
Dietary Supplementation with Epicatechin Improves Intestinal Barrier Integrity in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Management and Diet
2.2. Assessment of Weight Gain
2.3. Sample Collection
2.4. Measurement of Serum Parameters
2.5. Small Intestine Biochemical Analysis
2.5.1. Preparation of Intestinal Homogenates
2.5.2. Determination of Intestinal Antioxidant Capacity
2.5.3. Assessment of Intestinal Cytokine and Cell Apoptosis-Related Protein Contents
2.6. Detection of Intestinal Cell Apoptosis
2.7. Western Blot Assay
2.8. Statistical Analysis
3. Results
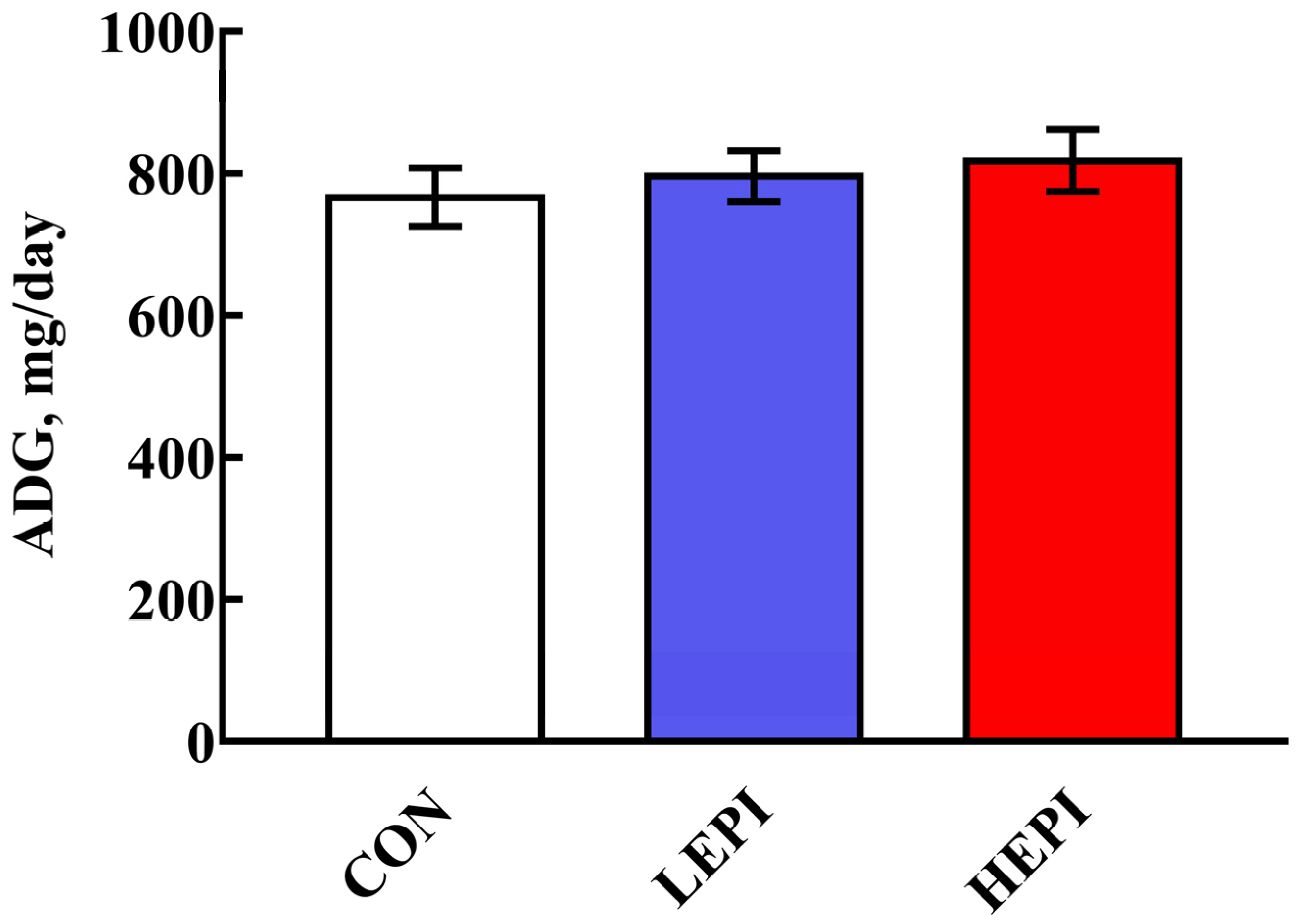
3.1. Growth Performance
3.2. Serum Indices
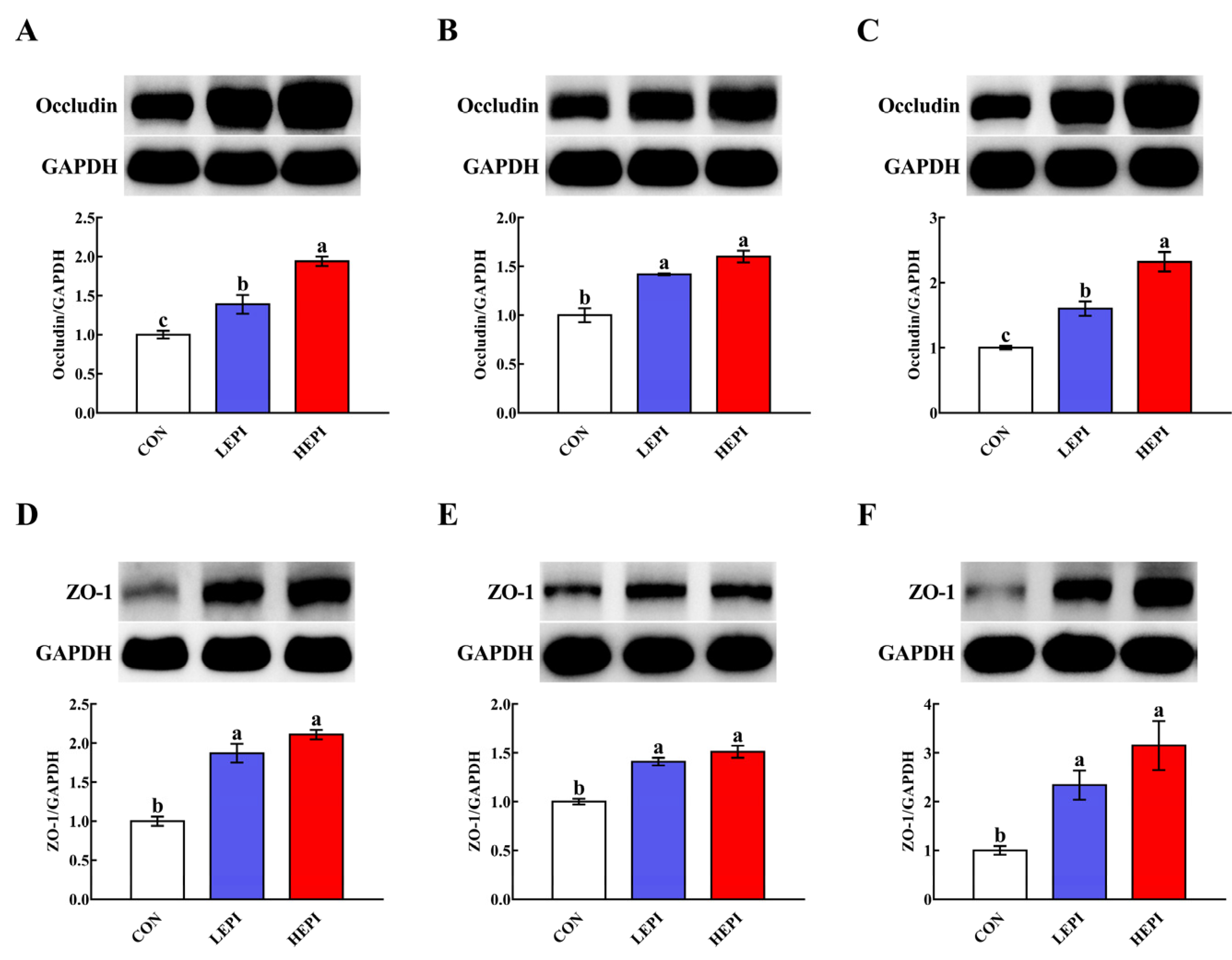
3.3. Abundance of Intestinal Tight Junction Proteins
3.4. Intestinal Antioxidant Capacity
3.5. Intestinal Cytokine Contents
3.6. Intestinal Cell Apoptosis
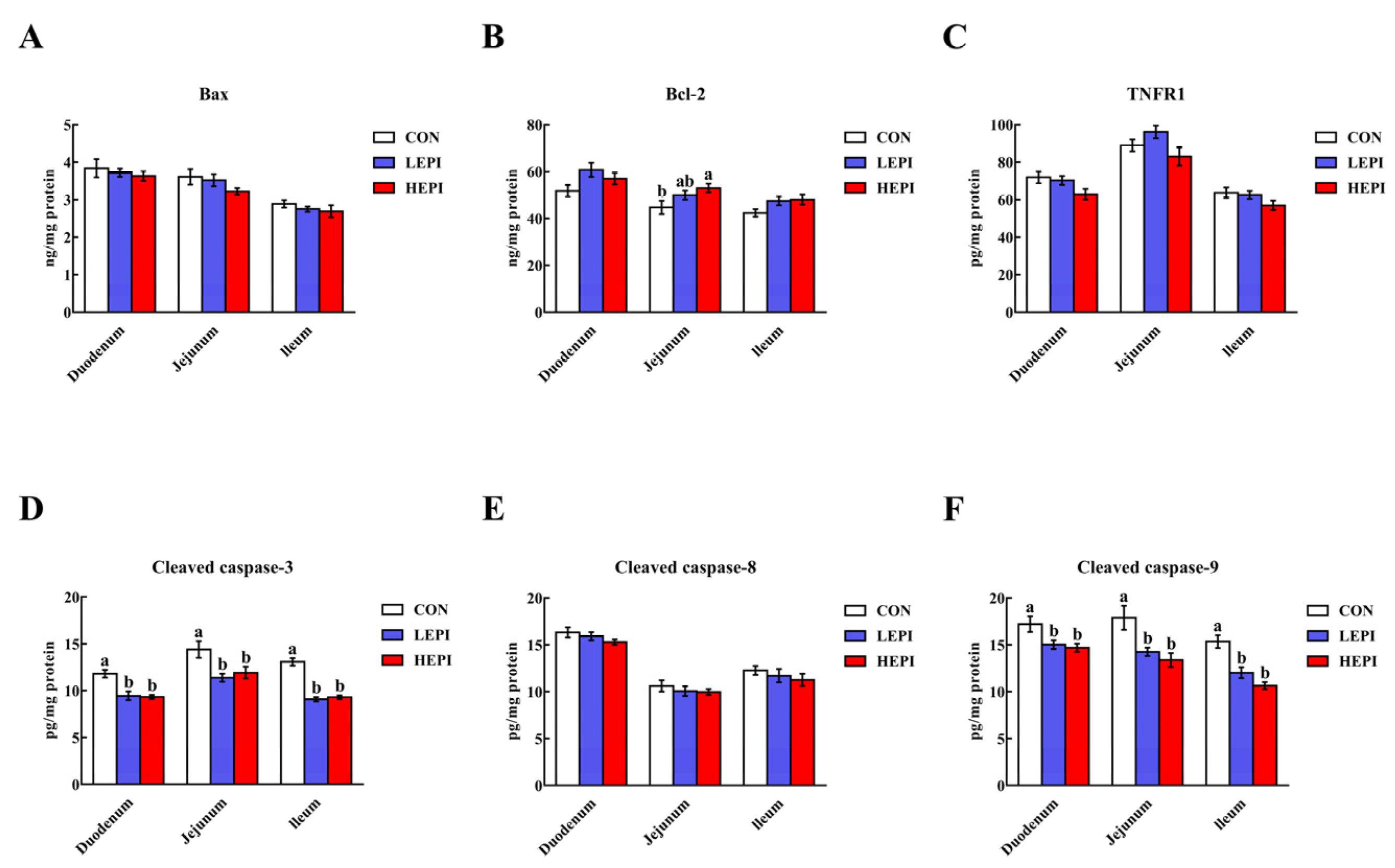
3.7. Intestinal Cell Apoptosis-Related Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Wijtten, P.J.A.; van der Meulen, J.; Verstegen, M.W.A. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Banan, A.; Fields, J.; Keshavarzian, A. Intestinal barrier: An interface between health and disease. J. Gastroenterol. Hepatol. 2003, 18, 479–497. [Google Scholar] [CrossRef]
- Blikslager, A.T.; Moeser, A.J.; Gookin, J.L.; Jones, S.L.; Odle, J. Restoration of barrier function in injured intestinal mucosa. Physiol. Rev. 2007, 87, 545–564. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Suzuki, T.; Hara, H. Role of flavonoids in intestinal tight junction regulation. J. Nutr. Biochem. 2011, 22, 401–408. [Google Scholar] [CrossRef]
- Vogiatzoglou, A.; Mulligan, A.A.; Luben, R.N.; Lentjes, M.A.H.; Heiss, C.; Kelm, M.; Merx, M.W.; Spencer, J.P.E.; Schroeter, H.; Kuhnle, G.G.C. Assessment of the dietary intake of total flavan-3-ols, monomeric flavan-3-ols, proanthocyanidins and theaflavins in the European union. Br. J. Nutr. 2014, 111, 1463–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harnly, J.M.; Doherty, R.F.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Bhagwat, S.; Gebhardt, S. Flavonoid content of U.S. fruits, vegetables, and nuts. J. Agric. Food Chem. 2006, 54, 9966–9977. [Google Scholar] [CrossRef]
- Quine, S.D.; Raghu, P.S. Effects of (−)-epicatechin, a flavonoid on lipid peroxidation and antioxidants in streptozotocin-induced diabetic liver, kidney and heart. Pharmacol. Rep. 2005, 57, 610–615. [Google Scholar] [PubMed]
- Shariati, S.; Kalantar, H.; Pashmforoosh, M.; Mansouri, E.; Khodayar, M.J. Epicatechin protective effects on bleomycin-induced pulmonary oxidative stress and fibrosis in mice. Biomed. Pharmacother. 2019, 114, 108776. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Prieto, M.A.; Bettaieb, A.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (−)-epicatechin prevents TNFα-induced activation of signaling cascades involved in inflammation and insulin sensitivity in 3T3-L1 adipocytes. Arch. Biochem. Biophys. 2012, 527, 113–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, J.; Yu, Z.L.; Zhang, X.Y.; Li, W.Y.; Gao, D.N.; Wang, J.; Ma, X.C.; Nie, X.S.; Wang, W. Epicatechin alleviates inflammation in lipopolysaccharide-induced acute lung injury in mice by inhibiting the p38 MAPK signaling pathway. Int. Immunopharmacol. 2019, 66, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Xie, Y.N.; Xu, Y.L.; Hu, Z.H.; Wan, X.; Huang, H.C.; Huang, D.B. Protective effect of epicatechin on apap-induced acute liver injury of mice through anti-inflammation and apoptosis inhibition. Nat. Prod. Res. 2020, 34, 855–858. [Google Scholar] [CrossRef]
- Li, Y.; Ma, S.S.; Zhang, Y.T.; Yao, M.H.; Zhu, X.Z.; Guan, F.X. (−)-epicatechin mitigates radiation-induced intestinal injury and promotes intestinal regeneration via suppressing oxidative stress. Free Radic. Res. 2019, 53, 851–864. [Google Scholar] [CrossRef]
- Cremonini, E.; Wang, Z.W.; Bettaieb, A.; Adamo, A.M.; Daveri, E.; Mills, D.A.; Kalanetra, K.M.; Haj, F.G.; Karakas, S.; Oteiza, P.I. (−)-epicatechin protects the intestinal barrier from high fat diet-induced permeabilization: Implications for steatosis and insulin resistance. Redox Biol. 2018, 14, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.W.; Litterio, M.C.; Müller, M.; Vauzour, D.; Oteiza, P.I. (−)-epicatechin and NADPH oxidase inhibitors prevent bile acid-induced Caco-2 monolayer permeabilization through ERK1/2 modulation. Redox Biol. 2020, 28, 101360. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Yap, A.S.; Mullin, J.M.; Stevenson, B.R. Molecular analyses of tight junction physiology: Insights and paradoxes. J. Membr. Biol. 1998, 163, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mariscal, L.; Betanzos, A.; Nava, P.; Jaramillo, B.E. Tight junction proteins. Prog. Biophys. Mol. Biol. 2003, 81, 1–44. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef] [PubMed]
- Fukudome, I.; Kobayashi, M.; Dabanaka, K.; Maeda, H.; Okamoto, K.; Okabayashi, T.; Baba, R.; Kumagai, N.; Oba, K.; Fujita, M.; et al. Diamine oxidase as a marker of intestinal mucosal injury and the effect of soluble dietary fiber on gastrointestinal tract toxicity after intravenous 5-fluorouracil treatment in rats. Med. Mol. Morphol. 2014, 47, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.; Lindholt, J.S.; Erlandsen, E.J.; Mortensen, F.V. D-lactate as a marker of venous-induced intestinal ischemia: An experimental study in pigs. Int. J. Surg. 2011, 9, 428–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Biosci. 2009, 14, 2765–2778. [Google Scholar] [CrossRef] [Green Version]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta Biomembr. 2009, 1788, 864–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, D.M.; Ma, I.; Ma, T.Y. Molecular mechanism of tumor necrosis factor-α modulation of intestinal epithelial tight junction barrier. Am. J. Physiol. Gastroint. Liver Physiol. 2006, 290, G496–G504. [Google Scholar] [CrossRef] [Green Version]
- Al-Sadi, R.; Ye, D.M.; Said, H.M.; Ma, T.Y. IL-1β-induced increase in intestinal epithelial tight junction permeability is mediated by MEKK-1 activation of canonical NF-κB pathway. Am. J. Pathol. 2010, 177, 2310–2322. [Google Scholar] [CrossRef]
- Madsen, K.L.; Lewis, S.A.; Tavernini, M.M.; Hibbard, J.; Fedorak, R.N. Interleukin 10 prevents cytokine-induced disruption of T84 monolayer barrier integrity and limits chloride secretion. Gastroenterology 1997, 113, 151–159. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Intestinal redox biology and oxidative stress. Semin. Cell Dev. Biol. 2012, 23, 729–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadiiska, M.B.; Gladen, B.C.; Baird, D.D.; Germolec, D.; Graham, L.B.; Parker, C.E.; Nyska, A.; Wachsman, J.T.; Ames, B.N.; Basu, S.; et al. Biomarkers of oxidative stress study II: Are oxidation products of lipids, proteins, and DNA markers of CCl4 poisoning? Free Radic. Biol. Med. 2005, 38, 698–710. [Google Scholar] [CrossRef]
- Pirinccioglu, A.G.; Gökalp, D.; Pirinccioglu, M.; Kizil, G.; Kizil, M. Malondialdehyde (MDA) and protein carbonyl (PCO) levels as biomarkers of oxidative stress in subjects with familial hypercholesterolemia. Clin. Biochem. 2010, 43, 1220–1224. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Günther, C.; Neumann, H.; Neurath, M.F.; Becker, C. Apoptosis, necrosis and necroptosis: Cell death regulation in the intestinal epithelium. Gut 2013, 62, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Ghobrial, I.M.; Witzig, T.E.; Adjei, A.A. Targeting apoptosis pathways in cancer therapy. CA-Cancer J. Clin. 2005, 55, 178–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The Bcl-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Duprez, L.; Wirawan, E.; Berghe, T.V.; Vandenabeele, P. Major cell death pathways at a glance. Microbes Infect. 2009, 11, 1050–1062. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A. Targeting death and decoy receptors of the tumour-necrosis factor superfamily. Nat. Rev. Cancer 2002, 2, 420–430. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Riedl, S.J.; Shi, Y.G. Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol. 2004, 5, 897–907. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items § | Treatment Groups ‡ | p-Value | ||
|---|---|---|---|---|
| CON | LEPI | HEPI | ||
| DAO, U/L | 9.04 ± 0.25 a | 7.48 ± 0.14 b | 7.27 ± 0.16 b | <0.001 |
| D-Lactic acid, ng/mL | 563.64 ± 8.17 a | 488.49 ± 5.73 b | 507.46 ± 4.32 b | <0.001 |
| Items § | Treatment Groups ‡ | p-Value | ||
|---|---|---|---|---|
| CON | LEPI | HEPI | ||
| Duodenum | ||||
| SOD, U/mg protein | 70.90 ± 2.89 | 81.74 ± 3.50 | 79.20 ± 4.66 | 0.127 |
| CAT, U/mg protein | 46.56 ± 2.48 c | 66.33 ± 2.88 b | 78.54 ± 2.79 a | <0.001 |
| GSH-PX, U/mg protein | 184.44 ± 4.55 b | 201.16 ± 4.75 b | 241.52 ± 7.83 a | <0.001 |
| MDA, nmol/mg protein | 2.97 ± 0.18 a | 1.66 ± 0.09 b | 1.47 ± 0.10 b | <0.001 |
| Jejunum | ||||
| SOD, U/mg protein | 50.14 ± 1.88 | 53.99 ± 2.62 | 51.10 ± 2.15 | 0.461 |
| CAT, U/mg protein | 23.26 ± 0.91 b | 29.07 ± 1.85 a | 30.65 ± 1.58 a | 0.006 |
| GSH-PX, U/mg protein | 170.46 ± 4.85 b | 180.83 ± 3.37 a, b | 185.56 ± 3.79 a | 0.044 |
| MDA, nmol/mg protein | 1.97 ± 0.08 a | 1.53 ± 0.07 b | 1.38 ± 0.04 b | <0.001 |
| Ileum | ||||
| SOD, U/mg protein | 54.23 ± 2.76 b | 71.78 ± 2.24 a | 80.04 ± 2.60 a | <0.001 |
| CAT, U/mg protein | 29.24 ± 1.29 | 30.53 ± 1.50 | 33.89 ± 1.88 | 0.123 |
| GSH-PX, U/mg protein | 127.92 ± 3.85 | 138.81 ± 5.06 | 141.77 ± 4.47 | 0.095 |
| MDA, nmol/mg protein | 2.69 ± 0.09 a | 2.11 ± 0.10 b | 2.01 ± 0.07 b | <0.001 |
| Items § | Treatment Groups ‡ | p-Value | ||
|---|---|---|---|---|
| CON | LEPI | HEPI | ||
| Duodenum, pg/mg protein | ||||
| IL-1β | 30.85 ± 1.38 | 32.75 ± 1.23 | 28.47 ± 1.13 | 0.075 |
| IL-10 | 486.92 ± 12.63 | 517.91 ± 16.42 | 497.73 ± 12.89 | 0.308 |
| TNF-α | 309.87 ±7.39 a | 230.81 ± 5.28 b | 212.71 ± 9.55 b | <0.001 |
| IFN-γ | 338.01 ± 9.43 | 324.94 ± 7.32 | 319.80 ± 8.59 | 0.315 |
| Jejunum, pg/mg protein | ||||
| IL-1β | 32.68 ± 1.18 | 30.68 ± 0.80 | 30.31 ± 0.85 | 0.193 |
| IL-10 | 440.39 ± 11.10 b | 535.18 ± 8.24 a | 466.93 ± 14.38 b | <0.001 |
| TNF-α | 325.84 ± 7.47 a | 301.07 ± 6.86 b | 297.19 ± 6.17 b | 0.015 |
| IFN-γ | 310.94 ± 6.12 | 301.32 ± 7.30 | 294.26 ± 6.78 | 0.238 |
| Ileum, pg/mg protein | ||||
| IL-1β | 27.69 ± 1.32 a | 22.74 ± 0.57 b | 25.46 ± 1.12 a,b | 0.011 |
| IL-10 | 351.85 ± 12.71 | 353.51 ± 10.48 | 380.03 ± 9.19 | 0.147 |
| TNF-α | 231.54 ± 6.88 a | 203.52 ± 5.97 b | 194.74 ± 9.31 b | 0.006 |
| IFN-γ | 240.08 ± 11.15 | 225.44 ± 8.35 | 234.89 ± 5.88 | 0.497 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, J.; Zhang, L.; Ruan, Z. Dietary Supplementation with Epicatechin Improves Intestinal Barrier Integrity in Mice. Foods 2022, 11, 3301. https://doi.org/10.3390/foods11203301
Wan J, Zhang L, Ruan Z. Dietary Supplementation with Epicatechin Improves Intestinal Barrier Integrity in Mice. Foods. 2022; 11(20):3301. https://doi.org/10.3390/foods11203301
Chicago/Turabian StyleWan, Jin, Li Zhang, and Zheng Ruan. 2022. "Dietary Supplementation with Epicatechin Improves Intestinal Barrier Integrity in Mice" Foods 11, no. 20: 3301. https://doi.org/10.3390/foods11203301
APA StyleWan, J., Zhang, L., & Ruan, Z. (2022). Dietary Supplementation with Epicatechin Improves Intestinal Barrier Integrity in Mice. Foods, 11(20), 3301. https://doi.org/10.3390/foods11203301