

Polymorphisms of ITGA9 Gene and Their Correlation with Milk Quality Traits in Yak (Bos grunniens)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Sample Collection of Test Animals
2.2.1. Milk Samples
2.2.2. Tissue Samples
2.3. Milk Composition Analysis
2.4. DNA Extraction
2.5. Genotyping
2.6. Statistical Analysis
2.7. LD Linkage Disequilibrium and Haplotype Analyses
2.8. KEGG Signalling Pathway Enrichment Analysis
3. Results
3.1. Genotype Results for ITGA9 and Genetic Parameter Analysis of the Loci in Yak
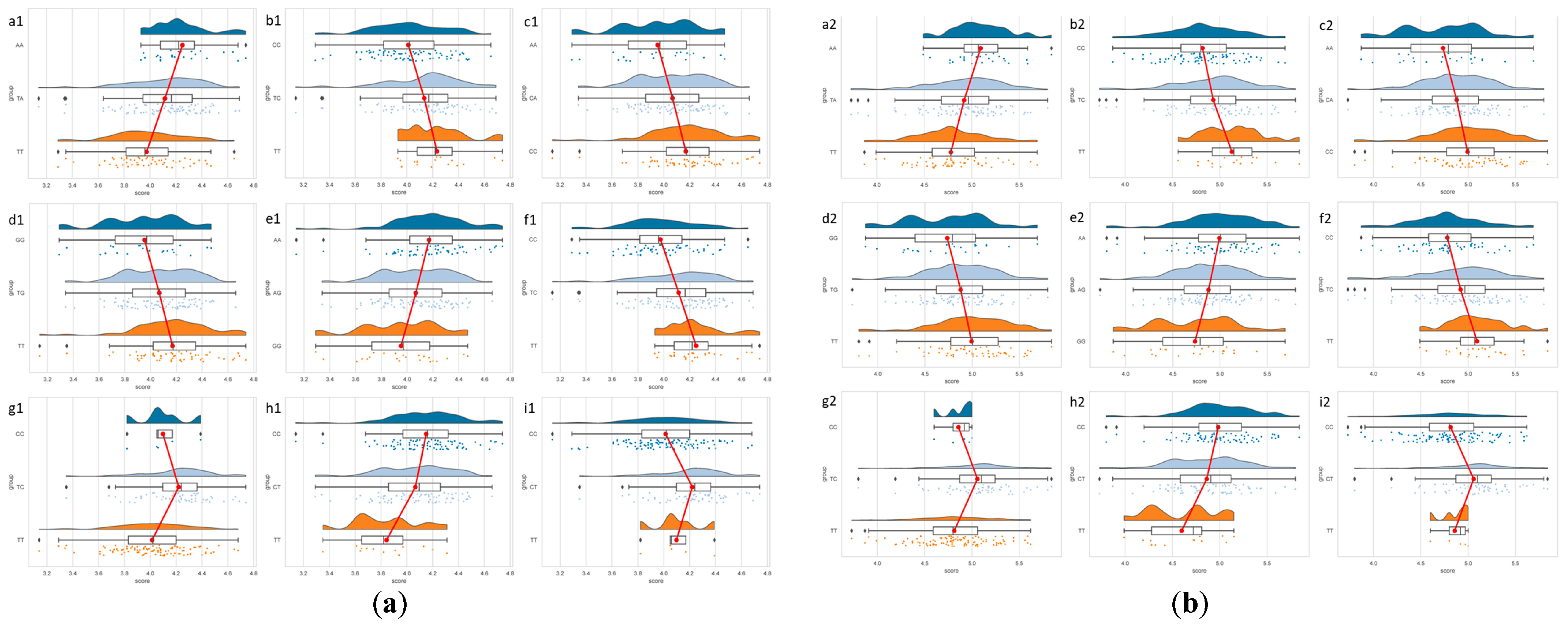
3.2. Association Analysis of ITGA9 Genotypes with Yak Milk Quality Traits
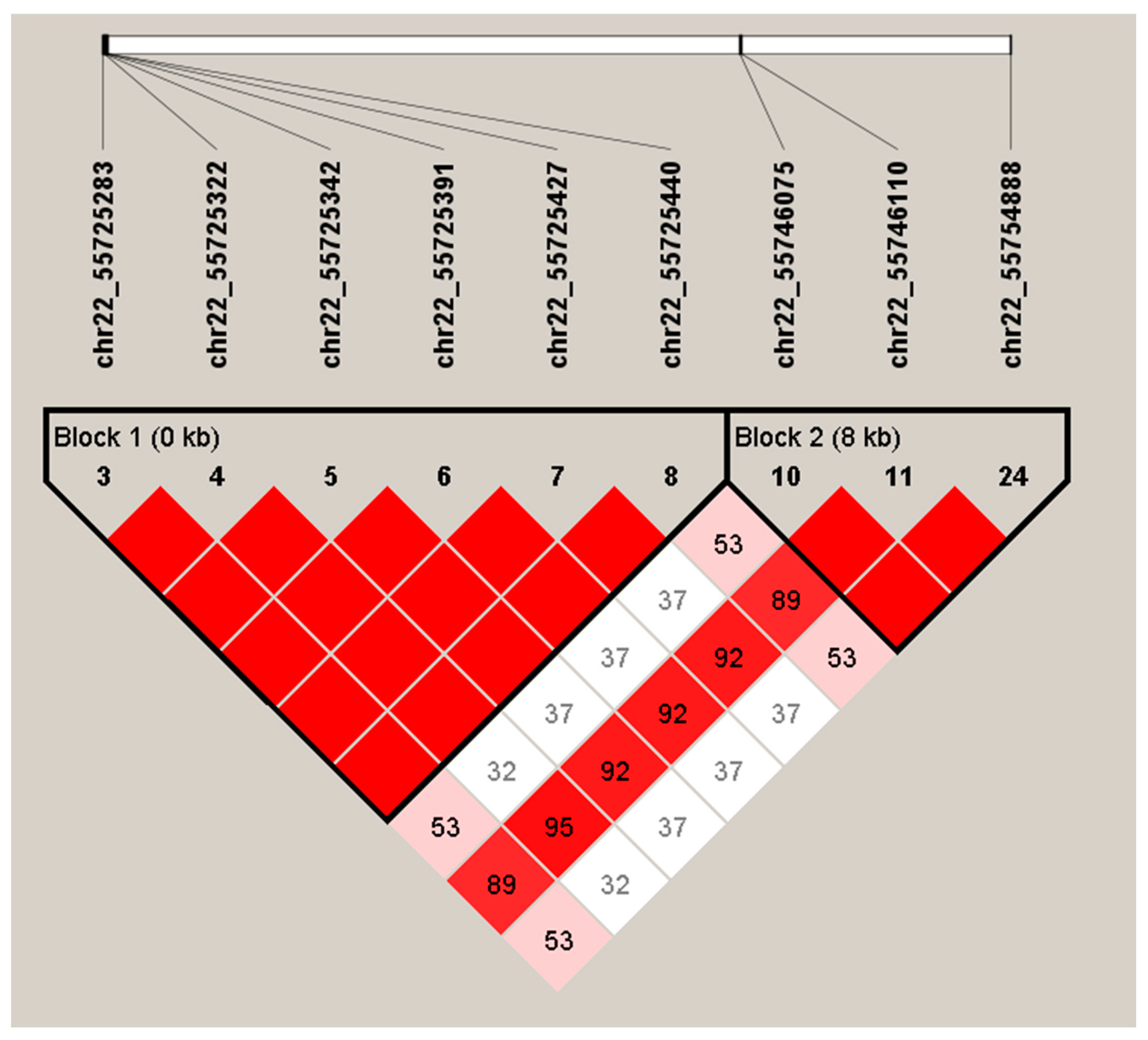
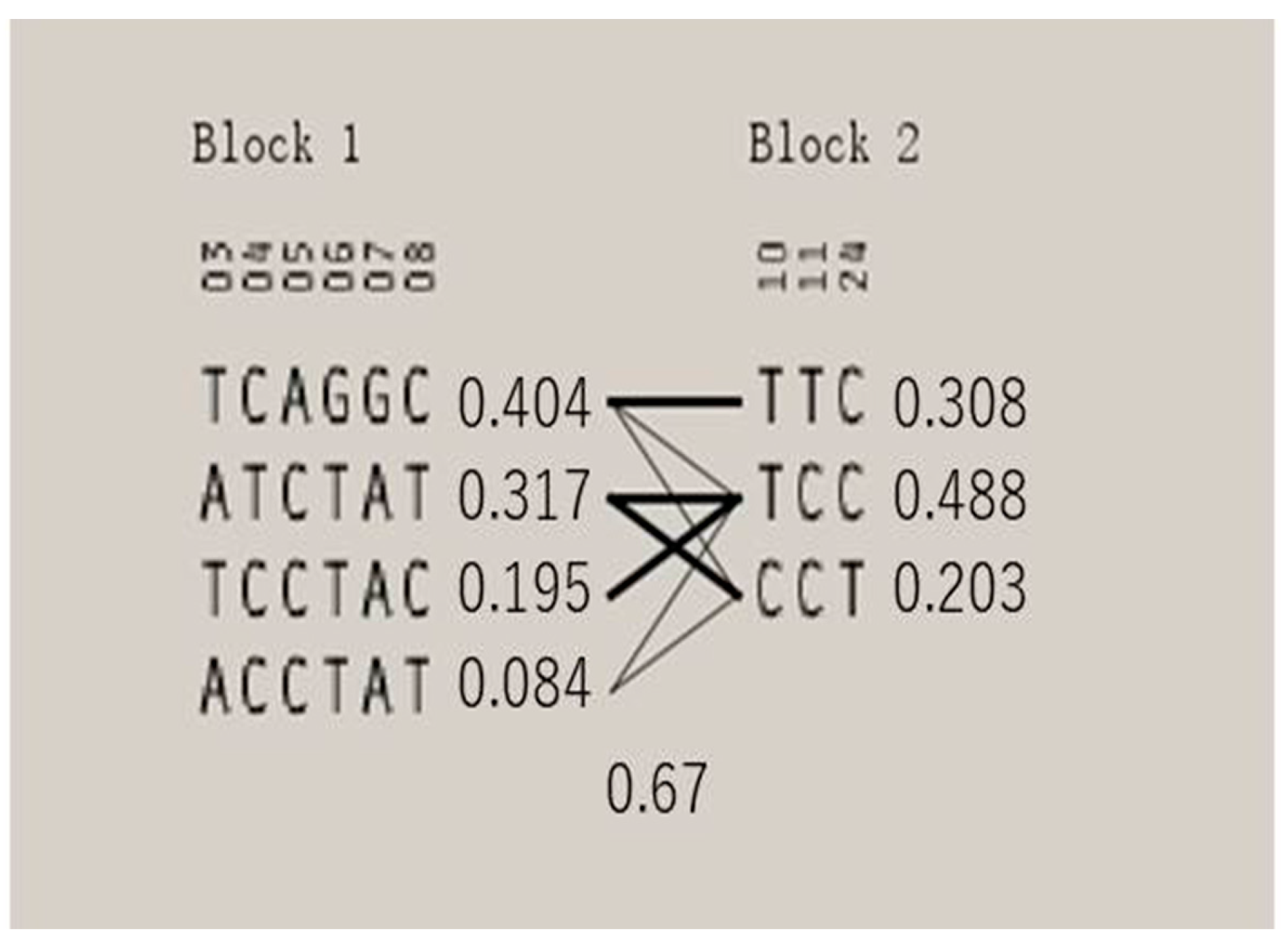
3.3. LD Analysis and Haplotype Analysis
3.4. KEGG Signalling Pathway Annotation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ma, J.; Zhu, Y.; Wang, Z.; Yu, X.; Hu, R.; Wang, X.; Cao, G.; Zou, H.; Shah, A.M.; Peng, Q.; et al. Comparing the Bacterial Community in the Gastrointestinal Tracts between Growth-Retarded and Normal Yaks on the Qinghai-Tibetan Plateau. Front. Microbiol. 2020, 11, 600516. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Ge, X.; Yang, L.; Ma, G.; Ma, J.; Yu, Q.-L.; Han, L. Ultrasound-assisted thawing of frozen white yak meat: Effects on thawing rate, meat quality, nutrients, and microstructure. Ultrason. Sonochemistry 2021, 70, 105345. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ma, Y.; Li, Q.; Wang, J.; Cheng, J.; Xue, J.; Shi, J. The Chemical Composition and Nitrogen Distribution of Chinese Yak (Maiwa) Milk. Int. J. Mol. Sci. 2011, 12, 4885–4895. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhou, Y.; Zheng, X.; Guo, J.; Duan, H.; Zhou, S.; Yan, W. Yak Milk: Nutritional Value, Functional Activity, and Current Applications. Foods 2023, 12, 2090. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, X.; Yu, S.; Cao, S. Quantitative proteomic analysis of whey proteins in the colostrum and mature milk of yak (Bos grunniens). J. Sci. Food Agric. 2015, 95, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chen, S.; Ren, F.; Luo, Z.; Zeng, S.S. Yak milk casein as a functional ingredient: Preparation and identification of angiotensin-I-converting enzyme inhibitory peptides. J. Dairy Res. 2007, 74, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.-Y.; Cheng, X.; Wang, X.; Wu, S.-J. Free-radical-scavenging and anti-inflammatory effect of yak milk casein before and after enzymatic hydrolysis. Food Chem. 2011, 126, 484–490. [Google Scholar] [CrossRef]
- Lin, K.; Zhang, L.-W.; Han, X.; Xin, L.; Meng, Z.-X.; Gong, P.-M.; Cheng, D.-Y. Yak milk casein as potential precursor of angiotensin I-converting enzyme inhibitory peptides based on in silico proteolysis. Food Chem. 2018, 254, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Phelan, M.; Aherne, A.; FitzGerald, R.J.; O’Brien, N.M. Casein-derived bioactive peptides: Biological effects, industrial uses, safety aspects and regulatory status. Int. Dairy J. 2009, 19, 643–654. [Google Scholar] [CrossRef]
- Aguilar, I.; Misztal, I.; Tsuruta, S. Short communication: Genetic trends of milk yield under heat stress for US Holsteins. J. Dairy Sci. 2010, 93, 1754–1758. [Google Scholar] [CrossRef] [PubMed]
- Bignardi, A.B.; El Faro, L.; Pereira, R.J.; Santana, M.L.; Stefani, G. Genetics of heat tolerance for milk yield and quality in Holsteins. Animal 2016, 11, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, Z.; Zhao, W.; Guo, L.; Sun, H.; Zhu, K.; Liu, G.; Shen, X.; Zhao, X.; Wang, Q.; et al. Genome-wide selection signatures detection in Shanghai Holstein cattle population identified genes related to adaption, health and reproduction traits. BMC Genom. 2021, 22, 747. [Google Scholar] [CrossRef]
- Peripolli, E.; Stafuzza, N.B.; Munari, D.P.; Lima, A.L.F.; Irgang, R.; Machado, M.A.; Panetto, J.; Ventura, R.V.; Baldi, F.; da Silva, M. Assessment of runs of homozygosity islands and estimates of genomic inbreeding in Gyr (Bos indicus) dairy cattle. BMC Genom. 2018, 19, 34. [Google Scholar] [CrossRef] [PubMed]
- Bisutti, V.; Mach, N.; Giannuzzi, D.; Vanzin, A.; Capra, E.; Negrini, R.; Gelain, M.E.; Cecchinato, A.; Ajmone-Marsan, P.; Pegolo, S. Transcriptome-wide mapping of milk somatic cells upon subclinical mastitis infection in dairy cattle. J. Anim. Sci. Biotechnol. 2023, 14, 93. [Google Scholar] [CrossRef] [PubMed]
- Mostovich, L.A.; Prudnikova, T.Y.; Kondratov, A.G.; Loginova, D.; Vavilov, P.V.; Rykova, V.I.; Sidorov, S.V.; Pavlova, T.V.; Kashuba, V.I.; Zabarovsky, E.R.; et al. Integrin alpha9 (ITGA9) expression and epigenetic silencing in human breast tumors. Cell Adhes. Migr. 2011, 5, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Jin, L.; He, Z.; Wei, T.; Luo, T.; Zhang, J.; Liu, C.; Dai, C.; Chao, A.; Liang, Y.; et al. A cell transcriptomic profile provides insights into adipocytes of porcine mammary gland across development. J. Anim. Sci. Biotechnol. 2023, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Long, R.; Kreuzer, M.; Ding, L.; Shang, Z.; Zhang, Y.; Yang, Y.; Cui, G. Importance of Functional Ingredients in Yak Milk-Derived Food on Health of Tibetan Nomads Living Under High-Altitude Stress: A Review. Crit. Rev. Food Sci. Nutr. 2014, 54, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Kalwar, Q.; Ma, X.; Xi, B.; Korejo, R.A.; Bhuptani, D.K.; Chu, M.; Yan, P. Yak milk and its health benefits: A comprehensive review. Front. Vet. Sci. 2023, 10, 1213039. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.P.; Arora, S.; Sarkar, M. Yak milk and milk products: Functional, bioactive constituents and therapeutic potential. Int. Dairy J. 2023, 142, 105637. [Google Scholar] [CrossRef]
- Kulyar, M.F.-E.-A.; Yao, W.; Ding, Y.; Li, K.; Zhang, L.; Li, A.; Waqas, M.; Huachun, P.; Quan, M.; Zeng, Z.; et al. Bioactive potential of yak’s milk and its products; pathophysiological and molecular role as an immune booster in antibiotic resistance. Food Biosci. 2021, 39, 100838. [Google Scholar] [CrossRef]
- Dilzer, A.; Park, Y. Implication of Conjugated Linoleic Acid (CLA) in Human Health. Crit. Rev. Food Sci. Nutr. 2012, 52, 488–513. [Google Scholar] [CrossRef] [PubMed]
- Thivierge, M.C.; Petitclerc, D.; Bernier, J.F.; Couture, Y.; Lapierre, H. Variations in Mammary Metabolism during the Natural Filling of the Udder with Milk over a 12-h Period between Two Milkings1. J. Dairy Sci. 2002, 85, 1839–1854. [Google Scholar] [CrossRef] [PubMed]
- Tosif, M.M.; Najda, A. A Comprehensive Review on the Interaction of Milk Protein Concentrates with Plant-Based Polyphenolics. Int. J. Mol. Sci. 2021, 22, 13548. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.H.; Yun, S.S.; Lee, W.J.; Kim, J.W.; Ha, H.K.; Yoo, M.; Hwang, H.J.; Jeon, W.M.; Han, K.S. Hydrolysis by Alcalase Improves Hypoallergenic Properties of Goat Milk Protein. Korean J. Food Sci. Anim. Resour. 2016, 36, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Perinelli, D.R.; Bonacucina, G.; Cespi, M.; Bonazza, F.; Palmieri, G.F.; Pucciarelli, S.; Polzonetti, V.; Attarian, L.; Polidori, P.; Vincenzetti, S. A comparison among β-caseins purified from milk of different species: Self-assembling behaviour and immunogenicity potential. Colloids Surf. B Biointerfaces 2019, 173, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.M.; Bhandari, B.R.; Bansal, N. Functionality of bovine milk proteins and other factors in foaming properties of milk: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 4800–4820. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Zhang, L.; Han, X.; Meng, Z.; Zhang, J.; Wu, Y.; Cheng, D. Quantitative Structure–Activity Relationship Modeling Coupled with Molecular Docking Analysis in Screening of Angiotensin I-Converting Enzyme Inhibitory Peptides from Qula Casein Hydrolysates Obtained by Two-Enzyme Combination Hydrolysis. J. Agric. Food Chem. 2018, 66, 3221–3228. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.R.; Silva, T.J.; Barros, E.; Guimarães, V.M.; Baracat-Pereira, M.C.; Eller, M.R.; dos Reis Coimbra, J.S.; de Oliveira, E.B. Anti-Hypertensive Peptides Derived from Caseins: Mechanism of Physiological Action, Production Bioprocesses, and Challenges for Food Applications. Appl. Biochem. Biotechnol. 2018, 185, 884–908. [Google Scholar] [CrossRef] [PubMed]
- Shastry, B.S. SNPs: Impact on gene function and phenotype. Methods Mol. Biol. 2009, 578, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.-Y.; Lee, K.H. From SNPs to functional polymorphism: The insight into biotechnology applications. Biochem. Eng. J. 2010, 49, 149–158. [Google Scholar] [CrossRef]
- Weikard, R.; Kühn, C.; Goldammer, T.; Freyer, G.; Schwerin, M. The bovine PPARGC1A gene: Molecular characterization and association of an SNP with variation of milk fat synthesis. Physiol. Genom. 2005, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dagnachew, B.S.; Thaller, G.; Lien, S.; Ådnøy, T. Casein SNP in Norwegian goats: Additive and dominance effects on milk composition and quality. Genet. Sel. Evol. 2011, 43, 31. [Google Scholar] [CrossRef] [PubMed]
- Dettori, M.L.; Pazzola, M.; Paschino, P.; Amills, M.; Vacca, G.M. Association between the GHR, GHRHR, and IGF1 gene polymorphisms and milk yield and quality traits in Sarda sheep. J. Dairy Sci. 2018, 101, 9978–9986. [Google Scholar] [CrossRef] [PubMed]
- Du, A.; Zhao, F.; Liu, Y.; Xu, L.; Chen, K.; Sun, D.; Han, B. Genetic polymorphisms of PKLR gene and their associations with milk production traits in Chinese Holstein cows. Front. Genet. 2022, 13, 1002706. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Shi, L.; Peng, P.; Han, B.; Liu, L.; Lv, X.; Ma, Z.; Zhang, S.; Sun, D. Determination of genetic effects and functional SNPs of bovine HTR1B gene on milk fatty acid traits. BMC Genom. 2021, 22, 575. [Google Scholar] [CrossRef]
- Hou, Y.; Xie, Y.; Yang, S.; Han, B.; Shi, L.; Bai, X.; Liang, R.; Dong, T.; Zhang, S.; Zhang, Q. EEF1D facilitates milk lipid synthesis by regulation of PI3K-Akt signaling in mammals. FASEB J. 2021, 35, e21455. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhao, Y.; Iqbal, A.; Xia, L.; Bai, Z.; Sun, H.; Fang, X.; Yang, R.; Zhao, Z. Effects of polymorphism of the GPAM gene on milk quality traits and its relation to triglyceride metabolism in bovine mammary epithelial cells of dairy cattle. Arch. Anim. Breed. 2021, 64, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lu, X.; Gao, Q.; Wang, M.; Arbab, A.A.I.; Sun, Y.; Chen, Z.; Zhang, H.; Karrow, N.A.; Yang, Z. A functional 3′ utr polymorphism of fads2 affects cow milk composition through modifying mir-744 binding. Animals 2019, 9, 1090. [Google Scholar] [CrossRef] [PubMed]
- Proskura, W.S.; Liput, M.; Zaborski, D.; Sobek, Z.; Yu, Y.-H.; Cheng, Y.-H.; Dybus, A. The effect of polymorphism in the FADS2 gene on the fatty acid composition of bovine milk. Arch. Anim. Breed. 2019, 62, 547–555. [Google Scholar] [CrossRef]
- Li, R.; Ma, Y.; Jiang, L. Research progress of dairy sheep milk genes. Agriculture 2022, 12, 169. [Google Scholar] [CrossRef]
- Routtu, J.; Hall, M.D.; Albere, B.; Beisel, C.; Bergeron, R.D.; Chaturvedi, A.; Choi, J.H.; Colbourne, J.; De Meester, L.; Stephens, M.T.; et al. An SNP-based second-generation genetic map of Daphnia magna and its application to QTL analysis of phenotypic traits. BMC Genom. 2014, 15, 1033. [Google Scholar] [CrossRef] [PubMed]
- Akey, J.; Jin, L.; Xiong, M. Haplotypes vs single marker linkage disequilibrium tests: What do we gain? Eur. J. Hum. Genet. EJHG 2001, 9, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.R.; Lai, E.H.; Gilbert, J.R.; Rogala, A.R.; Afshari, A.J.; Riley, J.; Finch, K.L.; Stevens, J.F.; Livak, K.J.; Slotterbeck, B.D.; et al. SNPing away at complex diseases: Analysis of single-nucleotide polymorphisms around APOE in Alzheimer disease. Am. J. Hum. Genet. 2000, 67, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Stephens, J.C.; Schneider, J.A.; Tanguay, D.A.; Choi, J.; Acharya, T.; Stanley, S.E.; Jiang, R.; Messer, C.J.; Chew, A.; Han, J.H.; et al. Haplotype variation and linkage disequilibrium in 313 human genes. Science 2001, 293, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yuan, X.; Li, P.; Wang, Y.; Yu, M.; Gao, X. Vaccarin promotes proliferation of and milk synthesis in bovine mammary epithelial cells through the Prl receptor-PI3K signaling pathway. Eur. J. Pharmacol. 2020, 880, 173190. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| SNPs | Position | Genotypic Frequencies | Allelic Frequencies | He | Ne | PIC | HWpval | MAF | Alleles | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| g.285,808 | Intron | AA | TA | TT | A | T | 0.483 | 1.934 | 0.366 | 0.772 | 0.407 | T:(A) |
| 0.160 | 0.494 | 0.346 | 0.407 | 0.593 | ||||||||
| g.285,847 | Intron | CC | TC | CC | C | T | 0.436 | 1.773 | 0.341 | 0.911 | 0.321 | C:(T) |
| 0.463 | 0.432 | 0.105 | 0.679 | 0.321 | ||||||||
| g.285,867 | Intron | AA | CA | CC | A | C | 0.477 | 1.911 | 0.363 | 0.533 | 0.392 | C:(A) |
| 0.142 | 0.500 | 0.358 | 0.392 | 0.608 | ||||||||
| g.285,916 | Intron | GG | TG | TT | G | T | 0.477 | 1.911 | 0.363 | 0.533 | 0.392 | T:(G) |
| 0.142 | 0.500 | 0.358 | 0.392 | 0.608 | ||||||||
| g.285,952 | Intron | AA | AG | GG | A | G | 0.477 | 1.911 | 0.363 | 0.533 | 0.392 | A:(G) |
| 0.358 | 0.500 | 0.142 | 0.608 | 0.392 | ||||||||
| g.285,965 | Intron | CC | TC | TT | C | T | 0.483 | 1.934 | 0.366 | 0.772 | 0.407 | C:(T) |
| 0.346 | 0.494 | 0.160 | 0.593 | 0.407 | ||||||||
| g.306,600 | Intron | CC | TC | TT | C | T | 0.324 | 1.480 | 0.272 | 0.404 | 0.204 | T:(C) |
| 0.031 | 0.346 | 0.629 | 0.204 | 0.796 | ||||||||
| g.306,635 | Intron | CC | CT | TT | C | T | 0.422 | 1.730 | 0.333 | 0.498 | 0.302 | C:(T) |
| 0.475 | 0.444 | 0.080 | 0.698 | 0.302 | ||||||||
| g.315,413 | Intron | CC | CT | TT | C | T | 0.324 | 1.480 | 0.272 | 0.404 | 0.204 | C:(T) |
| 0.623 | 0.346 | 0.031 | 0.796 | 0.204 | ||||||||
| SNP g.285,808T>A | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| AA | 4.25 ± 0.22 Aa | 5.09 ± 0.31 Aa | 6.09 ± 3.2 | 11.47 ± 0.52 a | 4.99 ± 0.16 | 12.79 ± 1.12 | 17.44 ± 3.05 a |
| TA | 4.12 ± 0.29 Ab | 4.92 ± 0.41 ABb | 5.87 ± 2.65 | 11.26 ± 0.48 ab | 4.97 ± 0.17 | 12.4 ± 1.4 | 17.03 ± 2.58 a |
| TT | 3.97 ± 0.28 Bc | 4.78 ± 0.37 Bb | 4.88 ± 2.25 | 11.18 ± 0.42 b | 5 ± 0.14 | 12.18 ± 1.17 | 15.91 ± 2.18 b |
| p-Value | p = 0.000 | p = 0.003 | p = 0.052 | p = 0.037 | p = 0.619 | p = 0.137 | p = 0.013 |
| SNPs g.285,847T>C | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| CC | 4.01 ± 0.28 Bb | 4.82 ± 0.37 Bb | 5.15 ± 2.3 | 11.22 ± 0.41 b | 5 ± 0.15 | 12.24 ± 1.19 b | 16.23 ± 2.26 |
| TC | 4.13 ± 0.3 ABa | 4.93 ± 0.41 ABab | 6.07 ± 2.97 | 11.25 ± 0.53 b | 4.96 ± 0.17 | 12.38 ± 1.41 b | 17.22 ± 2.87 |
| TT | 4.23 ± 0.24 Aa | 5.13 ± 0.32 Aa | 5.33 ± 2.49 | 11.53 ± 0.44 a | 4.98 ± 0.13 | 13.1 ± 0.99 a | 16.72 ± 2.46 |
| p-Value | p = 0.003 | p = 0.009 | p = 0.104 | p = 0.043 | p = 0.342 | p = 0.044 | p = 0.068 |
| SNPs g.285,867C>A | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| AA | 3.95 ± 0.31 Bb | 4.74 ± 0.45 B | 4.97 ± 2.58 | 11.09 ± 0.54 | 5.01 ± 0.16 | 12.15 ± 1.3 | 15.96 ± 2.39 |
| CA | 4.07 ± 0.27 ABb | 4.88 ± 0.36 AB | 5.43 ± 2.38 | 11.26 ± 0.4 | 4.99 ± 0.17 | 12.37 ± 1.25 | 16.56 ± 2.35 |
| CC | 4.17 ± 0.3 Aa | 4.99 ± 0.4 A | 5.99 ± 2.98 | 11.34 ± 0.54 | 4.96 ± 0.13 | 12.51 ± 1.35 | 17.21 ± 2.9 |
| p-Value | p = 0.006 | p = 0.023 | p = 0.240 | p = 0.096 | p = 0.356 | p = 0.527 | p = 0.113 |
| SNPs g.285,916T>G | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| GG | 3.95 ± 0.31 Bb | 4.74 ± 0.45 B | 4.97 ± 2.58 | 11.09 ± 0.54 | 5.01 ± 0.16 | 12.15 ± 1.3 | 15.96 ± 2.39 |
| TG | 4.07 ± 0.27 ABb | 4.88 ± 0.36 AB | 5.43 ± 2.38 | 11.26 ± 0.4 | 4.99 ± 0.17 | 12.37 ± 1.25 | 16.56 ± 2.35 |
| TT | 4.17 ± 0.3 Aa | 4.99 ± 0.4 A | 5.99 ± 2.98 | 11.34 ± 0.54 | 4.96 ± 0.13 | 12.51 ± 1.35 | 17.21 ± 2.9 |
| p-Value | p = 0.006 | p = 0.023 | p = 0.240 | p = 0.096 | p = 0.356 | p = 0.527 | p = 0.113 |
| SNPs g.285,952A>G | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| AA | 4.17 ± 0.3 Aa | 4.99 ± 0.4 A | 5.99 ± 2.98 | 11.34 ± 0.54 | 4.96 ± 0.13 | 12.51 ± 1.35 | 17.21 ± 2.9 |
| AG | 4.07 ± 0.27 ABb | 4.88 ± 0.36 AB | 5.43 ± 2.38 | 11.26 ± 0.4 | 4.99 ± 0.17 | 12.37 ± 1.25 | 16.56 ± 2.35 |
| GG | 3.95 ± 0.31 Bb | 4.74 ± 0.45 B | 4.97 ± 2.58 | 11.09 ± 0.54 | 5.01 ± 0.16 | 12.15 ± 1.3 | 15.96 ± 2.39 |
| p-Value | p = 0.006 | p = 0.023 | p = 0.240 | p = 0.096 | p = 0.356 | p = 0.527 | p = 0.113 |
| SNPs g.285,965T>C | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| CC | 3.97 ± 0.28 Bc | 4.78 ± 0.37 Bc | 4.88 ± 2.25 | 11.18 ± 0.42 b | 5 ± 0.14 | 12.18 ± 1.17 | 15.91 ± 2.18 b |
| TC | 4.12 ± 0.29 Ab | 4.92 ± 0.41 ABb | 5.87 ± 2.65 | 11.26 ± 0.48 ab | 4.97 ± 0.17 | 12.4 ± 1.4 | 17.03 ± 2.58 a |
| TT | 4.25 ± 0.22 Aa | 5.09 ± 0.31 Aa | 6.09 ± 3.2 | 11.47 ± 0.52 a | 4.99 ± 0.16 | 12.79 ± 1.12 | 17.44 ± 3.05 a |
| p-Value | p = 0.000 | p = 0.003 | p = 0.052 | p = 0.037 | p = 0.619 | p = 0.137 | p = 0.013 |
| SNPs g.306,600T>C | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| CC | 4.1 ± 0.21 AB | 4.86 ± 0.16 AB | 6.13 ± 3.57 | 11.14 ± 0.32 AB | 5.05 ± 0.15 | 12.3 ± 0.27 | 17.28 ± 3.57 ab |
| TC | 4.22 ± 0.26 A | 5.06 ± 0.38 A | 6.1 ± 2.88 | 11.4 ± 0.47 A | 4.96 ± 0.17 | 12.64 ± 1.27 | 17.4 ± 2.77 a |
| TT | 4.01 ± 0.29 B | 4.81 ± 0.39 B | 5.24 ± 2.44 | 11.19 ± 0.47 B | 4.99 ± 0.15 | 12.26 ± 1.32 | 16.3 ± 2.36 b |
| p-Value | p = 0.000 | p = 0.001 | p = 0.130 | p = 0.025 | p = 0.363 | p = 0.202 | p = 0.033 |
| SNPs g.306,635C>T | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| CC | 4.15 ± 0.28 Aa | 4.99 ± 0.37 Aa | 5.77 ± 2.84 | 11.33 ± 0.47 a | 4.96 ± 0.14 | 12.44 ± 1.32 | 16.96 ± 2.79 |
| CT | 4.07 ± 0.28 Aa | 4.86 ± 0.4 Aa | 5.44 ± 2.53 | 11.25 ± 0.47 a | 5 ± 0.17 | 12.43 ± 1.3 | 16.57 ± 2.44 |
| TT | 3.84 ± 0.27 Bb | 4.6 ± 0.37 Bb | 5.06 ± 2.1 | 10.97 ± 0.47 b | 5.01 ± 0.13 | 11.9 ± 1.04 | 15.95 ± 1.98 |
| p-Value | p = 0.001 | p = 0.002 | p = 0.581 | p = 0.036 | p = 0.143 | p = 0.370 | p = 0.356 |
| SNPs g.315,413C>T | |||||||
| Genotype | Casein/% | Protein/% | Fat/% | SNF/% | Lactose/% | Acidity/°T | TS/% |
| CC | 4.01 ± 0.29 B | 4.81 ± 0.39 B | 5.24 ± 2.44 | 11.19 ± 0.47 B | 4.99 ± 0.15 | 12.26 ± 1.32 | 16.3 ± 2.36 b |
| CT | 4.22 ± 0.26 A | 5.06 ± 0.38 A | 6.1 ± 2.88 | 11.4 ± 0.47 A | 4.96 ± 0.17 | 12.64 ± 1.27 | 17.4 ± 2.77 a |
| TT | 4.1 ± 0.21 AB | 4.86 ± 0.16 AB | 6.13 ± 3.57 | 11.14 ± 0.32 AB | 5.05 ± 0.15 | 12.3 ± 0.27 | 17.28 ± 3.57 ab |
| p-Value | p = 0.000 | p = 0.001 | p = 0.130 | p = 0.025 | p = 0.363 | p = 0.202 | p = 0.033 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Zha, X.; Ma, X.; La, Y.; Guo, X.; Chu, M.; Bao, P.; Yan, P.; Wu, X.; Liang, C. Polymorphisms of ITGA9 Gene and Their Correlation with Milk Quality Traits in Yak (Bos grunniens). Foods 2024, 13, 1613. https://doi.org/10.3390/foods13111613
Zhang M, Zha X, Ma X, La Y, Guo X, Chu M, Bao P, Yan P, Wu X, Liang C. Polymorphisms of ITGA9 Gene and Their Correlation with Milk Quality Traits in Yak (Bos grunniens). Foods. 2024; 13(11):1613. https://doi.org/10.3390/foods13111613
Chicago/Turabian StyleZhang, Mengfan, Xita Zha, Xiaoming Ma, Yongfu La, Xian Guo, Min Chu, Pengjia Bao, Ping Yan, Xiaoyun Wu, and Chunnian Liang. 2024. "Polymorphisms of ITGA9 Gene and Their Correlation with Milk Quality Traits in Yak (Bos grunniens)" Foods 13, no. 11: 1613. https://doi.org/10.3390/foods13111613
APA StyleZhang, M., Zha, X., Ma, X., La, Y., Guo, X., Chu, M., Bao, P., Yan, P., Wu, X., & Liang, C. (2024). Polymorphisms of ITGA9 Gene and Their Correlation with Milk Quality Traits in Yak (Bos grunniens). Foods, 13(11), 1613. https://doi.org/10.3390/foods13111613