Diversity of Filamentous Fungi Associated with Dairy Processing Environments and Spoiled Products in Brazil

 , , , , , , , , and
, , , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
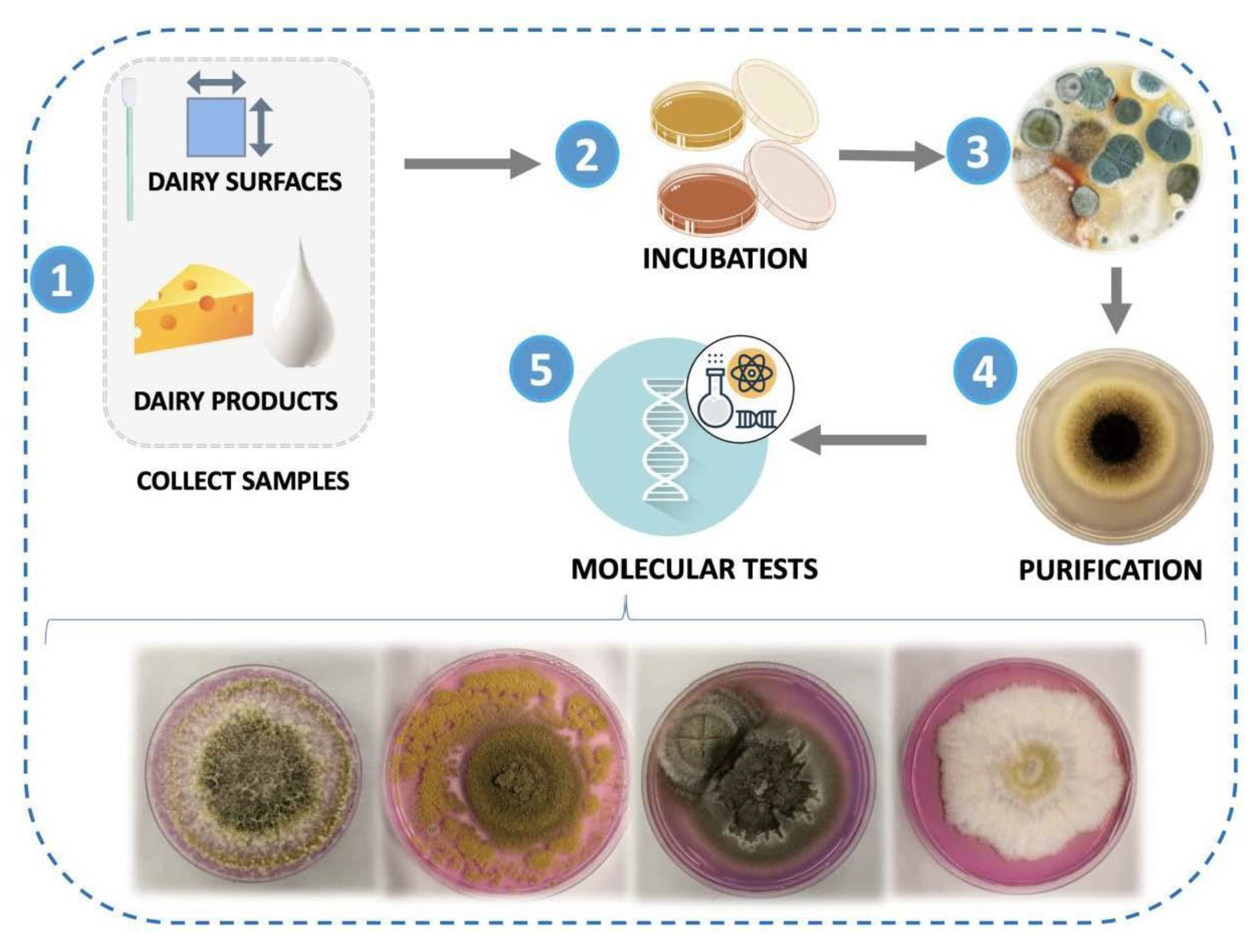
2.1. Isolation of Filamentous Fungi
2.2. Phenotypic Characterization
2.3. Molecular Identification of Fungal Isolates
2.3.1. Extraction of Fungal DNA
2.3.2. PCR Amplification, DNA Sequencing, and BLAST Identification
2.4. Phylogenetic Analysis
3. Results
3.1. Isolation of Filamentous Fungi
3.2. Identification of Fungal Isolates
3.3. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations The State of Food and Agriculture. Moving Forward on Food Loss and Waste Reduction; FAO: Rome, Italy, 2019; pp. 2–13. [Google Scholar]
- United Nations Environment Programme UNEP Food Waste Index—Report 2021. Available online: https://www.unep.org/resources/report/unep-food-waste-index-report-2021 (accessed on 13 November 2022).
- United States Department of Agriculture (USDA); Environmental Protection Agency (EPA) U.S. Food Loss & Waste 2030 Champions. Milestones Report—May 2021. Available online: https://www.usda.gov/sites/default/files/documents/food-loss-waste-champions-report.pdf (accessed on 12 November 2022).
- Brooks, J.C.; Martinez, B.; Stratton, J.; Bianchini, A.; Krokstrom, R.; Hutkins, R. Survey of Raw Milk Cheeses for Microbiological Quality and Prevalence of Foodborne Pathogens. Food Microbiol. 2012, 31, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Pitt John, I.; Hocking, A.D. The Ecology of Fungal Food Spoilage. In Fungi and Food Spoilage; Springer: Boston, MA, USA, 2009; pp. 3–9. ISBN 978-0-387-92207-2. [Google Scholar]
- Pattono, D.; Grosso, A.; Stocco, P.P.; Pazzi, M.; Zeppa, G. Survey of the Presence of Patulin and Ochratoxin A in Traditional Semi-Hard Cheeses. Food Control 2013, 33, 54–57. [Google Scholar] [CrossRef]
- Ráduly, Z.; Szabó, L.; Madar, A.; Pócsi, I.; Csernoch, L. Toxicological and Medical Aspects of Aspergillus-Derived Mycotoxins Entering the Feed and Food Chain. Front. Microbiol. 2020, 10, 2908. [Google Scholar] [CrossRef] [Green Version]
- Parussolo, G.; Bernardi, A.O.; Garcia, M.V.; Stefanello, A.; dos Santos Silva, T.; Copetti, M.V. Fungi in Air, Raw Materials and Surface of Dry Fermented Sausage Produced in Brazil. LWT 2019, 108, 190–198. [Google Scholar] [CrossRef]
- Pinches, S.E.; Apps, P. Production in Food of 1,3-Pentadiene and Styrene by Trichoderma Species. Int. J. Food Microbiol. 2007, 116, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Leyva Salas, M.; Mounier, J.; Valence, F.; Coton, M.; Thierry, A.; Coton, E. Antifungal Microbial Agents for Food Biopreservation—A Review. Microorganisms 2017, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, K.; Passeró, E.; Vallone, L.; Hymery, N.; Coton, M.; Jany, J.-L.; Mounier, J.; Coton, E. Occurrence of Roquefortine C, Mycophenolic Acid and Aflatoxin M1 Mycotoxins in Blue-Veined Cheeses. Food Control 2015, 47, 634–640. [Google Scholar] [CrossRef]
- Dobson, A.D.W. Chapter 23—Mycotoxins in Cheese. In Cheese, 4th ed.; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: San Diego, CA, USA, 2017; pp. 595–601. ISBN 978-0-12-417012-4. [Google Scholar]
- Anelli, P.; Haidukowski, M.; Epifani, F.; Cimmarusti, M.T.; Moretti, A.; Logrieco, A.; Susca, A. Fungal Mycobiota and Mycotoxin Risk for Traditional Artisan Italian Cave Cheese. Food Microbiol 2019, 78, 62–72. [Google Scholar] [CrossRef]
- Instituto Brasileiro de Geografia e Estatística (IBGE) Pesquisa Trimestral Do Leite. Available online: https://www.ibge.gov.br/estatisticas/economicas/agricultura-e-pecuaria/9209-pesquisa-trimestral-do-leite.html?=&t=destaques (accessed on 12 November 2022).
- Kure, C.F.; Wasteson, Y.; Brendehaug, J.; Skaar, I. Mould Contaminants on Jarlsberg and Norvegia Cheese Blocks from Four Factories. Int. J. Food Microbiol. 2001, 70, 21–27. [Google Scholar] [CrossRef]
- Kure, C.F.; Skaar, I.; Brendehaug, J. Mould Contamination in Production of Semi-Hard Cheese. Int. J. Food Microbiol. 2004, 93, 41–49. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Pawtowski, A.; Auhustsinava-Galerne, L.; Frotté, N.; Baroncelli, R.; Deniel, F.; Coton, E.; Mounier, J. Diversity of Spoilage Fungi Associated with Various French Dairy Products. Int. J. Food Microbiol. 2017, 241, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Buehler, A.J.; Evanowski, R.L.; Wiedmann, M.; Martin, N.H. Internal Transcribed Spacer (ITS) Sequence-Based Characterization of Fungal Isolates from Multiple Yogurt Facilities—A Case Study. J. Dairy Sci. 2019, 102, 3646–3653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, A.B.; Churey, J.J.; Worobo, R.W. Association of Fungal Genera from Spoiled Processed Foods with Physicochemical Food Properties and Processing Conditions. Food Microbiol. 2019, 83, 211–218. [Google Scholar] [CrossRef] [PubMed]
- de Souza, T.P.; Evangelista, S.R.; Passamani, F.R.F.; Bertechini, R.; de Abreu, L.R.; Batista, L.R. Mycobiota of Minas Artisanal Cheese: Safety and Quality. Int. Dairy J. 2021, 120, 105085. [Google Scholar] [CrossRef]
- le Lay, C.; Mounier, J.; Vasseur, V.; Weill, A.; le Blay, G.; Barbier, G.; Coton, E. In Vitro and in Situ Screening of Lactic Acid Bacteria and Propionibacteria Antifungal Activities against Bakery Product Spoilage Molds. Food Control 2016, 60, 247–255. [Google Scholar] [CrossRef]
- Legnani, P.; Leoni, E.; Berveglieri, M.; Mirolo, G.; Alvaro, N. Hygienic Control of Mass Catering Establishments, Microbiological Monitoring of Food and Equipment. Food Control 2004, 15, 205–211. [Google Scholar] [CrossRef]
- Visagie, C.M.; Hirooka, Y.; Tanney, J.B.; Whitfield, E.; Mwange, K.; Meijer, M.; Amend, A.S.; Seifert, K.A.; Samson, R.A. Aspergillus, Penicillium and Talaromyces Isolated from House Dust Samples Collected around the World. Stud. Mycol. 2014, 78, 63–139. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Journal of Molecular Evolution Evolutionary Trees from DNA Sequences: A Maximum Likelihood Approach. J Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions Through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the Number of Nucleotide Substitutions When There Are Strong Transition-Transversion and G+C-Content Biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef]
- Kuhnert, E.; Fournier, J.; Peršoh, D.; Luangsa-ard, J.J.D.; Stadler, M. New Hypoxylon Species from Martinique and New Evidence on the Molecular Phylogeny of Hypoxylon Based on ITS RDNA and β-Tubulin Data. Fungal Divers. 2014, 64, 181–203. [Google Scholar] [CrossRef]
- Abdolrasouli, A.; Gibani, M.M.; de Groot, T.; Borman, A.M.; Hoffman, P.; Azadian, B.S.; Mughal, N.; Moore, L.S.P.; Johnson, E.M.; Meis, J.F. A Pseudo-Outbreak of Rhinocladiella Similis in a Bronchoscopy Unit of a Tertiary Care Teaching Hospital in London, United Kingdom. Mycoses 2021, 64, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Gareth Jones, E.B.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal Diversity Notes 709–839: Taxonomic and Phylogenetic Contributions to Fungal Taxa with an Emphasis on Fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Herrera-Balandrano, D.D.; Shi, X.-C.; Wang, S.-Y.; Laborda, P. Biocontrol of Colletotrichum Brevisporum in Soybean Using a New Genistein-Producing Paecilomyces Strain. Biol. Control 2022, 169, 104877. [Google Scholar] [CrossRef]
- Codex Alimentarius Commission General Standar for Cheese (CXS 283-1978). Available online: https://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B283-1978%252FCXS_283e.pdf (accessed on 13 November 2022).
- Nájera, A.I.; Nieto, S.; Barron, L.J.R.; Albisu, M. A Review of the Preservation of Hard and Semi-Hard Cheeses: Quality and Safety. Int. J. Environ. Res. Public Health 2021, 18, 9789. [Google Scholar] [CrossRef]
- Decontardi, S.; Mauro, A.; Lima, N.; Battilani, P. Survey of Penicillia Associated with Italian Grana Cheese. Int. J. Food Microbiol. 2017, 246, 25–31. [Google Scholar] [CrossRef]
- Lund, F.; Filtenborg, O.; Frisvad, J.C. Associated Mycoflora of Cheese. Food Microbiol. 1995, 12, 173–180. [Google Scholar] [CrossRef]
- Kandasamy, S.; Park, W.S.; Yoo, J.; Yun, J.; Kang, H.B.; Seol, K.-H.; Oh, M.-H.; Ham, J.S. Characterisation of Fungal Contamination Sources for Use in Quality Management of Cheese Production Farms in Korea. Asian-Australas. J. Anim. Sci. 2020, 33, 1002–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, P.; Palmero, D.; Jurado, M. Occurrence of Moulds Associated with Ovine Raw Milk and Cheeses of the Spanish Region of Castilla La Mancha. Int. J. Dairy Technol. 2015, 68, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Moubasher, A.-A.H.; Abdel-Sater, M.A.; Soliman, Z.S.M. Yeasts and Filamentous Fungi Associated with Some Dairy Products in Egypt. J. Mycol. Med. 2018, 28, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Torkar, K.G.; Vengušt, A. The Presence of Yeasts, Moulds and Aflatoxin M1 in Raw Milk and Cheese in Slovenia. Food Control 2008, 19, 570–577. [Google Scholar] [CrossRef]
- Hymery, N.; Vasseur, V.; Coton, M.; Mounier, J.; Jany, J.L.; Barbier, G.; Coton, E. Filamentous Fungi and Mycotoxins in Cheese: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 437–456. [Google Scholar] [CrossRef]
- Akinyemi, M.O.; Braun, D.; Windisch, P.; Warth, B.; Ezekiel, C.N. Assessment of Multiple Mycotoxins in Raw Milk of Three Different Animal Species in Nigeria. Food Control 2022, 131, 108258. [Google Scholar] [CrossRef]
- Pinhão, M.; Tavares, A.M.; Loureiro, S.; Louro, H.; Alvito, P.; Silva, M.J. Combined Cytotoxic and Genotoxic Effects of Ochratoxin A and Fumonisin B1 in Human Kidney and Liver Cell Models. Toxicol Vitr. 2020, 68, 104949. [Google Scholar] [CrossRef]
- Claeys, L.; de Saeger, S.; Scelo, G.; Biessy, C.; Casagrande, C.; Nicolas, G.; Korenjak, M.; Fervers, B.; Heath, A.K.; Krogh, V.; et al. Mycotoxin Exposure and Renal Cell Carcinoma Risk: An Association Study in the EPIC European Cohort. Nutrients 2022, 14, 3581. [Google Scholar] [CrossRef]
- Cenci-Goga, B.; Cruciani, D.; Crotti, S.; Karama, M.; Ylldlrlm, G.; Bulut, M.; Marino, C.; Grispoldi, L. Diversity of Yeasts and Moulds in Dairy Products from Umbria, Central Italy. J. Dairy Res. 2021, 88, 217–220. [Google Scholar] [CrossRef]
- Barrios, M.J.; Medina, L.M.; Lopez, M.C.; Jordano, R. Fungal Biota Isolated from Spanish Cheeses. J. Food Saf. 1998, 18, 151–157. [Google Scholar] [CrossRef]
- Montagna, M.T.; Santacroce, M.P.; Spilotros, G.; Napoli, C.; Minervini, F.; Papa, A.; Dragoni, I. Investigation of Fungal Contamination in Sheep and Goat Cheeses in Southern Italy. Mycopathologia 2004, 158, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Seddek, N.H.; Gomah, N.H.; Osman, D.M. Fungal Flora Contaminating Egyptian Ras Cheese with Reference to Their Toxins and Enzymes. Food Sci. Technol. 2016, 4, 64–68. [Google Scholar] [CrossRef]
- Mbareche, H.; Veillette, M.; Bilodeau, G.J.; Duchaine, C. Fungal Aerosols at Dairy Farms Using Molecular and Culture Techniques. Sci. Total Environ. 2019, 653, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Moubasher, A.H.; Abdel-Kader, M.I.A.; El-Kady, I.A. Toxigenic Fungi Isolated from Roquefort Cheese. Mycopathologia 1979, 66, 187–190. [Google Scholar] [CrossRef]
- Giri, D.K.; Sims, W.P.; Sura, R.; Cooper, J.J.; Gavrilov, B.K.; Mansell, J. Cerebral and Renal Phaeohyphomycosis in a Dog Infected with Bipolaris Species. Vet. Pathol. 2011, 48, 754–757. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, O.; Spirin, V.; Vlasák, J.; Rivoire, B.; Stenroos, S.; Hibbett, D.S. Polypores and Genus Concepts in Phanerochaetaceae (Polyporales, Basidiomycota). MycoKeys 2016, 17, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.-H.; Ryvarden, L.; et al. A Revised Family-Level Classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and Phylogenetic Contributions to Fungi Associated with the Invasive Weed Chromolaena Odorata (Siam Weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Schroers, H.-J. A Monograph of Bionectria (Ascomycota, Hypocreales, Bionectriaceae) and Its Clonostachys Anamorphs. Stud. Mycol. 2001, 46, 1–214. [Google Scholar]
- Moraga-Suazo, P.; Opazo, A.; Zaldúa, S.; González, G.; Sanfuentes, E. Evaluation of Trichoderma Spp. and Clonostachys Spp. Strains to Control Fusarium Circinatum in Pinus Radiata Seedlings. Chil. J. Agric. Res. 2011, 71, 412–417. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Vicente, V.; Caligiorne, R.B.; Kantarcioglu, S.; Tintelnot, K.; van den Ende, A.H.G.G.; Haase, G. Species Diversity and Polymorphism in the Exophiala Spinifera Clade Containing Opportunistic Black Yeast-Like Fungi. J. Clin. Microbiol. 2003, 41, 4767–4778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidrich, D.; González, G.M.; Pagani, D.M.; Ramírez-Castrillón, M.; Scroferneker, M.L. Chromoblastomycosis Caused by Rhinocladiella Similis: Case Report. Med. Mycol. Case Rep. 2017, 16, 25–27. [Google Scholar] [CrossRef]
- De Andrade, T.S.; de Almeida, A.M.Z.; Basano, S.d.A.; Takagi, E.H.; Szeszs, M.W.; Melhem, M.S.C.; Albuquerque, M.; Camargo, J.d.S.A.A.; Gambale, W.; Camargo, L.M.A. Chromoblastomycosis in the Amazon Region, Brazil, Caused by Fonsecaea Pedrosoi, Fonsecaea Nubica, and Rhinocladiella Similis: Clinicopathology, Susceptibility, and Molecular Identification. Med. Mycol. 2020, 58, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.W.; Groenewald, J.Z.; Pfenning, L.H.; Yarden, O.; Crous, P.W.; Cai, L. The Phoma-like Dilemma. Stud. Mycol. 2020, 96, 309–396. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identification and Number of Isolates | Mozzarela Cheese (n = 3) | Reino Cheese (n = 2) | Montanhês Cheese (n = 3) | Parmesan Cheese (n = 2) | Ghee Butter (n = 2) | Gorgonzola Cheese (n = 1) | Provolone Cheese (n = 2) | Processed Cheese (n = 2) | Coalho Cheese (n = 1) | Total % in Spoiled Dairy Products | Total % in Air, Equipment, and Tools |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Penicillium (36) | 2 | 5 | 2 | 4 | 2 | 5 | 3 | - | 2 | 25 (22.94%) | 11 (10.09%) |
| Cladosporium (23) | - | - | 2 | - | - | - | - | 2 | 2 | 6 (5.50%) | 17 (15.60%) |
| Nigrospora (5) | - | - | - | - | - | - | - | - | - | - | 5 (4.58%) |
| Riopa (5) | - | - | - | - | - | - | - | - | - | - | 5 (4.58%) |
| Aspergillus (4) | - | - | - | - | - | - | - | - | - | - | 4 (3.67%) |
| Hipoxylon (2) | - | - | - | - | - | - | - | - | - | - | 2 (1.83%) |
| Fusarium (1) | - | - | 1 | - | - | - | - | - | - | 1 (0.92%) | - |
| Montagnula (1) | - | - | - | - | - | - | - | - | - | - | 1 (0.92%) |
| Clonostachys (1) | - | - | - | - | - | - | - | - | - | - | 1(0.92%) |
| Phaeosphaeria (1) | - | - | - | - | - | - | - | - | 1 | 1(0.92%) | - |
| Rhinocladiella (1) | - | - | - | - | - | - | - | - | - | - | 1(0.92%) |
| Coniochaeta (1) | - | - | - | - | - | - | - | - | - | - | 1(0.92%) |
| Trichoderma (1) | - | - | - | - | - | - | - | - | - | - | 1(0.92%) |
| Paecilomyces (1) | - | - | - | - | - | - | - | - | - | - | 1(0.92%) |
| Didymella (1) | - | - | - | - | - | - | - | - | - | - | 1(0.92%) |
| Bipolaris (1) | - | - | - | - | - | - | - | - | - | - | 1(0.92%) |
| Not identified (24) | 3 | 2 | 2 | - | - | - | - | 1 | - | 8 (7.33%) | 16(14.68%) |
| Total | 5 | 7 | 7 | 4 | 5 | 3 | 3 | 3 | 5 | 41 | 68 |
| Total (%) | 12.19 | 17.07 | 17.07 | 9.76 | 12.19 | 7.33 | 7.33 | 7.33 | 12.19 | 37.61 | 62.39 |
| Identification | Ware House | Cold Chamber | Ghee Stock Room | Product Refrigerators | Cheese Ripening Room | Gorgonzola Ripening Room | Production Room | Cheese Packing Room | Brine Room | Production Tables | Cheese Containers | Cheese Packaging | Cheese Tank |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Penicillium sp. | - | 1 | - | - | 1 | - | - | 1 | - | - | 6 | - | 2 |
| Cladosporium sp. | 1 | 1 | 2 | 2 | 4 | - | 2 | 1 | 1 | 1 | - | 1 | 1 |
| Nigrospora sp. | - | - | 1 | - | - | - | 1 | 2 | - | - | - | - | 1 |
| Riopa sp. | - | 1 | - | 1 | 1 | - | 1 | - | - | - | - | - | 1 |
| Aspergillus sp. | 1 | - | - | 1 | - | - | 1 | - | - | - | - | - | 1 |
| Hipoxylon sp. | - | - | - | - | - | - | - | - | - | - | - | - | 2 |
| Fusarium sp. | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Montagnula sp. | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Clonostachys sp. | - | - | 1 | - | - | - | - | - | - | - | - | - | - |
| Phaeosphaeria sp. | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Rhinocladiella sp. | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Coniochaeta sp. | - | - | - | - | - | - | 1 | - | - | - | - | - | - |
| Trichoderma sp. | - | - | - | - | - | 1 | - | - | - | - | - | - | - |
| Paecilomyces sp. | - | - | - | - | - | - | - | - | 1 | - | - | - | - |
| Didymella sp. | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| Bipolaris sp. | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| Not identified | 3 | - | 2 | - | 1 | - | 6 | 1 | - | 1 | 1 | 1 | - |
| Total | 5 | 3 | 6 | 4 | 7 | 1 | 14 | 5 | 2 | 2 | 8 | 3 | 8 |
| Total (%) | 7.35 | 4.42 | 8.82 | 5.89 | 10.29 | 1.47 | 20.59 | 7.35 | 2.94 | 2.94 | 11.76 | 4.42 | 11.76 |
| Sample | Length (nt) | Identification | BLASTN | ||
|---|---|---|---|---|---|
| Query Coverage | Percent Identify | Accession Number | |||
| 1.21 | 570 | H. griseobrunneum | 97% | 98.92% | NR_155184.1 |
| 4.24 | 640 | R. similis | 96% | 100.00% | NR_166008.1 |
| 4.27 | 600 | C. rosae | 94% | 95.13% | NR_157509.1 |
| 4.38 | 630 | P. maximus | 90% | 96.84% | NR_149329.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, L.V.; Rodrigues, R.d.S.; Fusieger, A.; da Silva, R.R.; de Jesus Silva, S.R.; Martins, E.; Machado, S.G.; Caggia, C.; Randazzo, C.L.; de Carvalho, A.F. Diversity of Filamentous Fungi Associated with Dairy Processing Environments and Spoiled Products in Brazil. Foods 2023, 12, 153. https://doi.org/10.3390/foods12010153
Souza LV, Rodrigues RdS, Fusieger A, da Silva RR, de Jesus Silva SR, Martins E, Machado SG, Caggia C, Randazzo CL, de Carvalho AF. Diversity of Filamentous Fungi Associated with Dairy Processing Environments and Spoiled Products in Brazil. Foods. 2023; 12(1):153. https://doi.org/10.3390/foods12010153
Chicago/Turabian StyleSouza, Luana Virgínia, Rafaela da Silva Rodrigues, Andressa Fusieger, Raiane Rodrigues da Silva, Sidney Rodrigues de Jesus Silva, Evandro Martins, Solimar Gonçalves Machado, Cinzia Caggia, Cinzia Lucia Randazzo, and Antonio Fernandes de Carvalho. 2023. "Diversity of Filamentous Fungi Associated with Dairy Processing Environments and Spoiled Products in Brazil" Foods 12, no. 1: 153. https://doi.org/10.3390/foods12010153