Abstract
5-hydroxytryptophan (5-HTP) is an important substance thought to improve depression. It has been shown that Lactobacillus can promote the secretion of 5-HTP in the body and thus ameliorate depression-like behavior in mice. However, the mechanism by which Lactobacillus promotes the secretion of 5-HTP is unclear. In this study, we investigated the promoting effect and mechanism of Lactobacillus, isolated from Chinese fermented foods, on the secretion of 5-HTP. The results showed that Lactobacillus (L.) pentosus LPQ1 exhibited the strongest 5-HTP secretion-promoting effect ((9.44 ± 0.69)-fold), which was dependent on the mixture of compounds secreted by L. pentosus LPQ1 (termed SLPQ1). In addition, the results of the RNA sequencing (RNA-seq) and quantitative real-time polymerase chain reaction (qRT-PCR) analyses indicated that SLPQ1 alters the TNF and oxidative phosphorylation signaling pathways. Moreover, the SLPQ1 ultrafiltration fraction (>10 kDa) showed a similar 5-HTP promoting effect as SLPQ1. Furthermore, reverse-phase liquid chromatography-tandem mass spectrometry (RPLC-MS/MS) identified 29 compounds of >10 kDa in SLPQ1, including DUF488 domain-containing protein, BspA family leucine-rich repeat surface protein, and 30S ribosomal protein S5, which together accounted for up to 62.51%. This study reports new findings on the mechanism by which L. pentosus LPQ1 promotes 5-HTP production in some cell lines in vitro.
1. Introduction
Depression is a major contributor to disability and also imposes a significant economic and health care burden on patients, their families, and the health system [1,2]. Experts predict that by 2030, the cost of curing mental illnesses, including depression, will be as high as $6 trillion [3]. Antidepressants, such as the selective 5-hydroxytryptamine (5-HT), also known as serotonin reuptake inhibitor fluoxetine, have antidepressant effects by blocking 5-HT reuptake [4]. However, patients take antidepressant medications for a long time before they relieve depression, and they are often accompanied by adverse side effects [5,6]. Food-derived substances without side effects have been reported to alleviate depression by improving neurotransmitter synthesis, modulating the activation of the brain-derived neurotrophic factor tropomyosin receptor kinase B (BDNF-TrκB) pathway, inhibiting the expression of inflammatory genes, and reshaping the intestinal flora of depressed mice [7,8]. Therefore, safe food-derived substances are thought to have the potential to improve depression.
Insufficient levels of synaptic 5-HT in the brain are considered to be a manifestation of the classical pathogenesis of depression [9]. The neurotransmitter 5-HT ameliorates depression mainly by acting on mood-related nervous systems [10]. However, 5-HT cannot cross the blood-brain barrier (BBB) and is synthesized locally in the brain from its precursors [11]. Both tryptophan and 5-hydroxytryptophan (5-HTP) are used in the biosynthesis of 5-HT. Roughly 90% of dietary tryptophan is used in the kynurenine pathway, a process that competes with the synthetic pathway of 5-HT [12,13]. Unlike tryptophan, 5-HTP freely crosses the BBB and increases the 5-HT levels in the brain to exert an antidepressant effect [14,15,16]. In 30 patients with their first depressive episode treated with 5-HTP for two weeks, 5-HTP showed some antidepressant effect in 73.33% of the patients [17]. Similarly, 40 out of 59 patients with depressive symptoms who were given 150–300 mg of 5-HTP daily for three weeks showed significant improvement in depressive symptoms [18]. However, oral 5-HTP has been reported to cause adverse side effects, such as abnormal psychological functioning, behavioral disturbances and intolerance [15].
Lactic acid bacteria (LAB) derived from fermented foods are considered natural and safe [19]. Accordingly, the use of probiotics for the alleviation of neurodegenerative diseases, including depression, have gained attention in recent years [20]. The relative abundance of Lactobacillus was reported to be closely related to the serum level of 5-HTP [21]. Lactococcus lactis WHH2078 was also found to modulate 5-HT metabolism by increasing colonic and serum 5-HTP levels, thus exerting antidepressant effects in mice [22]. Similarly, Bifidobacterium exhibiting 5-HTP synthesis modulating effects have been found to alleviate depressive symptoms and associated microbiota dysbiosis in mice [23]. However, the mechanisms by which LAB enhance 5-HTP secretion have not been reported.
In this study, we evaluated the promotion of 5-HTP secretion in RIN-14B cells by ten LAB strains from traditional Chinese fermented foods and investigated the active components and mechanism of action of the LAB strain with the highest 5-HTP-promoting effect. To the best of our knowledge, this is the first report of the mechanism by which active compounds secreted by LAB strains regulate the secretion of 5-HTP in RIN-14B cells.
2. Materials and Methods
2.1. Materials and Reagents
Lactobacillus (L.) pentosus LPQ1 and L. helveticus Q13 were from Qula, a raw yak milk cheese (Qinghai, China). L. plantarum M45, L. casei 96, L. plantarum 05015, L. plantarum Q11, and L. plantarum Q9 were from traditional fermented pickles (Chongqing, China). L. rhamnosus T53 and L. helveticus Y4 were from traditional fermented yogurt (Qinghai, China). L. plantarum 05007 was from traditional fermented yogurt (Chengdu, China). Hanks’ balanced salt solution (HBSS) and RNase-free water were obtained from Beyotime Biotechnology Inc. (Shanghai, China). Fetal bovine serum (FBS) was supplied by Biological Industries Beit HaEmek Ltd. (Beit HaEmek, Israel). TRIzol reagent was obtained from Thermo Fisher Scientific Inc. (Waltham, MA, USA). RPMI1640 medium was purchased from Beijing Solarbio Technology Co., Ltd. (Beijing, China).
2.2. RIN-14B Cell Culture
RIN-14B cells were provided by American Typical Culture Collection (ATCC, Manassas, VA, USA). RIN-14B cells were grown in RPMI 1640 with 10% FBS, 1% penicillin-streptomycin solution, in an incubator at 37 °C with a humidified atmosphere and 5% CO2. After reaching confluence of approximately 80%, cells were detached by digestion with trypsin and passaged. The culture medium was changed every 2–3 days. RIN-14B cells from the 10th to 30th generation were used in this study.
2.3. The Preparation of LAB Strains
The preparation of LAB was performed according to a previous study [19] with slight modifications. The LAB strains were inoculated (2%, v/v) in de Man-Rogosa-Sharpe (MRS) medium and then incubated at 37 °C for 18 h. After centrifuging at 4000× g for 10 min at 4 °C, the bacteria were collected, and then the number of bacterial cells was adjusted to 1 × 109 CFU/mL.
2.4. Preparation of the Mixture of Compounds Secreted by L. pentosus LPQ1 (SLPQ1)
L. pentosus LPQ1 at 1 × 109 CFU/mL was incubated in an HBSS solution for 30 min at 37 °C and centrifuged at 4000× g for 10 min at 4 °C. Afterwards, the supernatant was collected and then filtered through a 0.22-μL filter to obtain the mixture of secreted compounds. The mixture of compounds secreted by L. pentosus LPQ1 (termed SLPQ1) was lyophilized and stored at −80 °C for subsequent studies.
2.5. Evaluation of the Promoting Effect of LAB and Its Secreted Compounds on the Secretion of 5-HTP by RIN-14B Cells
As described previously, we performed experiments on the amount of 5-HTP secretion [24,25]. RIN-14B cells were inoculated into 24-well plates at a density of 4 × 105 cells/mL and incubated for 72 h. Then, after washing the cells with 1 mL of HBSS buffer with 0.1% bovine serum albumin (BSA) and 2 M fluoxetine, 1 mL of an HBSS suspension containing LAB or their secreted compounds was added to the experimental group, while only the HBSS solution without LAB or their secretions was added to the control group. After incubation for 30 min at 37 °C, the supernatants were collected and centrifuged to remove the precipitate.
2.6. Determination of 5-hydroxytryptophan Content
The test method for the measurement of the 5-HTP content was based on a previously reported method [24]. In brief, the cell supernatant was passed through a 0.22-μM filter. The 5-HTP content of the cell supernatant was assayed using a HPLC system (Shimadzu Corporation, Kyoto, Japan) equipped with a fluorescence detector (RF-20A, Shimadzu). The 20 μL of sample was loaded on an Agilent Zorbax XDB-C18 column (250 mm × 4.6 mm, 5 μm), and eluted with 0.1 M sodium acetate buffer (containing 0.1 mM EDTA-2Na) and methanol in a ratio of 85:15 (v/v). The column temperature was 30 °C, and the flow rate was 1.0 mL/min. The fluorescence intensity was detected at wavelengths of 330 nm (emission) and 290 nm (excitation).
2.7. Methyl Thiazolyl Tetrazolium (MTT) Assay
The MTT assay was performed as described previously [26], with slight modifications. RIN-14B cells were seeded into 96-well plates at a density of 4 × 105 cells/mL and grown for 72 h. Subsequently, after gently aspirating the medium and washing the cells once with HBSS, 100 μL of sample solution was added to each well and the plate was incubated for 30 min. Afterwards, 10 μL of MTT reagent was added to each well, and the plate was protected from light and incubated at 37 °C for 4 h. Then, after removing the supernatant from all wells, 100 L of dimethyl sulfoxide (DMSO) was added to each well, and after agitating the mixture for 15 min the absorbance (Abs) of each well was measured at 490 nm. The cell viability was estimated using the formula:
Cell viability (%) = (Abs sample − Abs blank) × 100/(Abs control − Abs blank).
2.8. RNA-Seq Analysis
RIN-14B cells were inoculated into 24-well plates at a density of 4 × 105 cells/mL and then incubated for 72 h. Subsequently, after discarding the medium and washing the cells with 1 mL of HBSS buffer containing 0.1% BSA and 2 μM fluoxetine, 1 mL of HBSS suspension containing 16 mg/mL of SLPQ1 was added to each well (the control group received HBSS solution without SLPQ1). After incubation at 37 °C for 30 min, the supernatant was gently aspirated and discarded. Then, after washing the attached cells with pre-cooled HBSS, the cells were lysed by adding 1 mL of TRIzol reagent, and the cell lysate was transferred into sterile enzyme-free lyophilization tubes, snap-frozen in liquid nitrogen and stored at −80 °C for subsequent RNA extractions. The samples were subjected to RNA-seq analysis using a previously described procedure [27]. In brief, total cellular RNA was extracted using TRIzol (Thermo Fisher Scientific, Waltham, MA, USA). The purity and concentration of the extracted RNA were assayed using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific), and its integrity was assessed by agarose gel electrophoresis. Afterwards, the mRNA was enriched from total RNA using oligo dT affinity beads and then randomly split into 300 bp fragments. These fragments were used as templates for reverse transcription and the production of double-stranded cDNA. A single nucleotide A and a connector were connected to the end-repaired double-stranded cDNA, following PCR amplification for the de novo RNA-seq cDNA library. Sequencing was performed using the NovaSeq 6000 System (Illumina, Inc., San Diego, CA, USA), and the bioinformatics analysis was performed using the free online platform of MajorBio Cloud Platform (https://cloud.majorbio.com, accessed on 18 October 2022).
2.9. qRT-PCR Analysis
Total RNA was extracted using TRIzol reagent, and subsequently its concentration and purity were determined. Then, total RNA was reverse transcribed into cDNA using a cDNA reverse transcription kit (Thermo Fisher Scientific Inc.) and stored at −80 °C for subsequent analysis. PCR reaction conditions were as follows: pre-denaturation at 95 °C for 10 min; followed by 40 cycles at 95 °C, 15 s; 60 °C, 1 min; 72 °C, 30 s. The 2−ΔΔCT method was used to calculate the expression of the target gene primer sequences, which are shown in Table S1.
2.10. Analysis of Active Compounds
Appropriate amounts of samples were separated using 10 kDa, 3 kDa and 1 kDa ultrafiltration tubes. The recovered fractions were then lyophilized and kept at −80 °C for further studies. The reverse-phase liquid chromatography-tandem mass spectrometry (RPLC-MS/MS) analysis of the fraction of >10 kDa was carried out according to a previously reported method [28], with some modifications. Briefly, the fraction of >10 kDa was denatured with 8 M urea, reduced with 10 mM iodoacetamide, and then alkylated with 20 mM iodoacetamide. Subsequently, the trypsin (Promega, WI, USA) at an enzyme/substrate ratio of 1:50 (w/w) was added for digestion at 37 °C for 18 h. The digested sample was desalted with C18 ZipTips microcolumns (Millipore, Burlington, MA, USA) and then lyophilized for further RPLC-MS/MS analysis.
The lyophilized sample was in 0.1% (v/v) formic acid and was analyzed by a QE-HF mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA) coupled to an Easy-nLC 1000 HPLC system (Thermo Fisher Scientific). The sample was loaded onto a C18 reversed-phase analytical column (Acclaim PepMap RSLC, 75 µm × 15 cm, nanoViper, 2 µm 100 Å C18 particles; Thermo Fisher Scientific) and eluted with a linear gradient of 0.1% (v/v) formic acid and 0.1% (v/v) formic acid in 80% (v/v) acetonitrile for 70 min at a flow rate of 400 nL/min. The MS1 scan was acquired in the mass range of 350–1550 m/z at a resolution of 120,000, with an automatic gain control (AGC) target value of 4 × 105 and a maximum injection time (IT) of 50 ms. The MS2 scan was performed at a resolution of 30,000 with an AGC target value of 1e5, a maximum IT of 100 ms, and a normalized collision energy of 32 eV. The raw MS files were analyzed using PEAKS Studio v7.5 software (Bioinformatic Solutions Inc, waterloo, ON, Canada).
2.11. Statistical Analysis
The data from each test, performed at least three times, were expressed as the mean standard deviation. All data were evaluated for statistically significant differences (p < 0.05) by one-way analysis of variance (ANOVA) with Duncan’s test using the IBM SPSS 23 software (IBM Corporation, Armonk, NY, USA).
3. Results
3.1. Effect of LAB Strains on 5-HTP Secretion
As shown in Figure 1A, among ten LAB strains, the strains LPQ1 and 05015 exhibited a strong 5-HTP production and secretion effect on RIN-14B cells. However, as shown in Figure 1B, the 05015 strain was somewhat toxic to the cells; for this reason, the LPQ1 strain was used as a potential strain to increase the secretion of 5-HTP in subsequent experiments.
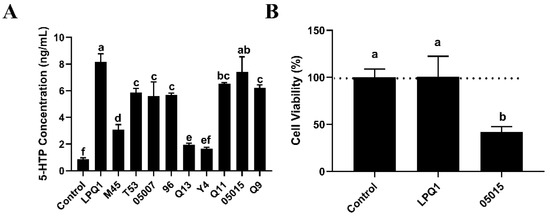
Figure 1.
Effects of different lactic acid bacteria (LAB) strains on the secretion of 5-hydroxytryptophan (5-HTP) by RIN-14B cells. (A) Effects of LAB strains on the secretion of 5-HTP by RIN-14B cells. (B) Cell viability. Control: RIN-14B cells; LPQ1: RIN-14B cells with Lactobacillus (L.) pentosus LPQ1; M45: RIN-14B cells with L. plantarum M45; T53: RIN-14B cells with L. rhamnosus T53; 05007: RIN-14B cells with L. plantarum 05007; 96: RIN-14B cells with L. casei 96; Q13: RIN-14B cells with L. helveticus Q13; Y4: RIN-14B cells with L. helveticus Y4; Q11: RIN-14B cells with L. plantarum Q11; 05015: RIN-14B cells with L. plantarum 05015; Q9: RIN-14B cells with L. plantarum Q9. Data are expressed as the mean ± standard deviation (n = 3). Different letters (a–f) indicate significant differences (p < 0.05).
3.2. Effect of SLPQ1 on 5-HTP Secretion by RIN-14B Cells
There was no significant difference between the amount of 5-HTP secreted by RIN-14B cells treated with L. pentosus LPQ1 and those treated with its secreted compounds SLPQ1, as shown in Figure 2A (p > 0.05). Therefore, the compound that affects the production and secretion of 5-HTP by RIN-14B cells may be a component of the SLPQ1 mixture. Indeed, SLPQ1 promoted 5-HTP secretion from RIN-14B cells in a concentration-dependent manner (Figure 2B). However, while 16 mg/mL SLPQ1 was the optimal concentration, SLPQ1 concentrations greater than 16 mg/mL had significant toxic effects on the cells (Figure 2C). These results suggested that the active ingredient of L. pentosus LPQ1 is among its secreted compounds.
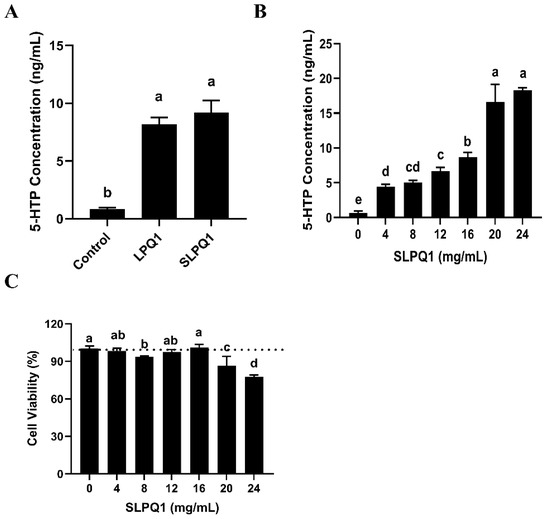
Figure 2.
Effect of active ingredients of the mixture of secreted compounds, SLPQ1, collected from Lactobacillus (L.) pentosus LPQ1 on the secretion of 5-hydroxytryptophan (5-HTP). (A) Identification of effective compounds. (B) Determination of the amount of SLPQ1 used. (C) Effect of different SLPQ1 amounts on cell activity. Control: control group; LPQ1: treatment group after L. pentosus LPQ1; LPQ1-S: treatment group after secretion of L. pentosus LPQ1 without L. pentosus LPQ1 (no freeze-drying); SLPQ1: treatment group after SLPQ1. Data are expressed as the mean ± standard deviation (n = 3). Different letters (a–e) indicate significant differences (p < 0.05).
3.3. Analysis of the Transcriptional Changes
We performed RNA-seq analysis on RIN-14B cells treated with SLPQ1 in order to understand the SLPQ1-mediated molecular regulation of 5-HTP production and secretion by RIN-14B cells. The total number of raw sequencing data entries generated per sample exceeded 60,000,000, with Q20 and Q30 exceeding 98 and 94%, respectively, as shown in Table S2.
Transcripts Per Million reads (TMPs) sequentially normalizes gene length and sequencing depth, a process that allows for consistent total expression between samples and thus comparison of expression between genes. Using the Pearson correlation coefficient, a measure of the linear correlation between two variables, the closer the R value is to 1, the stronger the correlation between the samples. Intra-group correlations were high for the control and SLPQ1 groups, but the inter-group correlations were low (Figure 3A). Principal component analysis (PCA) clustered the samples according to their expression level (high or low), and the higher the similarity of the samples, the closer the distribution of sample points. The control group and SLPQ1 group were closer within each group and further apart between the groups (Figure 3B). The Veen analysis can identify the number of genes co-expressed and specifically expressed between different groups. There are 295 genes specifically expressed in the control group and 165 genes specifically expressed in the SLPQ1 group, and a total of 12,829 genes expressed in both groups (Figure 3C). The volcano plot allowed visualization of upregulated or downregulated genes. As shown in Figure 3D, there were 485 changed genes, including 169 upregulated genes and 316 downregulated genes. These findings support the notion that SLPQ1 significantly affects functions of RIN-14B cells.
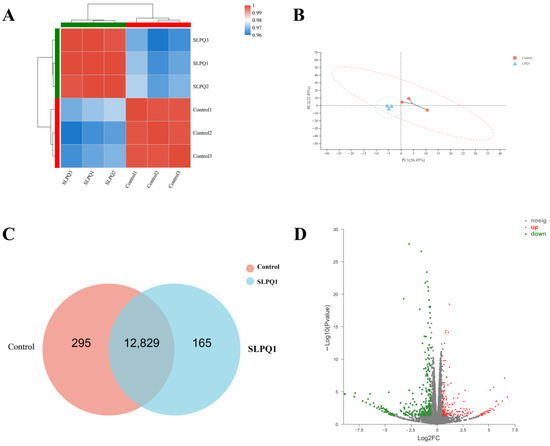
Figure 3.
Effects of the mixture of secreted compounds, SLPQ1, collected from Lactobacillus pentosus LPQ1 on the global gene expression profiling in RIN-14B cells. (A) Sample correlation heatmap. (B) Principal component analysis (PCA). (C) Veen diagram. (D) Volcano plot. Control: control group; SLPQ1: treatment group after SLPQ1. Control: control group; SLPQ1: treatment group after SLPQ1.
3.4. Differentially Expressed Genes Analysis by Gene Ontology Term and Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Analyses
The Gene Ontology (GO) database allows the qualification and description of genes, which are classified into three categories: biological process (BP), molecular function (MF) and cellular composition (CC) [19]. The top 30 enriched GO terms are shown in Figure 4A.
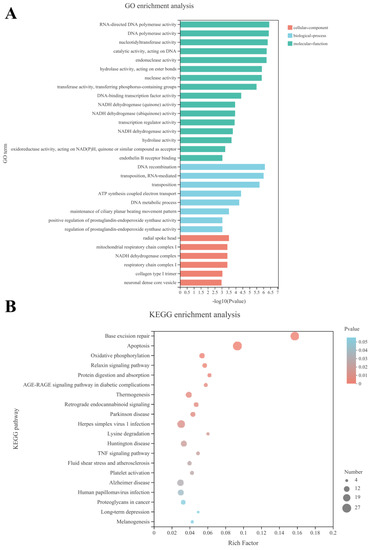
Figure 4.
Effects of the mixture of secreted compounds, SLPQ1, collected from Lactobacillus pentosus LPQ1 on Gene Ontology (GO) term enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis. (A) GO term enrichment analysis. (B) KEGG pathway enrichment analysis. Control: control group; SLPQ1: treatment group after SLPQ1.
Significant functional differences between the SLPQ1 and Control groups were found in the following GO CC categories enriched with differentially expressed genes (DEGs): radial spoke head, mitochondrial respiratory chain complex I, NADH dehydrogenase complex, etc., as shown in Figure 4A. Significant functional differences between the SLPQ1 and Control groups were found in the following GO BP categories enriched with DEGs: DNA recombination, transposition and RNA-mediated, transposition, etc., as shown in Figure 4A. Additionally, RNA-directed DNA polymerase activity, DNA polymerase activity, and nucleotidyltransferase activity, etc., are among a few of the noteworthy functional changes between the SLPQ1 and Control groups in the GO MF enriched with DEGs (Figure 4A).
The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis revealed the signaling pathways significantly enriched with DEGs/transcripts. The top 20 significantly enriched pathways identified by KEGG pathway enrichment analysis are shown in Figure 4B and include the signaling pathways involved in neurodegenerative pathologies such as Parkinson’s disease, Retrograde endocannabinoid signaling, Lysine degradation, Huntington disease, Alzheimer disease, Long-term depression, as well as Apoptosis, Oxidative phosphorylation, and TNF signaling pathways, which are involved in the regulation of cellular functions and are closely related to each other. Our experimental results suggest that the above-mentioned signaling pathways are the most likely potential mechanisms by which SLPQ1 promotes 5-HTP secretion in RIN-14B cells.
3.5. Effect of SLPQ1 on Oxidative Phosphorylation- and TNF Signaling Pathway-Related Gene Expression
TNF is a key mediator of inflammation, and inflammation is one of the main pathological features of depression [29,30]. Accordingly, regulation of TNF levels is considered to be important in the treatment of various Central Nervous System (CNS)-related diseases [31]. In addition, alleviation of oxidative phosphorylation was reported to improve neurological functions in mice [32]. RNA-seq analysis revealed that the Atf4 gene was significantly downregulated, while End1, Lif and Socs3 genes were significantly upregulated in the TNF signaling pathway in the SLPQ1 group compared with the control group. The results of the qRT-PCR expression analysis of Atf4, End1, Lif and Socs3 mRNAs were consistent with the RNA-seq results, as shown in Figure 5A. Among them, the Socs3 mRNA expression level was upregulated by 3.65-fold, and Lif and End1 mRNA expression levels were upregulated by 1.91- and 1.79-fold, respectively, whereas that of Atf4 mRNA was downregulated by 0.33-fold (Figure 5B).
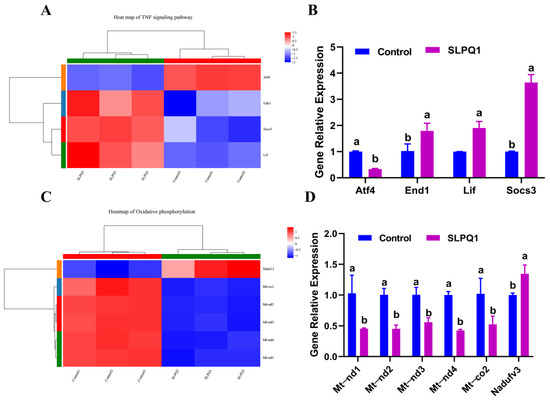
Figure 5.
Effects of the mixture of secreted compounds, SLPQ1, collected from Lactobacillus pentosus LPQ1 on the genes related to the TNF and oxidative phosphorylation signaling pathways. (A) Heatmap of the TNF signaling pathway. (B) Effect of SLPQ1 on the expression of the TNF signaling pathway-related genes in RIN-14B cells. (C) Heatmap of the oxidative phosphorylation signaling pathway. (D) Effect of SLPQ1 on the expression of the oxidative phosphorylation-related genes in RIN-14B cells. Control: control group; SLPQ1: treatment group after SLPQ1. Data are expressed as the mean ± standard deviation (n = 3). For the same gene, two different letters (a, b) represent significant differences (p < 0.05).
The RNA-seq analysis also revealed significant upregulation of Ndufv3 mRNA expression and significant downregulation of mt-nd1, mt-nd2, mt-nd3, mt-nd4 and mt-co2 mRNA expression in the oxidative phosphorylation pathway (Figure 5C). The results of the qRT-PCR expression analysis were similar to those detected by RNA-seq (Figure 5D). Compared with the control group, the Ndufv3 mRNA expression level was upregulated 1.35-fold, and mt-nd1, mt-nd2, mt-nd3, mt-nd4 and mt-co2 mRNA expression levels were downregulated 0.45-, 0.45-, 0.56-, 0.42- and 0.52-fold, respectively (Figure 5D). These results indicate that the expression levels obtained by RNA-seq and qRT-PCR analyses and the upregulation and downregulation of these genes are fairly close.
3.6. Effects of the Active Compounds Present in SLPQ1 on 5-HTP Secretion
Samples were separated using ultrafiltration tubes, which through centrifugal force retain compounds of different molecular weights present in the sample. The larger the molecular weight of the compounds in the fraction of the samples, the stronger the promotion of 5-HTP production in RIN-14B cells. The highest secretion of 5-HTP was observed with fractions of SLPQ1 > 10 kDa, followed by fractions of 3–10 kDa. The results obtained clearly show that the promotion of 5-HTP secretion by SLPQ1 in RIN-14B cells is achieved mainly through fractions > 10 kDa (Figure 6).
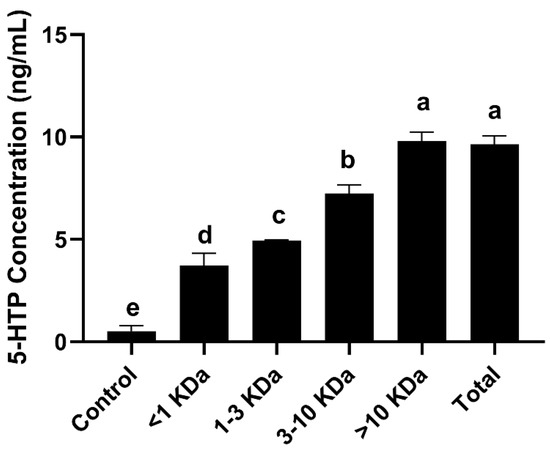
Figure 6.
Effects of the mixture of secreted compounds, SLPQ1, collected from Lactobacillus pentosus LPQ1 of different molecular weights on the promotion of 5-HTP secretion. Control: control group; SLPQ1: treatment group after SLPQ1. Data are expressed as the mean ± standard deviation (n = 3). Different letters indicate significant differences (p < 0.05).
Furthermore, the compounds larger than 10 kDa in SLPQ1 were identified using RPLC-MS/MS. As shown in Table 1, 29 proteins were found in the fractions of the samples > 10 kDa. The top five groups of compounds with relatively high contents were DUF488 structural domain protein, BspA family leucine-rich repeat surface protein, 30S ribosomal protein S5, polysaccharide biosynthesis protein and phage terminal enzyme small subunit P27 family.

Table 1.
Identification results of compounds larger than 10 kDa in the SLPQ1 collected from Lactobacillus pentosus LPQ1.
4. Discussion
The 5-HTP is thought to play a role in mood, behavior and sleep, and is a precursor of 5-HT or serotonin, the neurotransmitter used in the treatment of depression [33,34]. Studies have found that LAB promoting 5-HTP secretion have good depression-relieving properties [9,22,23]. In addition, depression-like behavior was reported to be improved by gavage-administered 5-HTP in mice through the regulation of intestinal flora [35]. In the present study, all LAB strains were found to promote 5-HTP secretion, and it is worth noting that L. pentosus LPQ1 showed the highest promotion of 5-HTP secretion, reaching (9.44 ± 0.69)-fold. Remarkably, we found that SLPQ1 contained the compound through which L. pentosus LPQ1 exerted its 5-HTP secretion-promoting effect. The SLPQ1 concentration of 16 mg/mL was non-toxic to the cells, and the secretion of 5-HTP could reach 13.90 ± 1.06-fold.
High-throughput sequencing technology is used in RNA-seq analysis to determine the sequence of mRNA, short RNA, and non-coding RNA, which can reflect the expression level of each gene, and RNA-seq analysis is widely used as a molecular biology method to study the molecular mechanism of biological processes in treated samples [36]. This study was based on RNA-seq analysis of gene expression changes after SLPQ1 treatment in RIN-14B cells. Veen plot analysis, correlation heat map analysis and PCA plot analysis revealed that gene expression in RIN-14B cells was altered after SLPQ1 treatment, with significant inter-group differences and good intra-group reproducibility. Volcano plot analysis showed a significant effect of SLPQ1 on RIN-14B cells, with 169 upregulated and 316 downregulated genes. Cellular gene expression was changed by the effect of SLPQ1 on RIN-14B cells. The GO term enrichment analysis and KEGG pathway enrichment analysis can be used to identify cell biological functions and signaling pathways based on the DEGs identified by RNA-seq analysis, respectively. In this study, we found that the GO BP categories enriched with DEGs are mainly involved in bioregulation and metabolic processes, the GO MF categories enriched with DEGs are mainly involved in binding, catalytic activity and translation regulatory activity, and the GO CC categories enriched with DEGs are mainly associated with cellular components, organelle components and composition.
Chronic low-grade inflammation plays an important role in the pathophysiology of major depressive disorder [37]. The elevated level of serum TNF-α is considered to be one of risk assessment indicators for depression [38]. Although there are reports claiming anti-TNF-α therapy and the improvement of mood symptoms are irrelevant [39,40], some studies have proved the antidepressant effects of selective TNF-α antagonists, such as infliximab and etanercept, in non-psychiatric patients, which have been summarized in a critical review by Sarit Uzzan et al. [41]. Moreover, many animal studies have reported that TNF and 5-HT levels were negatively correlated in depressed mice, and Lactobacillus helveticus NS8 and Bifidobacterium breve CCFM1025 alleviated depression-like behavior in mice by decreasing TNF level with elevated 5-HT level [24,42]. Remarkably, excessive and prolonged stress may lead to excessive activation of ATF4, thereby inducing expression of apoptotic proteins and causing neuronal death [43]. It has also been found that increased expression of SOCS3 promotes M2 polarization in microglia, which exerts anti-inflammatory and neuroprotective effects [44]. Similarly, End-1, which is widely distributed throughout the central nervous system, is believed to control hormone and neurotransmitter release [45]. In addition, Yes-related proteins can affect astrocyte maturation and differentiation by regulating endothelial LIF secretion, which has been shown to be closely associated with episodes of major depression [46]. Interestingly, it has been shown that oxidative phosphorylation can be considered as a common pathogenesis of systemic inflammatory diseases such as ankylosing spondylitis and neurodegenerative pathologies such as dementia [47]. In this study, we found that SLPQ1 significantly elevated the expression levels of End1, Lif, Socs3 and Ndufv3 mRNA, and significantly downregulated the expression levels of Atf4, Mt-nd1, Mt-nd2, Mt-nd3 mRNA. These findings demonstrated that the promotion of 5-HTP secretion by L. pentosus LPQ1 was associated with the expression of genes involved in the TNF and oxidative phosphorylation pathways.
Ultrafiltration is a widely used and practical technique for separating bioactive compounds [19]. Over the separation range, there was a positive correlation between the molecular weight of the SLPQ1 fraction and 5-HTP secretion. The effect of fractions larger than 10 kDa on 5-HTP secretion was not significantly different from the original solution (p > 0.05). It is slightly different from the study indicating that the 5-HTP secretion promoting compounds in RIN-14B cells are short-chain fatty acids [24]. In this study, macromolecules larger than 10 kDa in SLPQ1 and a total of 29 proteins were identified using RP-MS/MS. Zinc metalloproteinases belong to the family of matrix metalloproteinases (MMPs), which have been shown to be significantly involved in both the physiological and pathological aspects of depression, besides being important mediators of the immune response [48]. Furthermore, it has been shown that recombinant MMP-9 restores behavioral deficits manifested in learning memory in mice [49]. Flavodoxin is thought to cope with oxidative challenges, and elevated levels of oxidative stress are thought to be associated with depression [50,51]. Therefore, it can be concluded that among the 29 SLPQ1 molecules larger than 10 kDa, zinc metalloprotease (4.151%) and flavodoxin (0.004%) may be among the factors through which SLPQ1 exerts its enhancing effect on 5-HTP secretion.
5. Conclusions
In summary, we isolated the strain L. pentosus LPQ1 from the traditional Chinese fermented food Qula. L. pentosus LPQ1 and its secreted compounds, SLPQ1, significantly promoted the production and secretion of 5-HTP in RIN-14B cells. The promotion of 5-HTP secretion by SLPQ1 was associated with the regulation of the expression of genes involved in TNF and oxidative phosphorylation signaling pathway. Compounds larger than 10 kDa in SLPQ1 (mainly DUF488 domain-containing protein, BspA family leucine-rich repeat surface protein, and 30S ribosomal protein S5) played a key role in promoting 5-HTP secretion. The findings of this study shed light on the mechanism through which LAB strains induces 5-HTP secretion.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/foods11233895/s1. Table S1: Primer sequences. Table S2: Quality of the RNA-sequence data of each sample. SLPQ1, treatment group after SLPQ1 interacted with RIN-14B cells; Control, control group.
Author Contributions
Conceptualization, J.S. and H.S.; methodology, Y.Z. (Yixiu Zeng) and J.S.; software, Y.Z. (Yuhong Zhang) and F.Z.; validation, Y.Z. (Yixiu Zeng) and J.S.; formal analysis, Y.H.; investigation, Y.Z. (Yixiu Zeng); resources, H.S.; data analysis, Y.Z. (Yixiu Zeng); writing—original draft, Y.Z. (Yixiu Zeng) and J.S.; writing—review and editing, Y.Z. (Yixiu Zeng) and J.S.; visualization, Y.Z. (Yixiu Zeng); supervision, J.S. and H.S.; project administration, H.S.; funding acquisition, J.S. and H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This project was supported by Major Science and Technology Special Projects in Tibet Autonomous Region (XZ202201ZD0001N), University Innovation Research Group in Chongqing: (Grant No. CXQT21007), Chongqing TCM Key Discipline Construction Project (TCM Rehabilitation Science, Grant No. 2021-4322190044), Natural Science Foundation of Chongqing (Grant No. cstc2021jcyj-msxmX0772), and Fundamental Research Funds for the Central Universities (Grant No. SWU019026).
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to privacy restrictions.
Acknowledgments
RIN-14B cells were gifted by Weifang Rong (School of Medicine, Shanghai Jiao Tong University).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, H.; Wei, J.; Call, T.; Quintus, N.T.; Summers, A.J.; Carotenuto, S.; Johnson, R.; Ma, X.; Xu, C.; Park, J.G.; et al. Shisa6 mediates cell-type specific regulation of depression in the nucleus accumbens. Mol. Psychiatry 2021, 26, 7316–7327. [Google Scholar] [CrossRef] [PubMed]
- Steel, Z.; Marnane, C.; Iranpour, C.; Chey, T.; Jackson, J.W.; Patel, V.; Silove, D. The global prevalence of common mental disorders: A systematic review and meta-analysis 1980–2013. Int. J. Epidemiol. 2014, 43, 476–493. [Google Scholar] [CrossRef] [PubMed]
- Semahegn, A.; Torpey, K.; Manu, A.; Assefa, N.; Tesfaye, G.; Ankomah, A. Psychotropic medication non-adherence and its associated factors among patients with major psychiatric disorders: A systematic review and meta-analysis. Syst. Rev. Pharm. 2020, 9, 17. [Google Scholar] [CrossRef]
- Pereira, C.A.; Rodrigues, F.L.; Ruginsk, S.G.; Zanotto, C.Z.; Rodrigues, J.A.; Duarte, D.A.; Costa-Neto, C.M.; Resstel, L.B.; Carneiro, F.S.; Tostes, R.C. Chronic treatment with fluoxetine modulates vascular adrenergic responses by inhibition of pre- and post-synaptic mechanisms. Eur. J. Pharmacol. 2017, 800, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Gruenbaum, B.F.; Zlotnik, A.; Frenkel, A.; Fleidervish, I.; Boyko, M. Glutamate Efflux across the Blood-Brain Barrier: New Perspectives on the relationship between depression and the glutamatergic system. Metabolites 2022, 12, 459. [Google Scholar] [CrossRef] [PubMed]
- Rayff da Silva, P.; Diniz Nunes Pazos, N.; Karla Silva do Nascimento Gonzaga, T.; Cabral de Andrade, J.; Brito Monteiro, Á.; Caroline Ribeiro Portela, A.; Fernandes Oliveira Pires, H.; Dos Santos Maia, M.; Vilar da Fonsêca, D.; T Scotti, M.; et al. Anxiolytic and antidepressant-like effects of monoterpene tetrahydrolinalool and in silico approach of new potential targets. Curr. Top. Med. Chem. 2022, 22, 1530–1552. [Google Scholar]
- Xu, J.; Tang, M.; Wu, X.; Kong, X.; Liu, Y.; Xu, X. Lactobacillus rhamnosus zz-1 exerts preventive effects on chronic unpredictable mild stress-induced depression in mice via regulating the intestinal microenvironment. Food Funct. 2022, 13, 4331–4343. [Google Scholar] [CrossRef]
- Hao, W.; Ma, Q.; Tao, G.; Huang, J.; Chen, J. Oral coniferyl ferulate attenuated depression symptoms in mice via reshaping gut microbiota and microbial metabolism. Food Funct. 2021, 12, 12550–12564. [Google Scholar] [CrossRef]
- Tian, P.; Zhu, H.; Zou, R.; Kong, Q.; Xu, M.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. An in vitro screening method for probiotics with antidepressant-like effect using the enterochromaffin cell model. Food Funct. 2021, 12, 646–655. [Google Scholar] [CrossRef]
- Harmer, C.J. Serotonin and emotional processing: Does it help explain antidepressant drug action? Neuropharmacology 2008, 55, 1023–1028. [Google Scholar] [CrossRef]
- Pardridge, W.M. The role of blood-brain barrier transport of tryptophan and other neutral amino acids in the regulation of substrate-limited pathways of brain amino acid metabolism. J. Neural Transm. Suppl. 1979, 15, 43–54. [Google Scholar]
- Dehhaghi, M.; Kazemi Shariat Panahi, H.; Guillemin, G.J. Microorganisms, tryptophan metabolism, and kynurenine pathway: A complex interconnected loop influencing human health status. Int. J. Tryptophan Res. 2019, 12, 1178646919852996. [Google Scholar] [CrossRef] [PubMed]
- Teunis, C.; Nieuwdorp, M.; Hanssen, N. Interactions between tryptophan metabolism, the gut microbiome and the immune system as potential drivers of non-alcoholic fatty liver disease (NAFLD) and metabolic diseases. Metabolites 2022, 12, 514. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin synthesis and function: Evolutionary history in animals and plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef]
- Sharma, A.; Castellani, R.J.; Smith, M.A.; Muresanu, D.F.; Dey, P.K.; Sharma, H.S. 5-Hydroxytryptophan: A precursor of serotonin influences regional blood-brain barrier breakdown, cerebral blood flow, brain edema formation, and neuropathology. Int. Rev. Neurobiol. 2019, 146, 1–44. [Google Scholar]
- Fukuda, K. 5-HTP hypothesis of schizophrenia. Med. Hypotheses 2014, 82, 20–23. [Google Scholar] [CrossRef]
- Jangid, P.; Malik, P.; Singh, P.; Sharma, M.; Gulia, A.K.D. Comparative study of efficacy of l-5-hydroxytryptophan and fluoxetine in patients presenting with first depressive episode. Asian J. Psychiatr. 2013, 6, 29–34. [Google Scholar] [CrossRef]
- Nakajima, T.; Kudo, Y.; Kaneko, Z. Clinical evaluation of 5-hydroxy-L-tryptophan as an antidepressant drug. Folia Psychiatr. Neurol. Jpn. 1978, 32, 223–230. [Google Scholar] [CrossRef]
- Sha, J.; Song, J.; Huang, Y.; Zhang, Y.; Wang, H.; Zhang, Y.; Suo, H. Inhibitory effect and potential mechanism of Lactobacillus plantarum YE4 against dipeptidyl peptidase-4. Foods 2021, 11, 80. [Google Scholar] [CrossRef]
- Wang, W.; Xu, C.; Zhou, X.; Zhang, L.; Gu, L.; Liu, Z.; Ma, J.; Hou, J.; Jiang, Z. Lactobacillus plantarum combined with Galactooligosaccharides supplement: A neuroprotective regimen against neurodegeneration and memory impairment by regulating short-chain fatty acids and the c-Jun N-Terminal kinase signaling pathway in mice. J. Agric. Food Chem. 2022, 70, 8619–8630. [Google Scholar] [CrossRef]
- Wei, Z.; Zhou, N.; Zou, L.; Shi, Z.; Dun, B.; Ren, G.; Yao, Y. Soy protein alleviates malnutrition in weaning rats by regulating gut microbiota composition and serum metabolites. Front. Nutr. 2021, 8, 774203. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Farzi, A.; Ke, X.; Yu, Y.; Chen, C.; Chen, S.; Yu, T.; Wang, H.; Li, Y. Oral administration of Lactococcus lactis WHH2078 alleviates depressive and anxiety symptoms in mice with induced chronic stress. Food Funct. 2022, 13, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium with the role of 5-hydroxytryptophan synthesis regulation alleviates the symptom of depression and related microbiota dysbiosis. J. Nutr. Biochem. 2019, 66, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; O’Riordan, K.J.; Lee, Y.; Wang, G.; Zhao, J.; Zhang, H.; Cryan, J.F.; Chen, W. Towards a psychobiotic therapy for depression: Bifidobacterium breve CCFM1025 reverses chronic stress-induced depressive symptoms and gut microbial abnormalities in mice. Neurobiol. Stress 2020, 12, 100216. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, K.; Kawabata-Shoda, E.; Doihara, H.; Kojima, R.; Okada, H.; Mochizuki, S.; Sano, Y.; Inamura, K.; Matsushime, H.; Koizumi, T.; et al. TRPA1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3408–3413. [Google Scholar] [CrossRef]
- Li, H.; Li, L.; Chen, H.; Wang, R.; Ma, Y. The structure and properties of MFG-E8 and the In vitro assessment of its toxic effects on myoblast cells. Protein Expr. Purif. 2021, 178, 105720. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cui, L.; Zhang, P.; Dong, T.; Fang, J. Transcriptome and metabolite profiling reveal that spraying calcium fertilizer reduces grape berry cracking by modulating the flavonoid biosynthetic metabolic pathway. Food Chem. 2021, 2, 100025. [Google Scholar] [CrossRef]
- Chen, J.; Ren, Y.; Zhang, K.; Xiong, Y.L.; Wang, Q.; Shang, K.; Zhang, D. Site-specific incorporation of sodium tripolyphosphate into myofibrillar protein from mantis shrimp (Oratosquilla oratoria) promotes protein crosslinking and gel network formation. Food Chem. 2020, 312, 126113. [Google Scholar] [CrossRef]
- Zettlitz, K.A.; Lorenz, V.; Landauer, K.; Munkel, S.; Herrmann, A.; Scheurich, P.; Pfizenmaier, K.; Kontermann, R. ATROSAB, a humanized antagonistic anti-tumor necrosis factor receptor one-specific antibody. mAbs 2010, 2, 639–647. [Google Scholar] [CrossRef]
- Liu, S.; Guo, R.; Liu, F.; Yuan, Q.; Yu, Y.; Ren, F. Gut microbiota regulates depression-like behavior in rats through the neuroendocrine-immune-mitochondrial pathway. Neuropsychiatr. Dis. Treat. 2020, 16, 859–869. [Google Scholar] [CrossRef]
- Kotni, M.K.; Zhao, M.; Wei, D.Q. Gene expression profiles and protein-protein interaction networks in amyotrophic lateral sclerosis patients with C9orf72 mutation. Orphanet J. Rare Dis. 2016, 11, 148. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zhou, W. Amarogentin has protective effects against sepsis-induced brain injury via modulating the AMPK/SIRT1/NF-κB pathway. Brain Res. Bull. 2022, 189, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, B.; Ai, L. Advances in the microbial synthesis of 5-Hydroxytryptophan. Front. Bioeng. Biotechnol. 2021, 9, 624503. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.B.; Brinckmann, J.A.; Harter, D.E.V. From forest to pharmacy: Should we be depressed about a sustainable Griffonia simplicifolia (Fabaceae) seed supply chain? J. Ethnopharmacol. 2021, 278, 114202. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ran, L.; Wu, Y.; Liang, M.; Zeng, J.; Ke, F.; Wang, F.; Yang, J.; Lao, X.; Liu, L.; et al. Oral administration of 5-hydroxytryptophan restores gut microbiota dysbiosis in a mouse model of depression. Front. Microbiol. 2022, 13, 864571. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Li, X.; Zheng, J. Transcriptome profiling using Illumina- and SMRT-based RNA-seq of hot pepper for in-depth understanding of genes involved in CMV infection. Gene 2018, 666, 123–133. [Google Scholar] [CrossRef]
- Leonard, B.E. Inflammation and depression: A causal or coincidental link to the pathophysiology? Acta Neuropsychiatr. 2018, 30, 1–16. [Google Scholar] [CrossRef]
- Das, R.; Emon, M.P.Z.; Shahriar, M.; Nahar, Z.; Islam, S.M.A.; Bhuiyan, M.A.; Islam, S.N.; Islam, M.R. Higher levels of serum IL-1β and TNF-α are associated with an increased probability of major depressive disorder. Psychiatry Res. 2021, 295, 113568. [Google Scholar] [CrossRef]
- Bavaresco, D.V.; Uggioni, M.L.R.; Ferraz, S.D.; Marques, R.M.M.; Simon, C.S.; Dagostin, V.S.; Grande, A.J.; da Rosa, M.I. Efficacy of infliximab in treatment-resistant depression: A systematic review and meta-analysis. Pharmacol. Biochem. Behav. 2020, 188, 172838. [Google Scholar] [CrossRef]
- McIntyre, R.S.; Subramaniapillai, M.; Lee, Y.; Pan, Z.; Carmona, N.E.; Shekotikhina, M.; Rosenblat, J.D.; Brietzke, E.; Soczynska, J.K.; Cosgrove, V.E.; et al. Efficacy of adjunctive infliximab vs placebo in the treatment of adults with bipolar I/II depression: A randomized clinical trial. JAMA Psychiatry 2019, 76, 783–790. [Google Scholar] [CrossRef]
- Uzzan, S.; Azab, A.N. Anti-TNF-α compounds as a treatment for depression. Molecules 2021, 26, 2368. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wang, T.; Hu, X.; Luo, J.; Li, W.; Wu, X.; Duan, Y.; Jin, F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience 2015, 310, 561–577. [Google Scholar] [CrossRef]
- Rozpedek, W.; Markiewicz, L.; Diehl, J.A.; Pytel, D.; Majsterek, I. Unfolded protein response and PERK Kinase as a new therapeutic target in the pathogenesis of Alzheimer’s disease. Curr. Med. Chem. 2015, 22, 3169–3184. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, W.; Yin, J.; Chen, Y.; Guo, S.; Fan, H.; Li, X.; Zhang, X.; He, X.; Duan, C. TSG-6 attenuates inflammation-induced brain injury via modulation of microglial polarization in SAH rats through the SOCS3/STAT3 pathway. J. Neuroinflamm. 2018, 15, 231. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Kurihara, H.; Morita, H.; Cao, W.H.; Ling, G.Y.; Kumada, M.; Kimura, S.; Nagai, R.; Yazaki, Y.; Kuwaki, T. Role of endothelin-1 in stress response in the central nervous system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R515–R521. [Google Scholar] [CrossRef] [PubMed]
- Pantazatos, S.P.; Huang, Y.; Rosoklija, G.B.; Dwork, A.J.; Arango, V.; Mann, J.J. Whole-transcriptome brain expression and exon-usage profiling in major depression and suicide: Evidence for altered glial, endothelial and ATPase activity. Mol. Psychiatry 2017, 22, 760–773. [Google Scholar] [CrossRef]
- Lari, A.; Gholami Pourbadie, H.; Sharifi-Zarchi, A.; Aslani, S.; Nejatbakhsh Samimi, L.; Jamshidi, A.; Mahmoudi, M. Evaluation of the ankylosing spondylitis transcriptome for oxidative phosphorylation pathway: The shared pathway with neurodegenerative diseases. Iran. J. Allergy Asthma Immunol. 2021, 20, 563–573. [Google Scholar] [CrossRef]
- Beroun, A.; Mitra, S.; Michaluk, P.; Pijet, B.; Stefaniuk, M.; Kaczmarek, L. MMPs in learning and memory and neuropsychiatric disorders. Cell. Mol. Life Sci. 2019, 76, 3207–3228. [Google Scholar] [CrossRef]
- Nagy, V.; Bozdagi, O.; Matynia, A.; Balcerzyk, M.; Okulski, P.; Dzwonek, J.; Costa, R.M.; Silva, A.J.; Kaczmarek, L.; Huntley, G.W. Matrix metalloproteinase-9 is required for hippocampal late-phase long-term potentiation and memory. J. Neurosci. 2006, 26, 1923–1934. [Google Scholar] [CrossRef]
- Coba de la Peña, T.; Redondo, F.J.; Fillat, M.F.; Lucas, M.M.; Pueyo, J.J. Flavodoxin overexpression confers tolerance to oxidative stress in beneficial soil bacteria and improves survival in the presence of the herbicides paraquat and atrazine. J. Appl. Microbiol. 2013, 115, 236–246. [Google Scholar] [CrossRef]
- D’Cunha, N.M.; Sergi, D.; Lane, M.M.; Naumovski, N.; Gamage, E.; Rajendran, A.; Kouvari, M.; Gauci, S.; Dissanayka, T.; Marx, W.; et al. The effects of dietary advanced glycation End-products on neurocognitive and mental disorders. Nutrients 2022, 14, 2421. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).