
Metabolomics Profiling of White Button, Crimini, Portabella, Lion’s Mane, Maitake, Oyster, and Shiitake Mushrooms Using Untargeted Metabolomics and Targeted Amino Acid Analysis
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Untargeted Metabolomics
2.1.1. Chemicals, Standards, and Reagents
2.1.2. Mushroom Procurement
2.1.3. Mushroom Sample Processing and Homogenization
2.1.4. Mushroom Sample Preparation
2.1.5. Hydrophobic Liquid Chromatography Mass Spectrometry (LC/MS)
2.1.6. Hydrophilic Liquid Chromatography Mass Spectrometry (LC/MS)
2.1.7. Data Processing
2.1.8. Compound Annotation
2.1.9. Data Visualization
2.1.10. Statistical Analysis
2.1.11. Compound Curation and Identification of Potential Mushroom-Specific Compounds
2.2. Amino Acid Analysis
2.2.1. Sample Preparation
2.2.2. Hydrophilic Interaction Liquid Chromatography—Triple Quadrupole Mass Spectrometry (HILIC-QQQ-MS)
- Xs = measured concentration in mg/mL;
- Vt = total volume of concentrated extract (in mL);
- D = dilution factor if sample was extracted before analysis;
- Vi = volume of extract injected (in µL);
- Ws = weight of sample extracted in g.
3. Results
3.1. Untargeted Metabolomics Analysis Detects Thousands of Compounds in Mushrooms
3.2. Mushroom Samples Group Based on Species as Depicted by Principal Component Analysis (PCA) and Hierarchical Clustering (HC)
3.3. Statistical Analysis—One-Way ANOVA
3.4. Several Compounds Are Unique-to-Mushroom-Variety
3.5. Untargeted Metabolomics Analysis Reveals Potential Mushroom-Specific Compounds
3.6. Amino Acid Profiles Vary among Different Mushroom Varieties
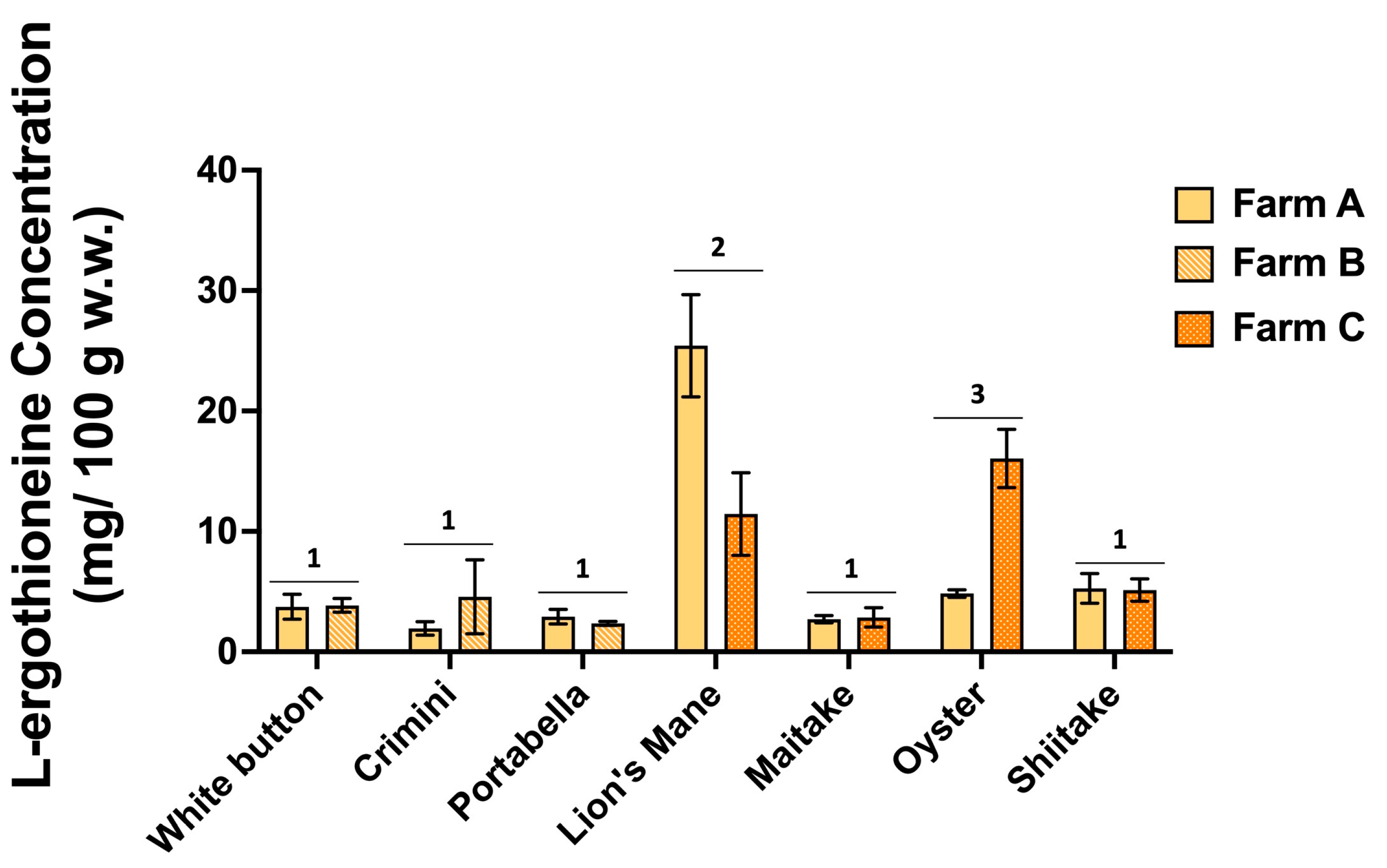
3.7. L-Ergothioneine Concentration Varies among Different Mushroom Varieties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feeney, M.J.; Miller, A.M.; Roupas, P. Mushrooms—Biologically Distinct and Nutritionally Unique: Exploring a “Third Food Kingdom”. Nutr. Today 2014, 49, 301–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach, F.; Helm, C.V.; Bellettini, M.B.; Maciel, G.M.; Haminiuk, C.W.I. Edible Mushrooms: A Potential Source of Essential Amino Acids, Glucans and Minerals. Int. J. Food Sci. Technol. 2017, 52, 2382–2392. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture (USDA) Agricultural Research Service. FoodData Central: Foundation Foods. White Button Mushrooms. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/1999629/nutrients (accessed on 31 July 2022).
- Dubost, N.J.; Beelman, R.B.; Peterson, D.; Royse, D.J. Identification and Quantification of Ergothioneine in Cultivated Mushrooms by Liquid Chromatography-Mass Spectroscopy. Int. J. Med. Mushrooms 2006, 8, 215–222. [Google Scholar] [CrossRef]
- Halliwell, B.; Cheah, I.K.; Drum, C.L. Ergothioneine, an Adaptive Antioxidant for the Protection of Injured Tissues? A Hypothesis. Biochem. Biophys. Res. Commun. 2016, 470, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Cheah, I.K.; Tang, R.M.Y. Ergothioneine—A Diet-Derived Antioxidant with Therapeutic Potential. FEBS Lett. 2018, 592, 3357–3366. [Google Scholar] [CrossRef] [Green Version]
- Cheah, I.K.; Halliwell, B. Ergothioneine; Antioxidant Potential, Physiological Function and Role in Disease. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 784–793. [Google Scholar] [CrossRef] [Green Version]
- Beelman, R.B.; Kalaris, M.D.; Phillips, A.T.; Richie, J.P., Jr. Is Ergothioneine a ‘Longevity Vitamin’ Limited in the American Diet? J. Nutr. Sci. 2020, 9, 1–5. [Google Scholar] [CrossRef]
- Prajapati, D.; Bhatt, A.; Gupte, S.; Gupte, A. Mushroom secondary metabolites: Chemistry and therapeutic applications. Int. J. Pharm. Sci. Res. 2021, 12, 5677–5689. [Google Scholar] [CrossRef]
- Cerletti, C.; Esposito, S.; Iacoviello, L. Edible Mushrooms and Beta-Glucans: Impact on Human Health. Nutrients 2021, 13, 2195. [Google Scholar] [CrossRef]
- Kala, K.; Kryczyk-Poprawa, A.; Rzewinska, A.; Muszynska, B. Fruiting Bodies of Selected Edible Mushrooms as a Potential Source of Lovastatin. Eur. Food Res. Technol. 2020, 246, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Witkowska, A.M.; Zujko, M.E.; Mirończuk-Chodakowska, I. Comparative Study of Wild Edible Mushrooms as Sources of Antioxidants. Int. J. Med. Mushrooms 2011, 13, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.A.; Rouf, R.; Tiralongo, E.; May, T.W.; Tiralongo, J. Mushroom Lectins: Specificity, Structure and Bioactivity Relevant to Human Disease. Int. J. Mol. Sci. 2015, 16, 7802–7838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and Medicinal Uses of Terpenes. In Medicinal Plants: From Farm to Pharmacy; Joshee, N., Parajuli, P., Dhekney, S.A., Eds.; Springer: Cham, Switzerland, 2019; pp. 333–359. ISBN 9783030312695. [Google Scholar]
- Ludwiczuk, A.; Skalicka-Woźniak, K.; Georgiev, M.I. Terpenoids. In Pharmacognosy: Fundamentals, Applications and Strategy; Badal, S., Delgoda, R., Eds.; Academic Press Inc.: Cambridge, MA, USA, 2017; pp. 233–266. ISBN 9780128020999. [Google Scholar]
- Graßmann, J. Terpenoids as Plant Antioxidants. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press Inc.: Cambridge, MA, USA, 2005; Volume 72, pp. 505–535. ISBN 0127098720. [Google Scholar]
- Zorrilla, J.G.; Evidente, A. Structures and Biological Activities of Alkaloids Produced by Mushrooms, a Fungal Subgroup. Biomolecules 2022, 12, 1025. [Google Scholar] [CrossRef]
- Chang, S.-T. Overview of Mushroom Cultivation and Utilization as Functional Foods. In Mushrooms as Functional Foods; Cheung, P.C.K., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; pp. 1–33. ISBN 978-0-470-05406-2. [Google Scholar]
- Zeng, X.; Li, J.; Lyu, X.; Chen, X.M.; Guo, S. Nutritional Characterization and Untargeted Metabolomics of Oyster Mushroom Produced Using Astragalus membranaceus Var. mongolicus Stems Leaves Substrates. Front. Plant Sci. 2022, 13, 802801. [Google Scholar] [CrossRef]
- Sato, M.; Miyagi, A.; Yoneyama, S.; Gisusi, S.; Tokuji, Y.; Kawai-Yamada, M. CE–MS-Based Metabolomics Reveals the Metabolic Profile of Maitake Mushroom (Grifola frondosa) Strains with Different Cultivation Characteristics. Biosci. Biotechnol. Biochem. 2017, 81, 2314–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandohee, J.; Stevenson, P.G.; Conlan, X.A.; Zhou, X.R.; Jones, O.A.H. Off-Line Two-Dimensional Liquid Chromatography for Metabolomics: An Example Using Agaricus Bisporus Mushrooms Exposed to UV Irradiation. Metabolomics 2015, 11, 939–951. [Google Scholar] [CrossRef]
- Cai, Z.X.; Chen, M.Y.; Lu, Y.P.; Guo, Z.J.; Zeng, Z.H.; Liao, J.H.; Zeng, H. Metabolomics and Transcriptomics Unravel the Mechanism of Browning Resistance in Agaricus Bisporus. PLoS ONE 2022, 17, e0255765. [Google Scholar] [CrossRef]
- Yang, F.; Wang, H.; Feng, G.; Zhang, S.; Wang, J.; Cui, L. Rapid Identification of Chemical Constituents in Hericium Erinaceus Based on LC-MS/MS Metabolomics. J. Food Qual. 2021, 2021, 5560626. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.; Wu, D.; Zhang, Z.; Yang, Y.; Zhang, J. Understanding the Promotion of Heat Treatment on the Flavor of Lentinula Edodes Using Metabolomics Integrated with Transcriptomics. Food Res. Int. 2022, 162, 112051. [Google Scholar] [CrossRef]
- Cruickshank-Quinn, C.; Quinn, K.D.; Powell, R.; Yang, Y.; Armstrong, M.; Mahaffey, S.; Reisdorph, R.; Reisdorph, N. Multi-Step Preparation Technique to Recover Multiple Metabolite Compound Classes for in-Depth and Informative Metabolomic Analysis. J. Vis. Exp. 2014, 89, e51670. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cruickshank-Quinn, C.; Armstrong, M.; Mahaffey, S.; Reisdorph, R.; Reisdorph, N. New Sample Preparation Approach for Mass Spectrometry-Based Profiling of Plasma Results in Improved Coverage of Metabolome. J. Chromatogr. A 2013, 1300, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Sutliff, A.K.; Saint-Cyr, M.; Hendricks, A.E.; Chen, S.S.; Doenges, K.A.; Quinn, K.; Westcott, J.; Tang, M.; Borengasser, S.J.; Reisdorph, R.M.; et al. Lipidomics-Based Comparison of Molecular Compositions of Green, Yellow, and Red Bell Peppers. Metabolites 2021, 11, 241. [Google Scholar] [CrossRef] [PubMed]
- Reisdorph, N.A.; Hendricks, A.E.; Tang, M.; Doenges, K.A.; Reisdorph, R.M.; Tooker, B.C.; Quinn, K.; Borengasser, S.J.; Nkrumah-Elie, Y.; Frank, D.N.; et al. Nutrimetabolomics Reveals Food-Specific Compounds in Urine of Adults Consuming a DASH-Style Diet. Sci. Rep. 2020, 10, 1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisdorph, N.; Reisdorph, R.; Quinn, K.; Doenges, K. Metabolomics Mass Spectrometry Data Processing: Applications in Food Analysis. In Comprehensive Foodomics; Cifuentes, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 339–352. ISBN 9780128163962. [Google Scholar]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.; Bivens, A. Methods for the Analysis of Underivatized Amino Acids by LC/MS. In Agilent Application Note; Agilent Technologies, Inc.: Santa Clara, CA, USA, 2017. [Google Scholar]
- Huang, Y.; Li, W.; Keller, A.; Anumol, T.; Bivens, A. Quantitative Analysis of Underivatized Amino Acids in Plant Matrix by Hydrophilic Interaction Chromatography (HILIC) with LC/MS Detection. In Agilent Application Note; Agilent Technologies, Inc.: Santa Clara, CA, USA, 2018. [Google Scholar]
- Nowak, R.; Nowacka-Jechalke, N.; Pietrzak, W.; Gawlik-Dziki, U. A New Look at Edible and Medicinal Mushrooms as a Source of Ergosterol and Ergosterol Peroxide-UHPLC-MS/MS Analysis. Food Chem. 2022, 369, 130927. [Google Scholar] [CrossRef]
- Krzyczkowski, W.; Malinowska, E.; Suchocki, P.; Kleps, J.; Olejnik, M.; Herold, F. Isolation and Quantitative Determination of Ergosterol Peroxide in Various Edible Mushroom Species. Food Chem. 2009, 113, 351–355. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture (USDA) Agricultural Research Service. FoodData Central: Foundation Foods. Lion’s Mane Mushrooms. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/1999626/nutrients (accessed on 7 January 2022).
- U.S. Department of Agriculture (USDA) Agricultural Research Service. FoodData Central: Foundation Foods. Oyster Mushrooms. Available online: https://fdc.nal.usda.gov/fdc-app.html#/food-details/1999627/nutrients (accessed on 31 July 2022).
- Merdivan, S.; Lindequist, U. Ergosterol Peroxide: A Mushroom-Derived Compound with Promising Biological Activities—A Review. Int. J. Med. Mushrooms 2017, 19, 93–105. [Google Scholar] [CrossRef]
- Oshsawa, T.; Yukawa, M.; Takao, C.; Murayama, M.; Bando, H. Studies on Constituents of Fruit Body of Polyporus Umbellatus and Their Cytotoxic Activity. Chem. Pharm. Bull. 1992, 40, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Sárközy, A.; Béni, Z.; Dékány, M.; Zomborszki, Z.P.; Rudolf, K.; Papp, V.; Hohmann, J.; Ványolós, A. Cerebrosides and Steroids from the Edible Mushroom Meripilus Giganteus with Antioxidant Potential. Molecules 2020, 25, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, J.; Zhang, X.; Yu, W.; Wang, R.; Xue, Z.; Kou, X. Identification and Evaluation of Bioactivity of Compounds from the Mushroom Pleurotus Nebrodensis (Agaricomycetes) against Breast Cancer. Int. J. Med. Mushrooms 2017, 19, 839–848. [Google Scholar] [CrossRef]
- Wen, X.Y.; Jing, P. Dietary Cerebrosides in Seven Edible Mushrooms: One Step Detection, Quantification, and Si-SPE Assisted Isolation. J. Food Compos. Anal. 2022, 108, 104452. [Google Scholar] [CrossRef]
- Kumar, K.; Mehra, R.; Guiné, R.P.F.; Lima, M.J.; Kumar, N.; Kaushik, R.; Ahmed, N.; Yadav, A.N.; Kumar, H. Edible Mushrooms: A Comprehensive Review on Bioactive Compounds with Health Benefits and Processing Aspects. Foods 2021, 10, 2996. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, C.E.; Klein, M.R.S.T. Phytosterols in the Treatment of Hypercholesterolemia and Prevention of Cardiovascular Diseases. Arq. Bras. Cardiol. 2017, 109, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Hamada, Y.; Yasumoto, T.; Hashino, Y.; Masuyama, A.; Nagai, K. Total Syntheses and Endoplasmic Reticulum Stress Suppressive Activities of Hericenes A−C and Their Derivatives. Tetrahedron Lett. 2018, 59, 1733–1736. [Google Scholar] [CrossRef]
- Kobayashi, S.; Tamura, T.; Koshishiba, M.; Yasumoto, T.; Shimizu, S.; Kintaka, T.; Nagai, K. Total Synthesis, Structure Revision, and Neuroprotective Effect of Hericenones C-H and Their Derivatives. J. Org. Chem. 2021, 86, 2602–2620. [Google Scholar] [CrossRef]
- Ryu, S.H.; Hong, S.M.; Khan, Z.; Lee, S.K.; Vishwanath, M.; Turk, A.; Yeon, S.W.; Jo, Y.H.; Lee, D.H.; Lee, J.K.; et al. Neurotrophic Isoindolinones from the Fruiting Bodies of Hericium Erinaceus. Bioorg Med. Chem. Lett. 2021, 31, 127714. [Google Scholar] [CrossRef]
- Li, W.; Zhou, W.; Kim, E.J.; Shim, S.H.; Kang, H.K.; Kim, Y.H. Isolation and Identification of Aromatic Compounds in Lion’s Mane Mushroom and Their Anticancer Activities. Food Chem. 2015, 170, 336–342. [Google Scholar] [CrossRef]
- Chong, P.S.; Poon, C.H.; Roy, J.; Tsui, K.C.; Lew, S.Y.; Phang, M.W.L.; Tan, R.J.Y.; Cheng, P.G.; Fung, M.L.; Wong, K.H.; et al. Neurogenesis-Dependent Antidepressant-like Activity of Hericium Erinaceus in an Animal Model of Depression. Chin. Med. 2021, 16, 1–24. [Google Scholar] [CrossRef]
- González, A.; Cruz, M.; Losoya, C.; Nobre, C.; Loredo, A.; Rodríguez, R.; Contreras, J.; Belmares, R. Edible Mushrooms as a Novel Protein Source for Functional Foods. Food Funct. 2020, 11, 7400–7414. [Google Scholar] [CrossRef] [PubMed]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A Rich Source of the Antioxidants Ergothioneine and Glutathione. Food Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Agriculture and U.S. Department of Health and Human Services Dietary Guidelines for Americans, 2020–2025. Available online: https://www.dietaryguidelines.gov/ (accessed on 17 October 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 |  |  |  |  |  |  | |
| White Button | Crimini | Portabella | Lion’s Mane | Maitake | Oyster | Shiitake | |
| A. bisporus | A. bisporus | A. bisporus | H. erinaceus | G. frondose | P. ostreatus | L. edodes | |
| Farm 1: | A | A | A | A | A | A | A |
| Farm 2: | B | B | B | C | C | C | C |
| Reps/Farm: | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Total Reps: | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
| Mushroom Variety | Crimini | Lion’s Mane | Maitake | Oyster | Portabella | Shiitake | White Button |
|---|---|---|---|---|---|---|---|
| Crimini | 5690 * | 2327 | 2487 | 2485 | 255 | 1974 | 285 |
| Lion’s Mane | 3363 | 5690 | 2312 | 2176 | 2398 | 2086 | 2202 |
| Maitake | 3203 | 3378 | 5690 | 1969 | 2571 | 2126 | 2313 |
| Oyster | 3205 | 3514 | 3721 | 5690 | 2547 | 2096 | 2349 |
| Portabella | 5435 | 3292 | 3119 | 3143 | 5690 | 2096 | 411 |
| Shiitake | 3716 | 3604 | 3564 | 3594 | 3594 | 5690 | 1856 |
| White button | 5405 | 3488 | 3377 | 3341 | 5279 | 3834 | 5690 |
| Mushroom Variety | Crimini | Lion’s Mane | Maitake | Oyster | Portabella | Shiitake | White Button |
|---|---|---|---|---|---|---|---|
| Crimini | 549 * | 212 | 216 | 194 | 49 | 212 | 22 |
| Lion’s Mane | 337 | 549 | 200 | 175 | 232 | 194 | 223 |
| Maitake | 333 | 349 | 549 | 177 | 247 | 190 | 224 |
| Oyster | 355 | 374 | 372 | 549 | 224 | 185 | 189 |
| Portabella | 500 | 317 | 302 | 325 | 549 | 220 | 38 |
| Shiitake | 337 | 355 | 359 | 364 | 329 | 549 | 194 |
| White button | 527 | 326 | 325 | 360 | 511 | 355 | 549 |
| Mushroom Variety | Total Compounds (Annotated) * | Previously Determined to be in that Food 1 | Probably/ Possibly in that Food 2 | Found in Some/Any Other Food | Natural Product 3 | Other 4 | Cannot Determine 5 |
|---|---|---|---|---|---|---|---|
| Detected in all 7 | 520 | 0 | 113 | 259 | 11 | 18 | 119 |
| UNIQUE TO: | |||||||
| White button | 13 | 0 | 5 | 4 | 0 | 2 | 2 |
| Crimini | 4 | 0 | 0 | 1 | 0 | 1 | 2 |
| Portabella | 13 | 0 | 1 | 5 | 0 | 2 | 5 |
| Lion’s mane | 211 | 11 | 20 | 60 | 7 | 25 | 88 |
| Maitake | 206 | 1 | 8 | 97 | 21 | 14 | 65 |
| Oyster | 126 | 8 | 4 | 56 | 10 | 9 | 39 |
| Shiitake | 126 | 8 | 9 | 46 | 0 | 3 | 60 |
| Sum | 1219 | 28 | 160 | 528 | 49 | 74 | 380 |
| Mushroom Variety | Categorization | Compound | Main Class | Subclass | Notes |
|---|---|---|---|---|---|
| White button | Probably/ possibly in that food | (3beta,5alpha,9alpha,22E,24R)-5,9-Epidioxy-3-hydroxyergosta-7,22-dien-6-one Esi + 13.774996 | Prenol Lipids | Sesquiterpenoids | HMDB: found in common and oyster mushrooms. Constituent of Hypsizygus marmoreus (bunashimeji) and Pleurotus ostreatus (oyster mushroom). |
| Ergosterol peroxide Esi + 13.684002 | Steroids and Steroid Derivatives | Ergostane Steroids | Ergosterol peroxide is a secondary metabolite commonly detected in different mushrooms [33,34]. | ||
| Oyster | Previously determined to be in that food | (3beta,5alpha,9alpha,22E,24R)-5,9-Epidioxy-3-hydroxyergosta-7,22-dien-6-one | Prenol Lipids | Sesquiterpenoids | HMDB: Constituent of Hypsizygus marmoreus (bunashimeji) and Pleurotus ostreatus (oyster mushroom). |
| 2-Acetoxy-3-geranylgeranyl-1,4-dihydroxybenzene | Prenol Lipids | Diterpenoids | HMDB: found in common and oyster mushrooms. FooDB: associated with common and oyster mushrooms. | ||
| Methyl (Z,Z)-10-hydroxy-2,8-decadiene-4,6-diynoate | Fatty Acyls | Fatty Alcohols | HMDB: found in common and oyster mushrooms. FooDB: associated with common and oyster mushrooms. | ||
| Probably/ possibly in that food | Polyporusterone E Esi + 0.83 | Steroids and Steroid Derivatives | Cholestane Steroids | HMDB: found in common and oyster mushrooms FooDB: associated with common and oyster mushrooms. | |
| All 7 varieties | Probably/ possibly in that food | Cerebroside B | Prenol Lipids | Triterpenoids | FoodDB: associated with common and oyster mushrooms. |
| N-(2R-Hydroxyhexadecanoyl)-2S-amino-9-methyl-4E,8E-octadecadiene-1,3R-diol Esi + 4.5509977 | Sphingolipids | Ceramides | HMDB: found in common and oyster mushrooms. FooDB: associated with common and oyster mushrooms. | ||
| (3beta,22E,24R)-Ergosta-4,6,8(14),22-tetraen-3-ol Esi + 15.40999 | Steroids and Steroid Derivatives | Ergostane Steroids | FoodDB: associated with common and oyster mushrooms. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uffelman, C.N.; Doenges, K.A.; Armstrong, M.L.; Quinn, K.; Reisdorph, R.M.; Tang, M.; Krebs, N.F.; Reisdorph, N.A.; Campbell, W.W. Metabolomics Profiling of White Button, Crimini, Portabella, Lion’s Mane, Maitake, Oyster, and Shiitake Mushrooms Using Untargeted Metabolomics and Targeted Amino Acid Analysis. Foods 2023, 12, 2985. https://doi.org/10.3390/foods12162985
Uffelman CN, Doenges KA, Armstrong ML, Quinn K, Reisdorph RM, Tang M, Krebs NF, Reisdorph NA, Campbell WW. Metabolomics Profiling of White Button, Crimini, Portabella, Lion’s Mane, Maitake, Oyster, and Shiitake Mushrooms Using Untargeted Metabolomics and Targeted Amino Acid Analysis. Foods. 2023; 12(16):2985. https://doi.org/10.3390/foods12162985
Chicago/Turabian StyleUffelman, Cassi N., Katrina A. Doenges, Michael L. Armstrong, Kevin Quinn, Richard M. Reisdorph, Minghua Tang, Nancy F. Krebs, Nichole A. Reisdorph, and Wayne W. Campbell. 2023. "Metabolomics Profiling of White Button, Crimini, Portabella, Lion’s Mane, Maitake, Oyster, and Shiitake Mushrooms Using Untargeted Metabolomics and Targeted Amino Acid Analysis" Foods 12, no. 16: 2985. https://doi.org/10.3390/foods12162985
APA StyleUffelman, C. N., Doenges, K. A., Armstrong, M. L., Quinn, K., Reisdorph, R. M., Tang, M., Krebs, N. F., Reisdorph, N. A., & Campbell, W. W. (2023). Metabolomics Profiling of White Button, Crimini, Portabella, Lion’s Mane, Maitake, Oyster, and Shiitake Mushrooms Using Untargeted Metabolomics and Targeted Amino Acid Analysis. Foods, 12(16), 2985. https://doi.org/10.3390/foods12162985