. 2020, 51, 1116–1129. [Google Scholar] [CrossRef]
- Iversen, R.; Amundsen, S.F.; Kleppa, L.; du Pré, M.F.; Stamnaes, J.; Sollid, L.M. Evidence That Pathogenic Transglutaminase 2 in Celiac Disease Derives from Enterocytes. Gastroenterology 2020, 159, 788–790. [Google Scholar] [CrossRef]
- Harris, L.A.; Park, J.Y.; Voltaggio, L.; Lam-Himlin, D. Celiac disease: Clinical, endoscopic, and histopathologic review. Gastrointest. Endosc. 2012, 76, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Abadie, V.; Kim, S.M.; Lejeune, T.; Palanski, B.A.; Ernest, J.D.; Tastet, O.; Voisine, J.; Discepolo, V.; Marietta, E.V.; Hawash, M.B.F.; et al. IL-15, gluten and HLA-DQ8 drive tissue destruction in coeliac disease. Nature 2020, 578, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Junker, Y.; Zeissig, S.; Kim, S.J.; Barisani, D.; Wieser, H.; Leffler, D.A.; Zevallos, V.; Libermann, T.A.; Dillon, S.; Freitag, T.L.; et al. Wheat amylase trypsin inhibitors drive intestinal inflammation via activation of toll-like receptor 4. J. Exp. Med. 2012, 209, 2395–2408. [Google Scholar] [CrossRef]
- Geisslitz, S.; Ludwig, C.; Scherf, K.A.; Koehler, P. Targeted LC-MS/MS Reveals Similar Contents of α-Amylase/Trypsin-Inhibitors as Putative Triggers of Nonceliac Gluten Sensitivity in all Wheat Species except Einkorn. J. Agric. Food Chem. 2018, 66, 12395–12403. [Google Scholar] [CrossRef] [PubMed]
- Zevallos, V.F.; Raker, V.K.; Maxeiner, J.; Scholtes, P.; Steinbrink, K.; Schuppan, D. Dietary wheat amylase trypsin inhibitors exacerbate murine allergic airway inflammation. Eur. J. Nutr. 2019, 58, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Bellinghausen, I.; Weigmann, B.; Zevallos, V.; Maxeiner, J.; Reissig, S.; Waisman, A.; Schuppan, D.; Saloga, J. Wheat amylase-trypsin inhibitors exacerbate intestinal and airway allergic immune responses in humanized mice. J. Allergy Clin. Immunol. 2019, 143, 201–212.e204. [Google Scholar] [CrossRef]
- Tundo, S.; Lupi, R.; Lafond, M.; Giardina, T.; Larre, C.; Denery-Papini, S.; Morisset, M.; Kalunke, R.; Sestili, F.; Masci, S. Wheat ATI CM3, CM16 and 0.28 Allergens Produced in Pichia Pastoris Display a Different Eliciting Potential in Food Allergy to Wheat (double dagger). Plants 2018, 7, 101. [Google Scholar] [CrossRef] [Green Version]
- Neumann, J.; Ziegler, K.; Gelleri, M.; Frohlich-Nowoisky, J.; Liu, F.; Bellinghausen, I.; Schuppan, D.; Birk, U.; Poschl, U.; Cremer, C.; et al. Nanoscale distribution of TLR4 on primary human macrophages stimulated with LPS and ATI. Nanoscale 2019, 11, 9769–9779. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P. What Is Gluten—Why Is It Special? Front. Nutr. 2019, 6, 101. [Google Scholar] [CrossRef]
- Zevallos, V.F.; Raker, V.; Tenzer, S.; Jimenez-Calvente, C.; Ashfaq-Khan, M.; Russel, N.; Pickert, G.; Schild, H.; Steinbrink, K.; Schuppan, D. Nutritional Wheat Amylase-Trypsin Inhibitors Promote Intestinal Inflammation via Activation of Myeloid Cells. Gastroenterology 2017, 152, 1100–1113 e1112. [Google Scholar] [CrossRef] [Green Version]
- Caminero, A.; McCarville, J.L.; Zevallos, V.F.; Pigrau, M.; Yu, X.B.; Jury, J.; Galipeau, H.J.; Clarizio, A.V.; Casqueiro, J.; Murray, J.A.; et al. Lactobacilli Degrade Wheat Amylase Trypsin Inhibitors to Reduce Intestinal Dysfunction Induced by Immunogenic Wheat Proteins. Gastroenterology 2019, 156, 2266–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Schuppan, D.; Rojas Tovar, L.E.; Zevallos, V.F.; Loponen, J.; Gänzle, M. Sourdough Fermentation Degrades Wheat Alpha-Amylase/Trypsin Inhibitor (ATI) and Reduces Pro-Inflammatory Activity. Foods 2020, 9, 943. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Koch, R.; Moschen, A.R. Proinflammatory wheat attacks on the intestine: Alpha-amylase trypsin inhibitors as new players. Gastroenterology 2013, 144, 1561–1563. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Hui, J.; Lu, Q.; Yang, A.; Yuan, J.; Gao, J.; Wu, Z.; Li, X.; Tong, P.; Chen, H. Effect of transglutaminase cross-linking on the allergenicity of tofu based on a BALB/c mouse model. Food Funct. 2020, 11, 404–413. [Google Scholar] [CrossRef]
- Vijaykrishnaraj, M.; Mohan Kumar, B.V.; Muthukumar, S.P.; Kurrey, N.K.; Prabhasankar, P. Antigen-Specific Gut Inflammation and Systemic Immune Responses Induced by Prolonging Wheat Gluten Sensitization in BALB/c Murine Model. J. Proteome Res. 2017, 16, 3514–3528. [Google Scholar] [CrossRef]
- Wickham, M.; Faulks, R.; Mills, C. In vitro digestion methods for assessing the effect of food structure on allergen breakdown. Mol. Nutr. Food Res. 2009, 53, 952–958. [Google Scholar] [CrossRef]
- Reyes-Pavón, D.; Cervantes-García, D.; Bermúdez-Humarán, L.G.; Córdova-Dávalos, L.E.; Quintanar-Stephano, A.; Jiménez, M.; Salinas, E. Protective Effect of Glycomacropeptide on Food Allergy with Gastrointestinal Manifestations in a Rat Model through Down-Regulation of Type 2 Immune Response. Nutrients 2020, 12, 2942. [Google Scholar] [CrossRef]
- Ladics, G.S.; Fry, J.; Goodman, R.; Herouet-Guicheney, C.; Hoffmann-Sommergruber, K.; Madsen, C.B.; Penninks, A.; Pomés, A.; Roggen, E.L.; Smit, J.; et al. Allergic sensitization: Screening methods. Clin. Transl. Allergy 2014, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Navarro, S.; Lopata, A.L. Current advances of murine models for food allergy. Mol. Immunol. 2016, 70, 104–117. [Google Scholar] [CrossRef]
- Oyoshi, M.K.; Oettgen, H.C.; Chatila, T.A.; Geha, R.S.; Bryce, P.J. Food allergy: Insights into etiology, prevention, and treatment provided by murine models. J. Allergy Clin. Immunol. 2014, 133, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Lundin, K.E.A.; Wijmenga, C. Coeliac disease and autoimmune disease-genetic overlap and screening. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Leffler, D.A.; Green, P.H.R.; Fasano, A. Extraintestinal manifestations of coeliac disease. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Guilherme, M.; Zevallos, V.F.; Pesi, A.; Stoye, N.M.; Nguyen, V.T.T.; Radyushkin, K.; Schwiertz, A.; Schmitt, U.; Schuppan, D.; Endres, K. Dietary Wheat Amylase Trypsin Inhibitors Impact Alzheimer’s Disease Pathology in 5xFAD Model Mice. Int. J. Mol. Sci. 2020, 21, 6288. [Google Scholar] [CrossRef]
- Bose, U.; Juhász, A.; Broadbent, J.A.; Byrne, K.; Howitt, C.A.; Colgrave, M.L. Identification and Quantitation of Amylase Trypsin Inhibitors Across Cultivars Representing the Diversity of Bread Wheat. J. Proteome Res. 2020, 19, 2136–2148. [Google Scholar] [CrossRef] [PubMed]
- Pickert, G.; Wirtz, S.; Matzner, J.; Ashfaq-Khan, M.; Heck, R.; Rosigkeit, S.; Thies, D.; Surabattula, R.; Ehmann, D.; Wehkamp, J.; et al. Wheat Consumption Aggravates Colitis in Mice via Amylase Trypsin Inhibitor-mediated Dysbiosis. Gastroenterology 2020, 159, 257–272.e17. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, K.; Neumann, J.; Liu, F.; Fröhlich-Nowoisky, J.; Cremer, C.; Saloga, J.; Reinmuth-Selzle, K.; Pöschl, U.; Schuppan, D.; Bellinghausen, I.; et al. Nitration of Wheat Amylase Trypsin Inhibitors Increases Their Innate and Adaptive Immunostimulatory Potential. Front. Immunol. 2018, 9, 3174. [Google Scholar] [CrossRef] [Green Version]
- Lebwohl, B.; Sanders, D.S.; Green, P.H.R. Coeliac disease. Lancet 2018, 391, 70–81. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Vanoli, A.; Giuffrida, P.; Luinetti, O.; Solcia, E.; Corazza, G.R. The function of tissue transglutaminase in celiac disease. Autoimmun. Rev. 2012, 11, 746–753. [Google Scholar] [CrossRef]
- Freitag, T.; Schulze-Koops, H.; Niedobitek, G.; Melino, G.; Schuppan, D. The role of the immune response against tissue transglutaminase in the pathogenesis of coeliac disease. Autoimmun. Rev. 2004, 3, 13–20. [Google Scholar] [CrossRef]
- Abadie, V.; Jabri, B. IL-15: A central regulator of celiac disease immunopathology. Immunol. Rev. 2014, 260, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Annunziato, F.; Romagnani, C.; Romagnani, S. The 3 major types of innate and adaptive cell-mediated effector immunity. J. Allergy Clin. Immunol. 2015, 135, 626–635. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, T.; Hu, S.; Min, F.; Li, J.; Shen, Y.; Yuan, J.; Gao, J.; Wu, Y.; Chen, H. Wheat Amylase Trypsin Inhibitors Aggravate Intestinal Inflammation Associated with Celiac Disease Mediated by Gliadin in BALB/c Mice. Foods 2022, 11, 1559. https://doi.org/10.3390/foods11111559
Yu T, Hu S, Min F, Li J, Shen Y, Yuan J, Gao J, Wu Y, Chen H. Wheat Amylase Trypsin Inhibitors Aggravate Intestinal Inflammation Associated with Celiac Disease Mediated by Gliadin in BALB/c Mice. Foods. 2022; 11(11):1559. https://doi.org/10.3390/foods11111559
Chicago/Turabian StyleYu, Tian, Shuai Hu, Fangfang Min, Jingjing Li, Yunpeng Shen, Juanli Yuan, Jinyan Gao, Yong Wu, and Hongbing Chen. 2022. "Wheat Amylase Trypsin Inhibitors Aggravate Intestinal Inflammation Associated with Celiac Disease Mediated by Gliadin in BALB/c Mice" Foods 11, no. 11: 1559. https://doi.org/10.3390/foods11111559
APA StyleYu, T., Hu, S., Min, F., Li, J., Shen, Y., Yuan, J., Gao, J., Wu, Y., & Chen, H. (2022). Wheat Amylase Trypsin Inhibitors Aggravate Intestinal Inflammation Associated with Celiac Disease Mediated by Gliadin in BALB/c Mice. Foods, 11(11), 1559. https://doi.org/10.3390/foods11111559