


Hunted Wild Boars in Sardinia: Prevalence, Antimicrobial Resistance and Genomic Analysis of Salmonella and Yersinia enterocolitica
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
- Mesenteric lymph nodes: at least 25 g of lymph nodes in the ileo-caecal regions were cut out with a sterile, disposable scalpel and collected in a sterile plastic bag (3M Health Care, Milan).
- Colon content: the colon was incised with a sterile, disposable scalpel and at least 25 g of its contents was collected in a sterile plastic bag.
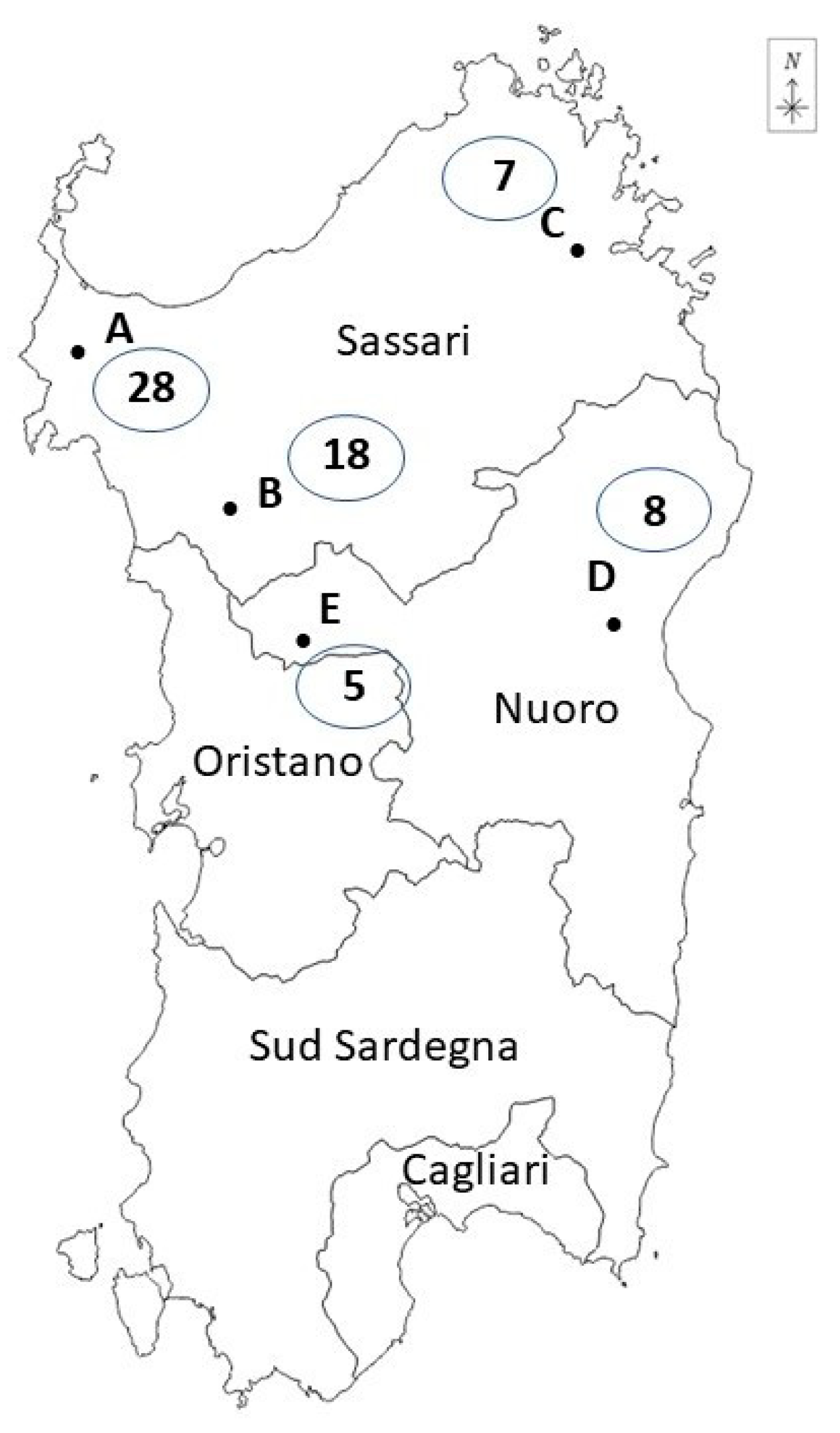
- Carcass surface: samples were taken after evisceration by means of a non-destructive method with a sterile sponge pre-moistened with 10 mL of sterile buffered peptone water (BPW, 3M Health Care, Milan) at the following points: ham, loins, abdomen and throat; these points were selected as they are indicated by European legislation for pig carcasses at the slaughterhouse (Reg. CE No. 2073/2005; ISO 17604:2015 [32]). Sampling was carried out using the same sponge for the four points, with a sterile 10 × 10 cm2 delimiter (Copan, Brescia, Italy), proceeding from the least contaminated point (ham) to the most contaminated (throat). The sponges were handled with a sterile glove and placed inside sterile sponge bags. All the samples were transported to the laboratory at +4 °C and processed within 24 h after collection. A total of 181 samples were collected, divided into 66 colon content samples, 66 mesenteric lymph node samples and 49 carcass surface samples. Carcass surface samples were collected from wild boars slaughtered in hunting houses A, B and C; in the other hunting houses (D, E) the skinning was conducted the day after harvesting, and it was therefore not possible to evaluate the surface contamination of the carcass.
2.2. Microbiological Analysis
2.3. Antimicrobial Susceptibility Testing
2.4. Whole Genome Sequencing
2.5. Statistical Analysis
3. Results
3.1. Prevalence of Salmonella and Y. enterocolitica
3.2. Antimicrobial Susceptibility Testing
3.3. Salmonella Characterization
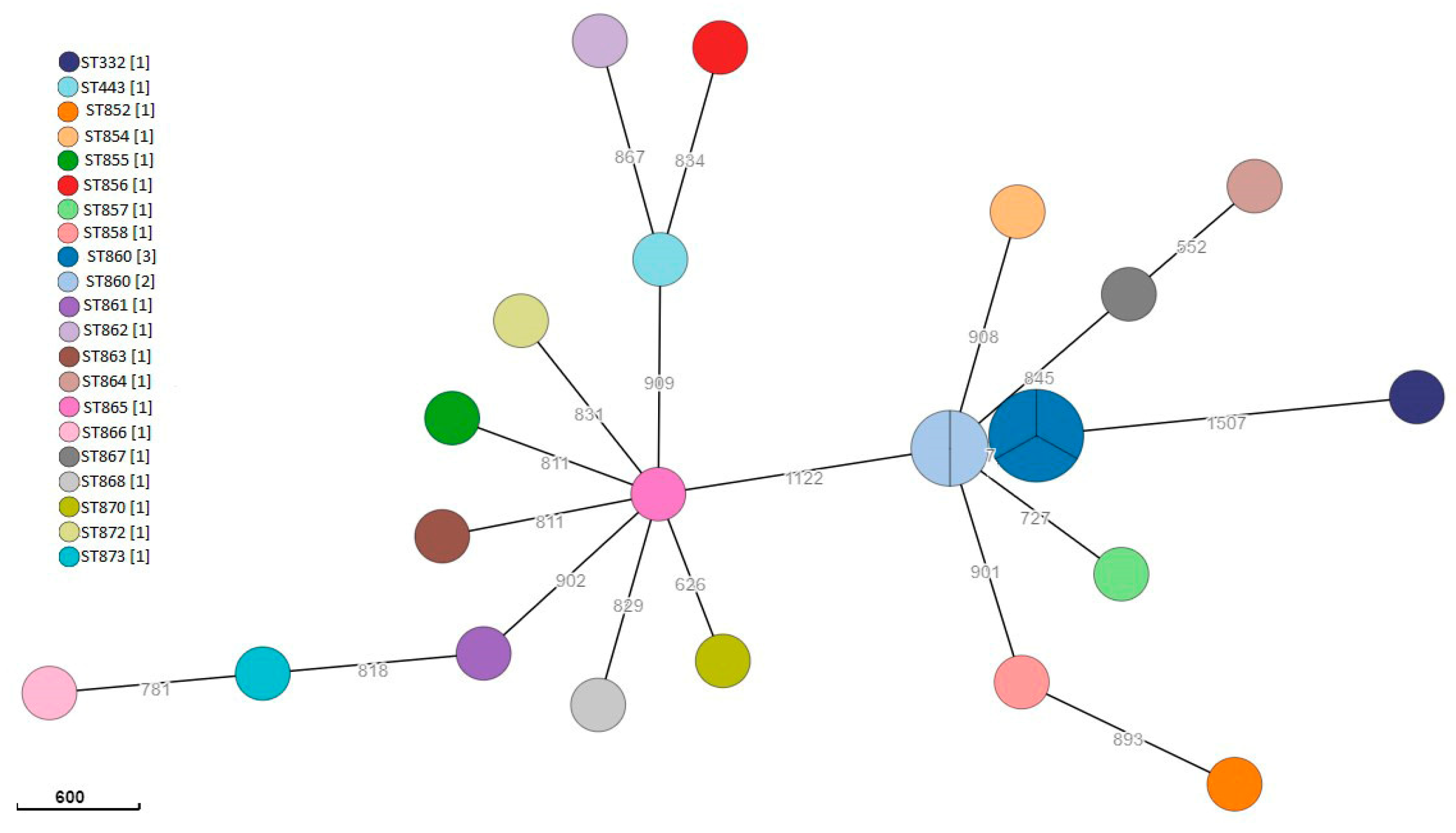
3.4. Yersinia enterocolitica Characterization
3.5. Bacterial Contamination of Carcasses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Powell, D.M. Pigs. In Grzimek’s Animal Life Encyclopedia, 2nd ed.; Kleiman, D.G., Geist, V., McDade, M., Eds.; Mammals IV; Michael Hutching, Ed.; Gale Research Inc.: Farmington Hills, MI, USA, 2003; Volume 15, pp. 275–290. [Google Scholar]
- Scandura, M.; Iacolina, L.; Cossu, A.; Apollonio, M. Effects of human perturbation on the genetic make-up of an island population: The case of the Sardinian Wild Boar. Heredity 2010, 106, 1012–1020. [Google Scholar] [CrossRef]
- Náhlik, A.; Cahill, S.; Cellina, S.; Gál, J.; Jánoska, F.; Rosell, C.; Massei, G. Wild Boar Management in Europe: Knowledge and Practice. In Ecology, Conservation and Management of Wild Pigs and Peccaries; Melletti, M., Meijaard, E., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 339–353. [Google Scholar] [CrossRef]
- Bieber, C.; Ruf, T. Population dynamics in wild boar Sus scrofa: Ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. J. Appl. Ecol. 2005, 42, 1203–1213. [Google Scholar] [CrossRef]
- Sales, J.; Kotrba, R. Meat from wild boar (Sus scrofa L.): A Review. Meat Sci. 2013, 94, 187–201. [Google Scholar] [CrossRef]
- Massei, G.; Kindberg, J.; Licoppe, A.; Gačić, D.; Šprem, N.; Kamler, J.; Baubet, E.; Hohmann, U.; Monaco, A.; Ozoliņš, J.; et al. Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag. Sci. 2015, 71, 492–500. [Google Scholar] [CrossRef]
- Lowe, S.M.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; The Invasive Species Specialist Group (ISSG) a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN): Auckland, New Zealand, 2000; Volume 12. [Google Scholar]
- Barrios-Garcia, M.N.; Ballari, S.A. Impact of wild boar (Sus scrofa) in its introduced and native range: A review. Biol. Invasions 2012, 14, 2283–2300. [Google Scholar] [CrossRef]
- Gortázar, C.; Ferroglio, E.; Höfle, U.; Frölich, K.; Vicente, J. Diseases shared between wildlife and livestock: A European perspective. Eur. J. Wildl. Res. 2007, 53, 241. [Google Scholar] [CrossRef]
- Risch, D.R.; Ringma, J.; Price, M.R. The global impact of wild pigs (Sus scrofa) on terrestrial biodiversity. Sci. Rep. 2021, 11, 13256. [Google Scholar] [CrossRef]
- Chiari, M.; Zanoni, M.; Tagliabue, S.; Lavazza, A.; Alborali, L.G. Salmonella serotypes in wild boars (Sus scrofa) hunted in northern Italy. Acta Vet. Scand. 2013, 55, 42. [Google Scholar] [CrossRef]
- Davidson, A.; Malkinson, D.; Shanas, U. Wild boar foraging and risk perception—Variation among urban, natural, and agricultural areas. J. Mammal. 2022, 103, 945–955. [Google Scholar] [CrossRef]
- Wacheck, S.; Fredriksson-Ahomaa, M.; König, M.; Stolle, A.; Stephan, R. Wild boars as an important reservoir for foodborne pathogens. Foodborne Pathog. Dis. 2010, 7, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, F.; Smulders, F.J.M.; Chopra-Dewasthaly, R.; Paulsen, P. Salmonella in the wildlife-human interface. Food Res. Int. 2012, 45, 603–608. [Google Scholar] [CrossRef]
- Brown, V.R.; Bowen, R.A.; Bosco-Lauth, A.M. Zoonotic pathogens from feral swine that pose a significant threat to public health. Transbound. Emerg. Dis. 2018, 65, 649–659. [Google Scholar] [CrossRef]
- Fredriksson-Ahomaa, M. Wild boar: A reservoir of Foodborne Zoonoses. Foodborne Pathog. Dis. 2019, 16, 153–165. [Google Scholar] [CrossRef]
- Branciari, R.; Ranucci, D. Editorial for the Special Issue: Game Meat and Game Meat Products: Safety, Quality and Consumer Perception. Foods 2022, 11, 2073. [Google Scholar] [CrossRef]
- Corradini, A.; Marescotti, M.E.; Demartini, E.; Gaviglio, A. Consumers’ perceptions and attitudes toward hunted wild game meat in the modern world: A literature review. Meat Sci. 2022, 194, 108955. [Google Scholar] [CrossRef]
- Gill, C.O. Microbiological conditions of meats from large game animals and birds. Meat Sci. 2007, 77, 149–160. [Google Scholar] [CrossRef]
- Gomes-Neves, E.; Abrantes, A.C.; Vieira-Pinto, M.; Müller, A. Wild game meat—A microbiological safety and hygiene challenge? Curr. Clin. Microbiol. Rep. 2021, 8, 31–39. [Google Scholar] [CrossRef]
- Giuggioli, G.; Olivastri, A.; Pennisi, L.; Paludi, D.; Ianieri, A.; Vergara, A. The hygiene-sanitary control in the wild game meats. Ital. J. Food Saf. 2018, 6, 222–224. [Google Scholar] [CrossRef]
- Ranucci, D.; Roila, R.; Onofri, A.; Cambiotti, F.; Primavilla, S.; Miraglia, D.; Andoni, E.; Di Cerbo, A.; Branciari, R. Improving Hunted wild boar carcass hygiene: Roles of different factors involved in the harvest phase. Foods 2021, 10, 1548. [Google Scholar] [CrossRef]
- Woolhouse, M.; Ward, M.; van Bunnik, B.; Farrar, J. Antimicrobial resistance in humans, livestock and the wider environment. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 1670. [Google Scholar] [CrossRef] [PubMed]
- Vittecoq, M.; Godreuil, S.; Prugnolle, F.; Durand, P.; Brazier, L.; Renaud, N.; Arnal, A.; Aberkane, S.; Jean-Pierre, H.; Gauthier-Clerc, M.; et al. Antimicrobial resistance in wildlife. J. Appl. Ecol. 2016, 53, 519–529. [Google Scholar] [CrossRef]
- Gilliver, M.A.; Bennett, M.; Begon, M.; Hazel, S.M.; Hart, C.A. Antibiotic resistance found in wild rodents. Nature 1999, 401, 233–234. [Google Scholar] [CrossRef]
- Österblad, M.; Norrdahl, K.; Korpimäki, E.; Huovinen, P. How wild are wild mammals? Nature 2001, 409, 37–38. [Google Scholar] [CrossRef]
- Tinoco Torres, R.; Fernandes, J.; Carvalho, J.; Cunha, M.V.; Caetano, T.; Mendo, S.; Serrano, E.; Fonseca, C. Wild boar as a reservoir of antimicrobial resistance. Sci. Total Environ. 2020, 717, 135001. [Google Scholar] [CrossRef]
- Plaza-Rodríguez, C.; Alt, K.; Grobbel, M.; Hammerl, J.A.; Irrgang, A.; Szabo, I.; Stingl, K.; Schuh, E.; Wiehle, L.; Pfefferkorn, B.; et al. Wildlife as sentinels of antimicrobial resistance in Germany? Front. Vet. Sci. 2021, 7, 627821. [Google Scholar] [CrossRef]
- Literak, I.; Dolejska, M.; Radimersky, T.; Klimes, J.; Friedman, M.; Aarestrup, F.M.; Hasman, H.; Cizek, A. Antimicrobial-resistant faecal Escherichia coli in wild mammals in Central Europe: Multiresistant Escherichia coli producing extended-spectrum beta-lactamases in wild boars. J. Appl. Microbiol. 2010, 108, 1702–1711. [Google Scholar] [CrossRef]
- Zottola, T.; Montagnaro, S.; Magnapera, C.; Sasso, S.; De Martino, L.; Bragagnolo, A.; D’Amici, L.; Condoleo, R.; Pisanelli, G.; Iovane, G.; et al. Prevalence and antimicrobial susceptibility of Salmonella in European wild boar (Sus scrofa); Latium region–Italy. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 161–168. [Google Scholar] [CrossRef]
- Modesto, P.; De Ciucis, C.G.; Vencia, W.; Pugliano, M.C.; Mignone, W.; Berio, E.; Masotti, C.; Ercolini, C.; Serracca, L.; Andreoli, T.; et al. Evidence of antimicrobial resistance and presence of pathogenicity genes in Yersinia enterocolitica isolate from wild boars. Pathogens 2021, 10, 398. [Google Scholar] [CrossRef]
- ISO 17604; Microbiology of the Food Chain. Carcass sampling for microbiological analysis. The International Organization for Standardization: Geneva, Switzerland, 2015.
- ISO 6579; Microbiology of the Food Chain. Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella. The International Organization for Standardization: Geneva, Switzerland, 2020.
- Kiskároly, F.; Morić, I.; Dokić, L.; Vasiljević, B.; Šenerović, L.; Mišić, D. Development of PCR-based identification of Salmonella enterica serovars. Acta Vet.-Beograd 2017, 67, 271–284. [Google Scholar] [CrossRef]
- ISO 10273; Microbiology of the Food Chain. Horizontal Method for the Detection of Pathogenic Yersinia Enterocolitica. The International Organization for Standardization: Geneva, Switzerland, 2017.
- Van Damme, I.; Berkvens, D.; Baré, J.; De Zutter, L. Influence of isolation methods on the occurrence of plasmid-carrying Yersinia enterocolitica serotype O:3 in slaughter pig tonsils, faeces and carcass surface swabs. Int. J. Food Microbiol. 2013, 164, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Piras, F.; Spanu, C.; Sanna, R.; Siddi, G.; Mocci, A.M.; Demontis, M.; Meloni, M.P.; Spanu, V.; De Santis, E.P.L.; Scarano, C. Detection, virulence genes and antimicrobial resistance of Yersinia enterocolitica in sheep and goat raw milk. Int. Dairy. J. 2021, 117, 105011. [Google Scholar] [CrossRef]
- Garzetti, D.; Susen, R.; Fruth, A.; Tietze, E.; Heesemann, J.; Rakin, A. A molecular scheme for Yersinia enterocolitica patho-serotyping derived from genome-wide analysis. Int. J. Med. Microbiol. 2014, 304, 275–283. [Google Scholar] [CrossRef] [PubMed]
- ISO 6887; Microbiology of the Food Chain. Preparation of Test Samples, Initial Suspension and Decimal Dilutions for Microbiological Examination. The International Organization for Standardization: Geneva, Switzerland, 2017.
- ISO 4833; Microbiology of the Food Chain. Horizontal Method for the Enumeration of Microorganisms. Part 1: Colony Count at 30 °C by the Pour Plate Technique. The International Organization for Standardization: Geneva, Switzerland, 2013.
- ISO 21528; Microbiology of the Food Chain. Horizontal Method for the Detection and Enumeration of Enterobacteriaceae. The International Organization for Standardization: Geneva, Switzerland, 2017.
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters 2023; Version 13.1. 2023. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 8 November 2023).
- Achtman, M.; Wain, J.; Weill, F.-X.; Nair, S.; Zhou, Z.; Sangal, V.; Krauland, M.G.; Hale, J.L.; Harbottle, H.; Uesbeck, A.; et al. Correction: Multilocus sequence typing as a replacement for serotyping in Salmonella enterica. PLoS Pathog. 2020, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Enterobase. Available online: http://enterobase.warwick.ac.uk/ (accessed on 8 November 2023).
- Zhou, Z.; Alikhan, N.-F.; Mohamed, K.; Fan, Y.; Achtman, M. The user’s guide to comparative genomics with EnteroBase, including case studies on transmissions of micro-clades of Salmonella, the phylogeny of ancient and modern Yersinia pestis genomes, and the core genomic diversity of all Escherichia. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Chattaway, M.A.; Reuter, S.; Savin, C.; Strauch, E.; Carniel, E.; Connor, T.; VanDamme, I.; Rajakaruna, L.; Rajendram, D.; et al. Use of whole-genus genome sequence data to develop a multilocus sequence typing tool that accurately identifies Yersinia isolates to the species and subspecies levels. J. Clin. Microbiol. 2015, 53, 35–42. [Google Scholar] [CrossRef] [PubMed]
- AMR Finder. Available online: https://github.com/ncbi/amr (accessed on 8 November 2023).
- ResFinder. Available online: https://cge.food.dtu.dk/services/ResFinder/ (accessed on 8 November 2023).
- PlasmidFinder. Available online: https://cge.cbs.dtu.dk/services/PlasmidFinder/ (accessed on 8 November 2023).
- VirulenceFinder. Available online: https://cge.food.dtu.dk/services/VirulenceFinder/ (accessed on 8 November 2023).
- Piras, F.; Spanu, V.; Siddi, G.; Gymoese, P.; Spanu, C.; Cibin, V.; Schjørring, S.; De Santis, E.P.L.; Scarano, C. Whole-genome sequencing analysis of highly prevalent Salmonella serovars in wild boars from a national park in Sardinia. Food Control 2021, 130, 108247. [Google Scholar] [CrossRef]
- Cilia, G.; Turchi, B.; Fratini, F.; Bilei, S.; Bossù, T.; De Marchis, M.L.; Cerri, D.; Pacini, M.I.; Bertelloni, F. Prevalence, virulence and antimicrobial susceptibility of Salmonella spp., Yersinia enterocolitica and Listeria monocytogenes in European Wild Boar (Sus scrofa) hunted in Tuscany (Central Italy). Pathogens 2021, 10, 93. [Google Scholar] [CrossRef] [PubMed]
- Razzuoli, E.; Listorti, V.; Martini, I.; Migone, L.; Decastelli, L.; Mignone, W.; Berio, E.; Battistini, R.; Ercolini, C.; Serracca, L.; et al. Prevalence and antimicrobial resistances of Salmonella spp. isolated from Wild Boars in Liguria Region, Italy. Pathogens 2021, 10, 568. [Google Scholar] [CrossRef]
- Botti, V.; Navillod, F.V.; Domenis, L.; Orusa, R.; Pepe, E.; Robetto Guidetti, C. Salmonella spp. and antibiotic-resistant strains in wild mammals and birds in north-western Italy from 2002 to 2010. Vet. Ital. 2013, 49, 195–202. [Google Scholar]
- Stella, S.; Tirloni, E.; Castelli, E.; Colombo, F.; Bernardi, C. Microbiological evaluation of carcasses of Wild Boar hunted in a hill area of Northern Italy. J. Food Prot. 2018, 81, 1519–1525. [Google Scholar] [CrossRef]
- Bonardi, S.; Bolzoni, L.; Zanoni, R.G.; Morganti, M.; Corradi, M.; Gilioli, S.; Pongolini, S. Limited exchange of Salmonella among domestic pigs and wild boars in Italy. Ecohealth 2019, 16, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Vieira-Pinto, M.; Morais, L.; Caleja, C.; Themudo, P.; Aranha, J.; Torres, C.; Igrejas, G.; Poeta, P.; Martins, C. Salmonella spp. in wild boar (Sus scrofa): A public and animal health concern. In Game Meat Hygiene in Focus; Wageningen Academic Publishers: Wageningen, The Netherlands, 2011; pp. 131–136. [Google Scholar] [CrossRef]
- Sannö, A.; Aspán, A.; Hestvik, G.; Jacobson, M. Presence of Salmonella spp.; Yersinia enterocolitica, Yersinia pseudotuberculosis and Escherichia coli O157:H7 in wild boars. Epidemiol. Infect. 2014, 142, 2542–2547. [Google Scholar] [CrossRef] [PubMed]
- Sannö, A.; Rosendal, T.; Aspán, A.; Backhans, A.; Jacobson, M. Distribution of enteropathogenic Yersinia spp. and Salmonella spp. in the Swedish wild boar population, and assessment of risk factors that may affect their prevalence. Acta Vet. Scand. 2018, 60, 40. [Google Scholar] [CrossRef] [PubMed]
- Gil Molino, M.; García Sánchez, A.; Risco Pérez, D.; Gonçalves Blanco, P.; Quesada Molina, A.; Rey Pérez, J.; Martín Cano, F.E.; Cerrato Horrillo, R.; Hermoso-de-Mendoza Salcedo, J.; Fernández Llario, P. Prevalence of Salmonella spp. in tonsils, mandibular lymph nodes and faeces of wild boar from Spain and genetic relationship between isolates. Transbound. Emerg. Dis. 2019, 66, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Vigo, G.B.; Cappuccio, J.A.; Piñeyro, P.E.; Salve, A.; Machuca, M.A.; Quiroga, M.A.; Moredo, F.; Giacoboni, G.; Cancer, J.L.; Caffer, I.G.; et al. Salmonella enterica subclinical infection: Bacteriological, serological, pulsed-field gel electrophoresis, and antimicrobial resistance profiles-longitudinal study in a three-site farrow-to-finish farm. Foodborne Pathog. Dis. 2009, 6, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Molla, B.; Sterman, A.; Mathews, J.; Artuso-Ponte, V.; Abley, M.; Farmer, W.; Rajala-Schultz, P.; Morrow, M.W.E.; Gebreyes, W.A. Salmonella enterica in commercial swine feed and subsequent isolation of phenotypically and genotypically related strains from fecal samples. Appl. Environ. Microbiol. 2010, 76, 21. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.F.; Funk, J.A.; Bolin, C. Risk factors associated with persistence of Salmonella shedding in finishing pigs. Prev. Vet. Med. 2014, 116, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, M.A.; Rodriguez-Zas, S.L.; Ellis, M.; Salak-Johnson, J.L. Breed and age affect baseline immune traits, cortisol, and performance in growing pigs. Sci. J. Anim. Sci. 2005, 83, 2087–2095. [Google Scholar] [CrossRef]
- Pesciaroli, M.; Cucco, L.; De Luca, S.; Massacci, F.R.; Maresca, C.; Medici, L.; Paniccià, M.; Scoccia, E.; Staffolani, M.; Pezzotti, G.; et al. Association between pigs with high caecal Salmonella loads and carcass contamination. Int. J. Food Microbiol. 2017, 242, 82–86. [Google Scholar] [CrossRef]
- Hidalgo-Vila, J.; Díaz-Paniagua, C.; de Frutos-Escobar, C.; Jiménez-Martínez, C.; Pérez-Santigosa, N. Salmonella in free living terrestrial and aquatic turtles. Vet. Microbiol. 2007, 119, 311–315. [Google Scholar] [CrossRef]
- Scheelings, T.F.; Lightfoot, D.; Holz, P. Prevalence of Salmonella in Australian reptiles. J. Wildl. Dis. 2011, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bjelland, A.M.; Sandvik, L.M.; Skarstein, M.M.; Svendal, L.; Debenham, J.J. Prevalence of Salmonella serovars isolated from reptiles in Norwegian zoos. Acta Vet. Scand. 2020, 62, 3. [Google Scholar] [CrossRef] [PubMed]
- Orlandella, B.M. Migrating birds and Infectious Diseases: Research on the role of the migrating birds in the spread of Salmonella. A First Report. Acta Med. Vet. 1986, 32, 73–87. [Google Scholar]
- Orlandella, V.; Alosi, C.; Campagna, A.; Conti, F.; Ilacqua, G.; Coppola, L. Studies on the epidemic-epizootologic role of Lacertilia in the diffusion of Salmonella. II. Isolation of S. elomrane, S. elomrane, var. 01, S. minnesota, S. siegburg, S. siegburg, var. sandinia, S. uphill and S. rhône from chalcides ocellatus tiligugu, Gmelin (syn. Gongyllus ocellatus, Bedriaga) of the family Scincidae. G. Di Batteriol. Virol. Immunol. Ann. Dell’ospedale Maria Vittor. Torino 1971, 64, 300–315. [Google Scholar]
- Bertrand, S.; Rimhanen-Finne, R.; Weill, F.X.; Rabsch, W.; Thornton, L.; Perevoščikovs, J.; van Pelt, W.; Heck, M. Salmonella infections associated with reptiles: The current situation in Europe. Euro Surveill. 2008, 13, 18902. [Google Scholar] [CrossRef] [PubMed]
- Wikström, V.O.; Fernström, L.; Melin, L.; Boqvist, S. Salmonella isolated from individual reptiles and environmental samples from terraria in private households in Sweden. Acta Vet. Scand. 2014, 56, 7. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Miranda, J.M.; Regal, P.; V’azquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Galán, J.E. Molecular genetic bases of Salmonella entry into host cells. Mol. Microbiol. 1996, 20, 263–271. [Google Scholar] [CrossRef]
- Bäumler, A.J.; Tsolis, R.M.; Valentine, P.J.; Ficht, T.A.; Heffron, F. Synergistic effect of mutations in invA and lpfC on the ability of Salmonella Typhimurium to cause murine typhoid. Infect. Immun. 1997, 65, 2254–2259. [Google Scholar] [CrossRef]
- van der Velden, A.W.M.; Bäumler, A.J.; Tsolis, R.M.; Heffron, F. Multiple fimbrial adhesins are required for full virulence of Salmonella Typhimurium in mice. Infect. Immun. 1998, 66, 2803–2808. [Google Scholar] [CrossRef]
- Weening, E.H.; Barker, J.D.; Laarakker, M.C.; Humphries, A.D.; Tsolis, R.M.; Bäumler, A.J. The Salmonella enterica serotype Typhimurium lpf, bcf, stb, stc, std, and sth fimbrial operons are required for intestinal persistence in mice. Infect. Immun. 2005, 73, 3358–3366. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, I.E.; Ghori, N.; Falkow, S.; Monack, D. Mig-14 is an inner membrane-associated protein that promotes Salmonella typhimurium resistance to CRAMP, survival within activated macrophages and persistent infection. Mol. Microbiol. 2005, 55, 954–972. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Puente, J.L.; Calva, E. Salmonella virulence plasmid: Pathogenesis and ecology. Pathog. Dis. 2017, 75, ftx070. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Humphries, A.D.; Weening, E.H.; De Zoete, M.R.; Winter, S.; Papaconstantinopoulou, A.; Dougan, G.; Bäumler, A.J. Molecular and phenotypic analysis of the CS54 island of Salmonella enterica serotype Typhimurium: Identification of intestinal colonization and persistence determinants. Infect. Immun. 2003, 71, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Matthews, T.D.; Schmieder, R.; Silva, G.G.; Busch, J.; Cassman, N.; Dutilh, B.E.; Green, D.; Matlock, B.; Heffernan, B.; Olsen, G.J. Genomic comparison of the closely-related Salmonella enterica serovars Enteritidis, Dublin and Gallinarum. PLoS ONE 2015, 10, 0126883. [Google Scholar] [CrossRef]
- Miller, R.; Wiedmann, M. Dynamic Duo-The Salmonella cytolethal distending toxin combines adp-ribosyltransferase and nuclease activities in a novel form of the cytolethal distending toxin. Toxins 2016, 8, 121. [Google Scholar] [CrossRef]
- Bonardi, S.; Brémont, S.; Vismarra, A.; Poli, I.; Diegoli, G.; Bolzoni, L.; Corradi, M.; Gilioli, S.; Le Guern, A.S. Is Yersinia bercovieri surpassing Yersinia enterocolitica in Wild Boars (Sus scrofa)? Ecohealth 2020, 17, 388–392. [Google Scholar] [CrossRef]
- Bancerz-Kisiel, A.; Platt-Samoraj, A.; Szczerba-Turek, A.; Syczyło, K.; Szweda, W. The first pathogenic Yersinia enterocolitica bioserotype 4/O:3 strain isolated from a hunted wild boar (Sus scrofa) in Poland. Epidemiol. Infect. 2015, 143, 2758–2765. [Google Scholar] [CrossRef]
- von Altrock, A.; Seinige, D.; Kehrenberg, C. Yersinia enterocolitica Isolates from Wild Boars Hunted in Lower Saxony, Germany. Appl. Environ. Microbiol. 2015, 81, 4835–4840. [Google Scholar] [CrossRef]
- Arrausi-Subiza, M.; Gerrikagoitia, X.; Alvarez, V.; Ibabe, J.C.; Barral, M. Prevalence of Yersinia enterocolitica and Yersinia pseudotuberculosis in wild boars in the Basque Country, northern Spain. Acta Vet. Scand. 2016, 58, 4. [Google Scholar] [CrossRef]
- Slee, K.J.; Skilbeck, N.W. Epidemiology of Yersinia pseudotuberculosis and Y. enterocolitica infections in sheep in Australia. J. Clin. Microbiol. 1992, 30, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Söderqvist, K.; Boqvist, S.; Wauters, G.; Vågsholm, I.; Thisted-Lambertz, S. Yersinia enterocolitica in sheep-a high frequency of biotype 1A. Acta Vet. Scand. 2012, 54, 39. [Google Scholar] [CrossRef] [PubMed]
- Piras, F.; Siddi, G.; Le Guern, A.S.; Brémont, S.; Fredriksson-Ahomaa, M.; Sanna, R.; Meloni, M.P.; De Santis, E.P.L.; Scarano, C. Traceability, virulence and antimicrobial resistance of Yersinia enterocolitica in two industrial cheese-making plants. Int. J. Food Microbiol. 2023, 398, 110225. [Google Scholar] [CrossRef] [PubMed]
- Milnes, A.S.; Sayers, A.R.; Stewart, I.; Clifton-Hadley, F.A.; Davies, R.H.; Newell, D.G.; Cook, A.J.; Evans, S.J.; Smith, R.P.; Paiba, G.A. Factors related to the carriage of Verocytotoxigenic E. coli, Salmonella, thermophilic Campylobacter and Yersinia enterocolitica in cattle, sheep and pigs at slaughter. Epidemiol. Infect. 2009, 137, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Mair, N.S. Yersiniosis in wildlife and its public health implications. J. Wildl. Dis. 1973, 9, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Bancerz-Kisiel, A.; Pieczywek, M.; Łada, P.; Szweda, W. The most important virulence markers of Yersinia enterocolitica and their role during infection. Genes 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Ellison, D.W.; Young, B.; Nelson, K.; Miller, V.L. YMOA negatively regulates expression of invasin from Yersinia enterocolitica. J. Bacteriol. 2003, 185, 7153–7159. [Google Scholar] [CrossRef] [PubMed]
- Fetherston, J.D.; Mier, I.; Truszczynska, H.; Perry, R.D. The YFE and feo transporters are involved in microaerobic growth and virulence of Yersinia pestis in bubonic plague. Infect. Immun. 2012, 80, 3880–3891. [Google Scholar] [CrossRef]
- Howard, S.L.; Gaunt, M.W.; Hinds, J.; Witney, A.A.; Stabler, R.; Wren, B.W. Application of comparative phylogenomics to study the evolution of Yersinia enterocolitica and to identify genetic differences relating to pathogenicity. J. Bacteriol. 2006, 188, 3645–3653. [Google Scholar] [CrossRef]
- Falcão, J.P.Y.; Batt, C.A.; Tortorello, M.L. Encyclopedia of Food Microbiology, 2nd ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-384733-1. [Google Scholar]
- Sprague, L.D.; Neubauer, H. Yersinia aleksiciae sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 831–835. [Google Scholar] [CrossRef]
- Stock, I.; Heisig, P.; Wiedemann, B. β-Lactamase expression in Yersinia enterocolitica biovars 1A, 1B and 3. J. Med. Microbiol. 2000, 49, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Seoane, A.; García Lobo, J.M. Identification of a streptogramin, an acetyltransferase gene in the chromosome of Yersinia enterocolitica. Antimicrob. Agents Chemother. 2000, 44, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Stogios, P.J.; Kuhn, M.L.; Evdokimova, E.; Courvalin, P.; Anderson, W.F.; Savchenko, A. Potential for reduction of streptogramin A resistance revealed by structural analysis of acetyltransferase VatA. Antimicrob. Agents Chemother. 2014, 58, 7083–7092. [Google Scholar] [CrossRef] [PubMed]
- Mirceta, J.; Petrovic, J.; Malesevic, M.; BlaAgojevic, B.; Antic, D. Assessment of microbial carcass contamination of hunted wild boars. Eur. J. Wildl. Res. 2017, 63, 37–44. [Google Scholar] [CrossRef]
- Bonardi, S.; Tansini, C.; Cacchioli, A.; Soliani, L.; Poli, L.; Lamperti, L.; Corradi, M.; Gilioli, S. Enterobacteriaceae and Salmonella contamination of wild boar (Sus scrofa) carcasses: Comparison between different sampling strategies. Eur. J. Wildl. Res. 2021, 67, 88. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Tested Animals | Positive Animals | Positive Samples | |||
|---|---|---|---|---|---|---|
| Colon Content | Mesenteric Lymph Nodes | Carcass Surface | Total | |||
| Salmonella | 66 | 3 | 3/66 (4.5) | 0/66 | 0/49 | 3/181 (1.6) |
| Y. enterocolitica | 66 | 20 | 18/66 (27.3) | 3/66 (4.5) | 3/49 (6.1) | 24/181 (13.2) |
| Hunting Houses | Sampled Wild Boars | Total Positive Wild Boars | Pathogen | |
|---|---|---|---|---|
| Salmonella | Y. enterocolitica | |||
| A | 28 | 12 | 2 | 10 |
| B | 18 | 7 | 1 | 6 |
| C | 7 | 1 | 0 | 1 |
| D | 8 | 2 | 0 | 2 |
| E | 5 | 1 | 0 | 1 |
| Microorganism | MLST ST | Phenotypic Profile | Genotypic Profile |
|---|---|---|---|
| Salmonella | 10546 | ND | - |
| 7139 | ND | - | |
| 10597 | ND | - | |
| Y. enterocolitica | 332 | Amp Aug | blaA, vat(F) |
| 443 | Amp Aug Fox | blaA | |
| 852 | Amp Aug | blaA | |
| 854 | Amp Aug | blaA | |
| 855 | Amp Aug | blaA | |
| 856 | Amp | blaA | |
| 857 | ND | blaA | |
| 858 | Amp Aug | blaA | |
| 860 (1) | Amp Aug Fox | blaA | |
| 860 (2) | Amp Aug | blaA | |
| 860 (3) | Amp Aug | blaA | |
| 860 (4) | Amp Aug | blaA | |
| 860 (5) | Amp Aug Fox | blaA | |
| 861 | Amp Aug Fox | blaA | |
| 862 | Amp Aug Fox | blaA | |
| 863 | Amp Aug | blaA | |
| 864 | Amp Aug Fox | blaA | |
| 865 | ND | blaA | |
| 866 | Amp Aug Fox | blaA | |
| 867 | Amp Aug | blaA | |
| 868 | Amp Aug | blaA | |
| 870 | Amp Aug Fox | blaA | |
| 872 | Amp Aug Fox | blaA | |
| 873 | Amp Aug Fox | blaA |
| Sampling Days | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hunting Houses | A | A | B | A | C | A | B | A | B | A | B | B | |
| Number of Samples | 7 | 1 | 8 | 8 | 3 | 5 | 2 | 6 | 4 | 1 | 2 | 2 | |
| ACC | Mean | 1.61 | 1.33 | 2.68 | 2.85 | 3.43 | 1.69 | 2.83 | 2.48 | 2.70 | 2.87 | 3.12 | 1.85 |
| Median | 1.50 | 1.33 | 2.50 | 3.02 | 3.37 | 1.32 | 2.83 | 1.96 | 2.61 | 2.87 | 3.21 | 1.85 | |
| Minimum value | 1.19 | - | 1.78 | 1.33 | 2.93 | 1.07 | 1.97 | 1.11 | 1.81 | - | 2.51 | 1.47 | |
| Maximum value | 2.14 | - | 3.83 | 4.15 | 4.00 | 2.89 | 3.68 | 4.35 | 3.78 | - | 3.91 | 2.23 | |
| Ent | Mean | 0.58 | 0.15 | 0.89 | 3.68 | 2.52 | 0.10 | 0.84 | 1.52 | 0.57 | 0.30 | 0.77 | 0.00 |
| Median | 0.33 | 0.15 | 0.61 | 4.00 | 2.48 | 0.00 | 0.84 | 1.53 | 0.29 | 0.30 | 0.77 | 0.00 | |
| Minimum value | 0.00 | - | 0.00 | 2.67 | 1.60 | 0.00 | 0.15 | 0.00 | 0.00 | - | 0.67 | - | |
| Maximum value | 2.01 | - | 2.16 | 4.00 | 3.48 | 0.51 | 1.53 | 2.87 | 1.70 | - | 0.87 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddi, G.; Piras, F.; Meloni, M.P.; Gymoese, P.; Torpdahl, M.; Fredriksson-Ahomaa, M.; Migoni, M.; Cabras, D.; Cuccu, M.; De Santis, E.P.L.; et al. Hunted Wild Boars in Sardinia: Prevalence, Antimicrobial Resistance and Genomic Analysis of Salmonella and Yersinia enterocolitica. Foods 2024, 13, 65. https://doi.org/10.3390/foods13010065
Siddi G, Piras F, Meloni MP, Gymoese P, Torpdahl M, Fredriksson-Ahomaa M, Migoni M, Cabras D, Cuccu M, De Santis EPL, et al. Hunted Wild Boars in Sardinia: Prevalence, Antimicrobial Resistance and Genomic Analysis of Salmonella and Yersinia enterocolitica. Foods. 2024; 13(1):65. https://doi.org/10.3390/foods13010065
Chicago/Turabian StyleSiddi, Giuliana, Francesca Piras, Maria Pina Meloni, Pernille Gymoese, Mia Torpdahl, Maria Fredriksson-Ahomaa, Mattia Migoni, Daniela Cabras, Mario Cuccu, Enrico Pietro Luigi De Santis, and et al. 2024. "Hunted Wild Boars in Sardinia: Prevalence, Antimicrobial Resistance and Genomic Analysis of Salmonella and Yersinia enterocolitica" Foods 13, no. 1: 65. https://doi.org/10.3390/foods13010065
APA StyleSiddi, G., Piras, F., Meloni, M. P., Gymoese, P., Torpdahl, M., Fredriksson-Ahomaa, M., Migoni, M., Cabras, D., Cuccu, M., De Santis, E. P. L., & Scarano, C. (2024). Hunted Wild Boars in Sardinia: Prevalence, Antimicrobial Resistance and Genomic Analysis of Salmonella and Yersinia enterocolitica. Foods, 13(1), 65. https://doi.org/10.3390/foods13010065