
High Antioxidant Capacity of Lacticaseibacillus paracasei TDM-2 and Pediococcus pentosaceus TCM-3 from Qinghai Tibetan Plateau and Their Function towards Gut Modulation

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Isolation of LAB
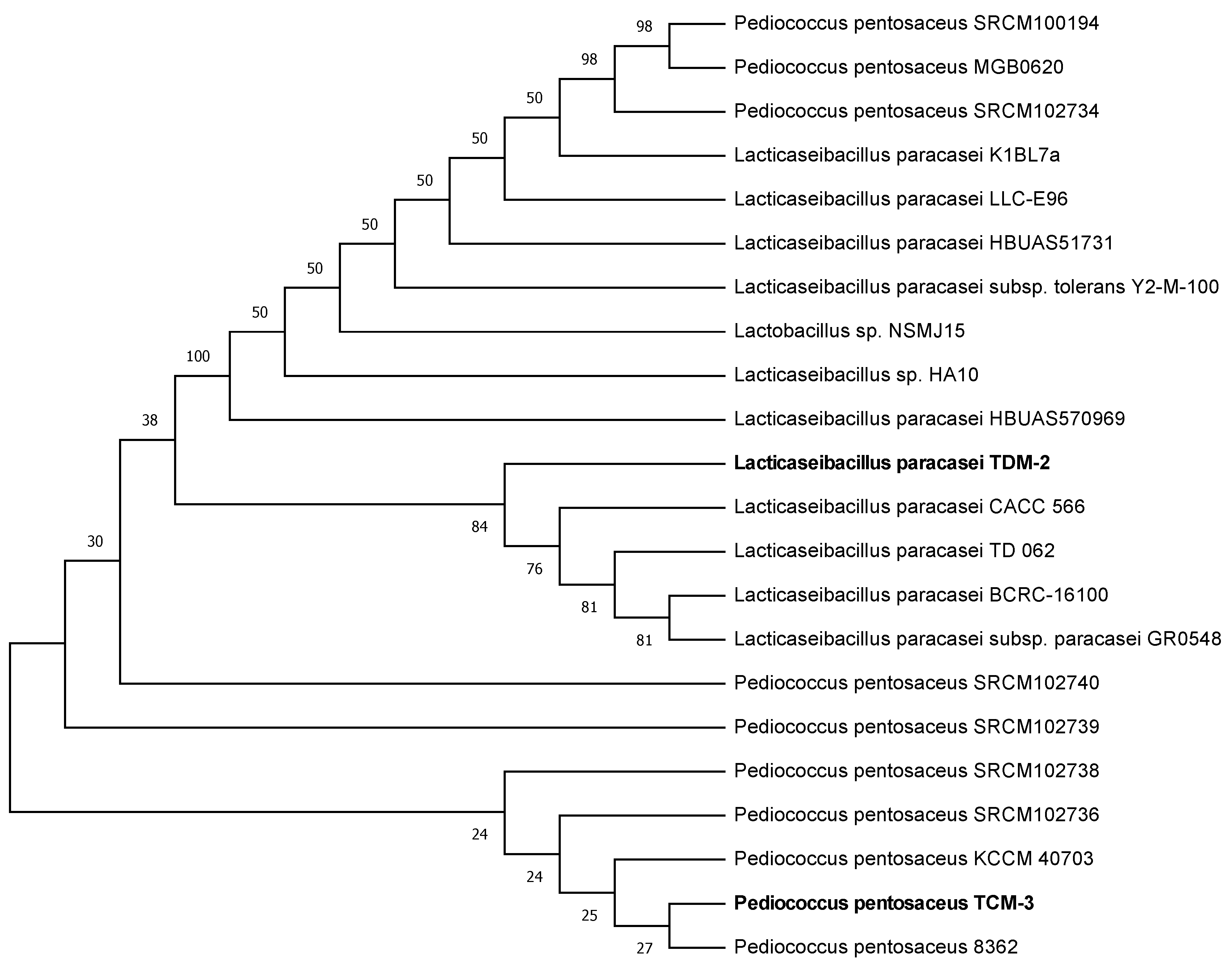
2.2. Identification of LAB
2.3. Antioxidant Activity Screening of Probiotics
2.3.1. Preparation of Cell-Free Extracts
2.3.2. H2O2-Resistant Ability
2.3.3. DPPH Radical Scavenging Activity
2.3.4. Superoxide Anion Radical (O2−) Scavenging Activity
2.3.5. Hydroxyl Radical (HO•) Scavenging Activity
2.3.6. Reductive Activity
2.3.7. Total Antioxidant Capacity (T-AOC)
2.4. In Vitro Probiotics Characteristic Determination
2.5. In Vitro Fermentation and Microbial Analysis
3. Result
3.1. Higher Antioxidant of P. pentosaceus TCM-3 and L. paracasei TDM-2 Compared with the Reference LAB
3.2. Resistance to Acid and Bile Salt under Gastric and Intestinal Conditions
3.3. Auto-Aggregation and Cell Surface Hydrophobicity
3.4. Susceptibility to Antibiotics
3.5. Addition of TCM-3 or TDM-2 Significantly Alters Microbiota Structure In Vitro
3.6. Pediococcus Is Positively Correlated with SCFA Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sidhu, M.; Poorten, D.V.D. The gut microbiome. Aust. Fam. Physician 2017, 46, 206–211. [Google Scholar] [PubMed]
- Liu, K.; Chen, S.; Huang, J.; Ren, F.; Yang, T.; Long, D.; Li, H.; Huang, X. Oral microbiota of children is conserved across Han, Tibetan and Hui Groups and is correlated with diet and gut microbiota. Microorganisms 2021, 9, 1030. [Google Scholar] [CrossRef]
- Persichetti, E.; Michele, A.D.; Codini, M.; Traina, G. Antioxidative capacity of Lactobacillus fermentum LF31 evaluated in vitro by oxygen radical absorbance capacity assay. Nutrition 2014, 30, 936–938. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, D.; Cui, L.; Ying, C. Effect of viable Bifidobacterium supplement on the immune status and inflammatory response in patients undergoing resection for colorectal cancer. Zhonghua Wei Chang Wai Ke Za Zhi 2010, 13, 40–43. [Google Scholar]
- Wang, Y.; Wu, Y.; Wang, Y.; Fu, A.; Gong, L.; Li, W.; Li, Y. Bacillus amyloliquefaciens SC06 alleviates the oxidative stress of IPEC-1 via modulating Nrf2/Keap1 signaling pathway and decreasing ROS production. Appl. Microbiol. Biotechnol. 2017, 101, 3015–3026. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Shang, N.; Li, P. In vitro and in vivo antioxidant activity of Bifidobacterium animalis 01 isolated from centenarians. Curr. Microbiol. 2011, 62, 1097–1103. [Google Scholar] [CrossRef]
- Isolauri, E.; Mykkänen, H.; Ling, W.H.; Salminen, S. Oral bacteriotherapy for viral gastroenteritis. Dig. Dis. Sci. 1994, 39, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Ostrakhovitch, E.A. Oxidative stress in rheumatoid arthritis leukocytes: Suppression by rutin and other antioxidants and chelators. Biochem. Pharmacol. 2001, 62, 743–746. [Google Scholar] [CrossRef]
- Williams, N.T. Probiotics. Am. J. Health Syst. Pharm. 2010, 67, 449–458. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. J. Dairy Sci. 2017, 100, 6025–6041. [Google Scholar] [CrossRef]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Wang, H.; Liu, Z.; Liu, Y.; DeJi; Li, B.; Huang, X. Recent Perspective of Lactobacillus in Reducing Oxidative Stress to Prevent Disease. Antioxidants 2023, 12, 769. [Google Scholar] [CrossRef] [PubMed]
- Ahire, J.J.; Mokashe, N.U.; Patil, H.J.; Chaudhari, B.L. Antioxidative potential of folate producing probiotic Lactobacillus helveticus CD6. J. Food Sci. Technol. 2013, 50, 26–34. [Google Scholar] [CrossRef]
- Han, K.J.; Lee, J.E.; Lee, N.K.; Paik, H.D. Antioxidant and anti-Inflammatory effect of probiotic Lactobacillus plantarum KU15149 derived from Korean homemade diced-radish kimchi. J. Microbiol. Biotechnol. 2020, 30, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wu, R.; Xu, W.; Cocolin, L.; Liang, H.; Ji, C.; Zhang, S.; Chen, Y.; Lin, X. Combined effects of lipase and Lactiplantibacillus plantarum 1-24-LJ on physicochemical property, microbial succession and volatile compounds formation in fermented fish product. J. Sci. Food Agric. 2023, 103, 2304–2312. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Lin, X.; Liang, H.; Zhang, S.; Ji, C. Screening of Lactiplantibacillus plantarum with high stress tolerance and high esterase activity and their effect on promoting protein metabolism and flavor formation in suanzhayu, a Chinese fermented fish. Foods 2022, 11, 112851–112951. [Google Scholar]
- Kim, K.T.; Yang, S.J.; Paik, H.D. Probiotic properties of novel probiotic Levilactobacillus brevis KU15147 isolated from radish kimchi and its antioxidant and immune-enhancing activities. Food Sci. Biotechnol. 2021, 30, 257–265. [Google Scholar] [CrossRef]
- Youssef, M.; Ahmed, H.Y.; Zongo, A.; Korin, A.; Zhan, F.; Hady, E.; Umair, M.; Shahid Riaz Rajoka, M.; Xiong, Y.; Li, B. Probiotic supplements: Their strategies in the therapeutic and prophylactic of human fife-threatening diseases. Int. J. Mol. Sci. 2021, 22, 11290. [Google Scholar] [CrossRef]
- Kim, J.C.K.; Kim, J.S.; Jung, H.C.; Kim, B.; Park, M.S.; Ji, G.E.; Cho, J.Y.; Hong, K.S. Probiotic treatment induced change of inflammation related metabolites in IBS-D patients/double-blind, randomized, placebo-controlled trial. Food Sci. Biotechnol. 2020, 29, 837–844. [Google Scholar] [CrossRef]
- Shi, L.; Pan, R.; Lin, G.; Liang, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. Lactic acid bacteria alleviate liver damage caused by perfluorooctanoic acid exposure via antioxidant capacity, biosorption capacity and gut microbiota regulation. Ecotoxicol. Environ. Saf. 2021, 222, 112515. [Google Scholar] [CrossRef]
- Li, W.; Guo, L.; Huang, W.; Ma, Y.; Muhammad, I.; Hanif, A.; Ding, Z.; Guo, X. Antioxidant properties of lactic acid bacteria isolated from traditional fermented yak milk and their probiotic effects on the oxidative senescence of Caenorhabditis elegans. Food Funct. 2022, 13, 3690–3703. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eftekharzadeh, B.; Maghsoudi, N.; Khodagholi, F. Stabilization of transcription factor Nrf2 by tBHQ prevents oxidative stress-induced amyloid β formation in NT2N neurons. Biochimie 2010, 92, 245–253. [Google Scholar] [CrossRef]
- Griendling, K.K.; FitzGerald, G.A. Oxidative stress and cardiovascular injury: Part I: Basic mechanisms and in vivo monitoring of ROS. Circulation 2003, 108, 1912–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Tsao, S.P.; Nurrahma, B.A.; Kumar, R.; Wu, C.H.; Yeh, T.H.; Chiu, C.C.; Lee, Y.P.; Liao, Y.C.; Huang, C.H.; Yeh, Y.T. Probiotic Enhancement of Antioxidant Capacity and Alterations of Gut Microbiota Composition in 6-Hydroxydopamin-Induced Parkinson’s Disease Rats. Antioxidants 2021, 10, 1823. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.H.; Zheng, W.X.; Zhao, Z.T.; Meng, N.; Zhang, Q.R.; Zhu, W.J.; Li, R.D.; Liang, X.L.; Li, Q.Y. A Pilot Study of the Effect of Lactobacillus casei Obtained from Long-Lived Elderly on Blood Biochemical, Oxidative, and Inflammatory Markers, and on Gut Microbiota in Young Volunteers. Nutrients 2021, 13, 3891. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Zhang, T.; Rong, Z.; Hu, L.; Gu, Z.; Wu, Q.; Dong, S.; Liu, Q.; Lin, Z.; Deutschova, L.; et al. Population transcriptomes reveal synergistic responses of DNA polymorphism and RNA expression to extreme environments on the Qinghai-Tibetan Plateau in a predatory bird. Mol. Ecol. 2017, 26, 2993–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Jin, M.; Fei, X.; Yuan, Z.; Wang, Y.; Quan, K.; Wang, T.; Yang, J.; He, M.; Wei, C. Transcriptome Comparison Reveals the Difference in Liver Fat Metabolism between Different Sheep Breeds. Animals 2022, 12, 1650. [Google Scholar] [CrossRef]
- Ai, H.; Yang, B.; Li, J.; Xie, X.; Chen, H.; Ren, J. Population history and genomic signatures for high-altitude adaptation in Tibetan pigs. BMC Genom. 2014, 15, 834–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Fan, Z.; Han, E.; Hou, R.; Zhang, L.; Galaverni, M.; Huang, J.; Liu, H.; Silva, P.; Li, P.; et al. Hypoxia adaptations in the grey wolf (Canis lupus chanco) from Qinghai-Tibet Plateau. PLoS Genet. 2014, 10, e1004466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M.; et al. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, R.L.; Cai, Q.; Shen, Y.Y.; San, A.; Ma, L.; Zhang, Y.; Yi, X.; Chen, Y.; Yang, L.; Huang, Y.; et al. Draft genome sequence of the Tibetan antelope. Nat. Commun. 2013, 4, 1858–1865. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Sun, J.; Liu, B.; Wang, J.; Zeng, T. Cushion plants as critical pioneers and engineers in alpine ecosystems across the Tibetan Plateau. Ecol. Evol. 2021, 11, 11554–11558. [Google Scholar] [CrossRef]
- Zheng, W.; Yu, S.; Zhang, W.; Zhang, S.; Fu, J.; Ying, H.; Pingcuo, G.; Liu, S.; Zhao, F.; Wu, Q.; et al. The content and diversity of carotenoids associated with high-altitude adaptation in Tibetan peach fruit. Food Chem. 2023, 398, 133909. [Google Scholar] [CrossRef]
- Wu, H.; Gu, Q.; Xie, Y.; Lou, Z.; Xue, P.; Fang, L.; Yu, C.; Jia, D.; Huang, G.; Zhu, B.; et al. Cold-adapted Bacilli isolated from the Qinghai-Tibetan Plateau are able to promote plant growth in extreme environments. Environ. Microbiol. 2019, 21, 3505–3526. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Liu, G.; Chen, T.; Li, S.; An, L.; Zhang, G.; Li, G.; Chang, S.; Zhang, W.; Chen, X.; et al. Characterization of the genome of a Nocardia strain isolated from soils in the Qinghai-Tibetan Plateau that specifically degrades crude oil and of this biodegradation. Genomics 2019, 111, 356–366. [Google Scholar] [CrossRef]
- Miao, Y.Z.; Xu, H.; Fei, B.J.; Qiao, D.R.; Cao, Y. PCR-RFLP analysis of the diversity of phytate-degrading bacteria in the Tibetan Plateau. Can. J. Microbiol. 2013, 59, 245–251. [Google Scholar] [CrossRef]
- Navani, N.K.; Ghosh, T. Indian Provisional. Patent No. 201811019299, 26 April 2018. [Google Scholar]
- Kozaki, M.; Uchimura, T.; Okada, S. Experimental Manual for Lactic Acid Bacteria; Asakurasyoten: Tokyo, Japan, 1992; pp. 29–72. [Google Scholar]
- Adimpong, D.B.; Nielsen, D.S.; Sorensen, K.I.; Derkx, P.M.F.; Jespersen, L. Genotypic characterization and safety assessment of lactic acid bacteria from indigenous African fermented food products. BMC Microbiol. 2012, 12, 12–75. [Google Scholar] [CrossRef] [Green Version]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef]
- Lin, M.Y.; Chang, F.J. Antioxidative effect of intestinal bacteria Bifidobacterium longum ATCC 15708 and Lactobacillus acidophilus ATCC 4356. Digest. Dis. Sci. 2000, 45, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Leite, A.M.; Miguel, M.A.; Peixoto, R.S.; Ruas-Madiedo, P.; Paschoalin, V.M.; Mayo, B.; Delgado, S. Probiotic potential of selected lactic acid bacteria strains isolated from Brazilian kefir grains. J. Dairy Sci. 2015, 98, 3622–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Liu, T.; Radji, C.A.; Yang, J.; Chen, L. Isolation, identification, and evaluation of new lactic acid bacteria strains with both cellular antioxidant and bile salt hydrolase activities in vitro. J. Food Prot. 2016, 79, 1919–1928. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, G. The derivatization and antioxidant activities of yeast mannan. Int. J. Biol. Macromol. 2018, 107, 755–761. [Google Scholar] [CrossRef]
- He, H.L.; Cao, C.H.; Cui, Z.W. Photometric determination of hydroxyl free radical in Fenton system by brilliant green. Chin. J. Health Lab. Technol. 2004, 6, 236–237. [Google Scholar]
- Lin, M.Y.; Yen, C.Y. Antioxidative ability of lactic acid bacteria. J. Agric. Food Chem. 1999, 47, 1460–1466. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Xing, Z.; Li, C.; Wang, J.; Wang, Y. Molecular mechanisms and in vitro antioxidant effects of Lactobacillus plantarum MA2. Food Chem. 2017, 221, 1642–1649. [Google Scholar] [CrossRef]
- Guo, C.F.; Zhang, L.W.; Han, X.; Yi, H.X.; Li, J.Y.; Tuo, Y.F.; Zhang, Y.C.; Du, M.; Shan, Y.J.; Yang, L. Screening for cholesterol-lowering probiotic based on deoxycholic acid removal pathway and studying its functional mechanisms in vitro. Anaerobe 2012, 18, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.; Tsakalidou, E.; Nychas, G.J.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Sieladie, D.V.; Zombou, N.F.; Kaktcham, P.M.; Cresci, A.; Fontech, F. Probiotic properties of lactobacilli strains isolated from raw cow milk in the western highlands of Cameroon. Food Biotechnol. 2011, 19, 12–28. [Google Scholar]
- Del, R.B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, auto-aggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Isolauri, E.; Salminen, S. Specific probiotic strains and their combinations counteract adhesion of Enterobacter sakazakii to intestinal mucus. FEMS Microbiol. Lett. 2008, 285, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to 543 hydrocarbons: A simple method for measuring cell surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Crow, V.L.; Gopal, P.K. Cell surface differences of lactococcal strains. Int. Dairy J. 1895, 5, 45–68. [Google Scholar] [CrossRef]
- Bellon-Fontaine, M.N.; Rault, J.; Oss, C.J.V. Microbial adhesion to solvents: A novel method to determine the electron-donor/electron-acceptor or Lewis acid-base properties of microbial-cells. Colloids Surf. 1996, 7, 47–53. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement. 2012. Available online: https://m.ibric.org/miniboard/down.php?Board=exp_qna&filename=CLSI%20-%20M100%20S22E.pdf&id=531983&fidx=1 (accessed on 22 February 2023).
- Haller, D.; Colbus, H.; Ganzle, M.G.; Scherenbacher, P.; Bode, C.; Hammes, W.P. Metabolic and functional properties of lactic acid bacteria in the gastro-intestinal ecosystem: A comparative in vitro study between bacteria of intestinal and fermented food origin. Syst. Appl. Microbiol. 2001, 24, 218–226. [Google Scholar] [CrossRef]
- Liu, H.; Gong, J.; Chabot, D.; Miller, S.S.; Cui, S.W.; Ma, J.; Zhong, F.; Wang, Q. Incorporation of polysaccharides into sodium caseinate-low melting point fat microparticles improves probiotic bacterial survival during simulated gastrointestinal digestion and storage. Food Hydrocoll. 2016, 54, 328–337. [Google Scholar] [CrossRef]
- Zhao, Y.; Bi, J.; Yi, J.; Wu, X.; Ma, Y.; Li, R. Pectin and homogalacturonan with small molecular mass modulate microbial community and generate high SCFAs via in vitro gut fermentation. Carbohydr. Polym. 2021, 269, 118326. [Google Scholar] [CrossRef]
- Yu, L.; Duan, H.; Kellingray, L.; Cen, S.; Tian, F.; Zhao, J.; Zhang, H.; Gall, G.L.; Mayer, M.J.; Zhai, Q.; et al. Lactobacillus plantarum-mediated regulation of dietary aluminum induces changes in the human gut microbiota: An in vitro colonic fermentation study. Probiotics Antimicrob. Proteins 2021, 13, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Henrique-Bana, F.C.; Wang, X.; Costa, G.N.; Spinosa, W.A.; Miglioranza, L.H.S.; Scorletti, E.; Calder, P.C.; Byrne, C.D.; Gibson, G.R. In vitro effects of Bifidobacterium lactis-based synbiotics on human faecal bacteria. Food Res. Int. 2020, 128, 108776. [Google Scholar] [CrossRef]
- Erwin, E.S.; Marco, G.J.; Emery, E.M. Volatile Fatty Acid Analyses of Blood and Rumen Fluid by Gas Chromatography. J. Dairy Sci. 1961, 44, 1768–1771. [Google Scholar] [CrossRef]
- Jones, R.M.; Neish, A.S. Redox signaling mediated by the gut microbiota. Free Radic. Biol. Med. 2017, 105, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Nardone, G.; Compare, D.; Liguori, E.; Di Mauro, V.; Rocco, A.; Barone, M.; Napoli, A.; Lapi, D.; Iovene, M.R.; Colantuoni, A. Protective effects of Lactobacillus paracasei F19 in a rat model of oxidative and metabolic hepatic injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G669–G676. [Google Scholar] [CrossRef] [Green Version]
- Mikelsaar, M.; Zilmer, M. Lactobacillus fermentum ME-3—An antimicrobial and antioxidative probiotic. Microb. Ecol. Health Dis. 2009, 21, 1–27. [Google Scholar] [PubMed] [Green Version]
- Gao, Y.; Li, D.; Liu, S.; Liu, Y. Probiotic potential of L. sake C2 isolated from traditional Chinese fermented cabbage. Eur. Food Res. Technol. 2011, 234, 45–51. [Google Scholar] [CrossRef]
- Prasad, J.; Gill, H.; Smart, J.; Gopal, P.K. Selection and Characterisation of Lactobacillus and Bifidobacterium Strains for Use as Probiotics. Int. Dairy J. 1998, 8, 993–1002. [Google Scholar] [CrossRef]
- Tokatli, M.; Gulgor, G.; Bagder Elmaci, S.; Arslankoz Isleyen, N.; Ozcelik, F. In vitro properties of potential probiotic indigenous lactic acid bacteria originating from traditional pickles. Biomed Res. Int. 2015, 2015, 315819. [Google Scholar] [CrossRef] [Green Version]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N.; Floros, G.; Hatzikamari, M. Probiotic and technological properties of facultatively heterofermentative lactobacilli from Greek traditional cheeses. Food Biotechnol. 2013, 26, 85–105. [Google Scholar]
- Ahmadova, A.; Todorov, S.D.; Choiset, Y.; Rabesona, H.; Mirhadi, T.Z.; Kuliyev, A.; Franco, B.D.G.D.M.; Chobert, J.M.; Haertlé, T. Evaluation of antimicrobial activity, probiotic properties and safety of wild strain Enterococcus faecium AQ71 isolated from Azerbaijani Motal cheese. Food Control. 2013, 30, 631–641. [Google Scholar] [CrossRef]
- Miele, E.; Pascarella, F.; Giannetti, E.; Quaglietta, L.; Baldassano, R.N.; Staiano, A. Effect of a probiotic preparation (VSL#3) on induction and maintenance of remission in children with ulcerative colitis. Am. J. Gastroenterol. 2009, 104, 437–443. [Google Scholar] [PubMed]
- Sun, Y.; Zhang, S.; Nie, Q.; He, H.; Tan, H.; Geng, F.; Ji, H.; Hu, J.; Nie, S. Gut firmicutes: Relationship with dietary fiber and role in host homeostasis. Crit. Rev. Food Sci. Nutr. 2022, 1–16. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Strukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, S.; Wang, Q.; Guan, X.; Qian, L.; Li, J.; Zheng, Y.; Lin, B. Pediococcus pentosaceus B49 from human colostrum ameliorates constipation in mice. Food Funct. 2020, 11, 5607–5620. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, B.; Wang, Q.; Tang, L.; Zou, P.; Zeng, Z.; Zhang, H.; Gong, L.; Li, W. Protective Effects of Lactobacillus plantarum Lac16 on Clostridium perfringens Infection-Associated Injury in IPEC-J2 Cells. Int. J. Mol. Sci. 2021, 22, 12388. [Google Scholar] [CrossRef]
- Koh, A.; Vadder, F.D.; Kovatcheva-Datchary, P.; Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalile, B.; Oudenhove, L.V.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, F.; Xiao, F.; Li, X.; Li, Y.; Wang, X.; Yu, G.; Zhang, T.; Wang, Y. Pediococcus pentosaceus CECT 8330 protects DSS-induced colitis and regulates the intestinal microbiota and immune responses in mice. J. Transl. Med. 2022, 20, 33. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TDM-2 | TCM-3 | LGG | |
|---|---|---|---|
| DPPH free radical scavenging activity (%) | 84.15 ± 0.03 b | 87.99 ± 0.07 a | 84.13 ± 0.02 b |
| H2O2-resistant ability (%) | |||
| 0 mmol L−1 H2O2 | 77.70 ± 0.09 b | 89.19 ± 0.08 a | 67.40 ± 0.02 c |
| 1.0 mmol L−1 H2O2 | 73.43 ± 0.03 b | 84.92 ± 0.01 a | 55.40 ± 0.10 c |
| 1.5 mmol L−1 H2O2 | 72.74 ± 0.06 b | 79.70 ± 0.07 a | 53.96 ± 0.06 c |
| 2.0 mmol L−1 H2O2 | 63.65 ± 0.04 b | 71.79 ± 0.09 a | 53.17 ± 0.06 c |
| HO• scavenging activity (%) | 92.33 ± 0.09 b | 97.00 ± 0.07 a | 84.02 ± 0.06 b |
| O2− scavenging activity (%) | 17.17 ± 0.08 c | 37.59 ± 0.02 a | 26.92 ± 0.02 b |
| Reductive activity (%) | 29.48 ± 0.03 a | 13.38 ± 0.00 b | 10.20 ± 0.01 c |
| T-AOC (μmoL/mL) | 4.10 ± 0.17 a | 3.68 ± 0.08 b | 3.53 ± 0.02 b |
| TDM-2 | TCM-3 | LGG | ||
|---|---|---|---|---|
| Survival Rate of Acid after 3 h (%) | pH 3 | 104.88 ± 0.76 a | 97.26 ± 0.49 b | 104.48 ± 0.81 a |
| Survival Rate of Bile after 4 h (%) | pH 8 | 121.19 ± 0.69 a | 107.46 ± 0.52 b | 108.22 ± 0.04 b |
| Gastric juice | 3 h | 113.31 ± 0.12 a | 97.98 ± 0.25 b | 101.28 ± 1.10 b |
| Intestinal juice | 3 h | 118.57 ± 0.78 a | 108.38 ± 0.93 b | 103.93 ± 0.44 c |
| 6 h | 108.33 ± 1.21 a | 103.27 ± 0.27 a | 94.48 ± 0.77 b | |
| 24 h | 94.79 ± 0.10 b | 96.20 ± 0.18 a | 87.05 ± 0.17 b | |
| Auto-aggregation (%) | 0 h | 64.72 ± 0.28 b | 68.75 ± 0.08 c | 66.98 ± 0.13 a |
| 2 h | 86.26 ± 0.14 a | 72.26 ± 0.01 c | 70.67 ± 1.43 b | |
| 4 h | 93.08 ± 0.07 a | 77.72 ± 0.01 c | 81.88 ± 0.03 b | |
| 6 h | 93.55 ± 0.01 a | 87.12 ± 0.04 c | 86.01 ± 0.01 b | |
| Hydrophobicity (%) | 86.84 ± 0.21 a | 30.82 ± 0.07 b | 26.03 ± 0.10 b |
| TDM-2 | TCM-3 | |
|---|---|---|
| Antibiotic Susceptibility | ||
| VAN | S | S |
| AM | S | S |
| CN | S | S |
| CLI | S | S |
| ERY | S | S |
| TE | S | S |
| AMC | S | S |
| CPL | S | S |
| KAN | S | R |
| SM | R | S |
| SCFAs | Time of Incubation (h) | TCM-3 (mmol/L) | TDM-2 (mmol/L) | Control (mmol/L) |
|---|---|---|---|---|
| Acetic acid | 0 h | 3.18 ± 0.22 a | 2.71 ± 0.04 b | 2.83 ± 0.05 b |
| 6 h | 3.82 ± 0.01 a | 3.61 ± 0.01 b | 3.64 ± 0.04 b | |
| 12 h | 4.54 ± 0.09 a | 4.06 ± 0.03 b | 4.00 ± 0.05 b | |
| 24 h | 4.99 ± 0.02 a | 4.90 ± 0.08 b | 4.82 ± 0.09 b | |
| Propanoic acid | 0 h | 0.88 ± 0.00 a | 0.88 ± 0.01 a | 0.88 ± 0.01 a |
| 6 h | 0.97 ± 0.00 a | 0.956 ± 0.00 b | 0.96 ± 0.01 b | |
| 12 h | 1.24 ± 0.02 a | 1.06 ± 0.03 b | 1.05 ± 0.02 b | |
| 24 h | 1.35 ± 0.01 a | 1.24 ± 0.02 b | 1.22 ± 0.01 b | |
| Butyric acid | 0 h | 0.50 ± 0.00 a | 0.05 ± 0.00 a | 0.049 ± 0.00 a |
| 6 h | 0.49 ± 0.00 a | 0.05 ± 0.00 a | 0.049 ± 0.00 a | |
| 12 h | 0.49 ± 0.00 a | 0.05 ± 0.00 b | 0.049 ± 0.00 b | |
| 24 h | 0.05 ± 0.00 a | 0.05 ± 0.00 b | 0.049 ± 0.00 b | |
| Valeric acid | 0 h | 0.10 ± 0.00 a | 0.10 ± 0.00 a | 0.10 ± 0.00 a |
| 6 h | 0.11 ± 0.01 a | 0.12 ± 0.01 a | 0.12 ± 0.01 a | |
| 12 h | 0.11 ± 0.01 a | 0.11 ± 0.01 a | 0.11 ± 0.01 a | |
| 24 h | 0.10 ± 0.00 a | 0.11 ± 0.01 a | 0.11 ± 0.01 a | |
| Isobutyric acid | 0 h | 0.40 ± 0.00 a | 0.40 ± 0.00 a | 0.40 ± 0.00 a |
| 6 h | 0.40 ± 0.00 a | 0.40 ± 0.00 a | 0.40 ± 0.00 a | |
| 12 h | 0.40 ± 0.00 a | 0.40 ± 0.00 b | 0.40 ± 0.00 ab | |
| 24 h | 0.42 ± 0.00 a | 0.41 ± 0.01 a | 0.42 ± 0.01 a | |
| Isovaleric acid | 0 h | 0.059 ± 0.00 a | 0.058 ± 0.00 a | 0.06 ± 0.00 a |
| 6 h | 0.057 ± 0.00 a | 0.058 ± 0.00 a | 0.06 ± 0.00 a | |
| 12 h | 0.058 ± 0.00 a | 0.058 ± 0.00 b | 0.06 ± 0.00 ab | |
| 24 h | 0.06 ± 0.00 a | 0.058 ± 0.00 b | 0.06 ± 0.00 b | |
| Total fatty acids | 0 h | 4.67 ± 0.22 a | 4.21 ± 0.04 b | 4.34 ± 0.07 b |
| 6 h | 5.41 ± 0.02 a | 5.20 ± 0.01 b | 5.22 ± 0.04 b | |
| 12 h | 6.40 ± 0.10 a | 5.74 ± 0.01 b | 5.68 ± 0.07 b | |
| 24 h | 6,97 ± 0.02 a | 6.67 ± 0.10 b | 6.68 ± 0.11 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Fan, X.; Li, D.; Zhao, T.; Wu, D.; Liu, Z.; Long, D.; Li, B.; Huang, X. High Antioxidant Capacity of Lacticaseibacillus paracasei TDM-2 and Pediococcus pentosaceus TCM-3 from Qinghai Tibetan Plateau and Their Function towards Gut Modulation. Foods 2023, 12, 1814. https://doi.org/10.3390/foods12091814
Yang T, Fan X, Li D, Zhao T, Wu D, Liu Z, Long D, Li B, Huang X. High Antioxidant Capacity of Lacticaseibacillus paracasei TDM-2 and Pediococcus pentosaceus TCM-3 from Qinghai Tibetan Plateau and Their Function towards Gut Modulation. Foods. 2023; 12(9):1814. https://doi.org/10.3390/foods12091814
Chicago/Turabian StyleYang, Tingyu, Xueni Fan, Diantong Li, Tingting Zhao, Dan Wu, Zhenjiang Liu, Danfeng Long, Bin Li, and Xiaodan Huang. 2023. "High Antioxidant Capacity of Lacticaseibacillus paracasei TDM-2 and Pediococcus pentosaceus TCM-3 from Qinghai Tibetan Plateau and Their Function towards Gut Modulation" Foods 12, no. 9: 1814. https://doi.org/10.3390/foods12091814
APA StyleYang, T., Fan, X., Li, D., Zhao, T., Wu, D., Liu, Z., Long, D., Li, B., & Huang, X. (2023). High Antioxidant Capacity of Lacticaseibacillus paracasei TDM-2 and Pediococcus pentosaceus TCM-3 from Qinghai Tibetan Plateau and Their Function towards Gut Modulation. Foods, 12(9), 1814. https://doi.org/10.3390/foods12091814