


Characterization of Selected Microalgae Species as Potential Sources of Nutrients and Antioxidants
 ,
,  ,
,  , ,
, , 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Protein Content
2.3. Amino Acid Composition
2.4. Determination of Mineral Profile
2.5. Fatty Acids Composition
2.6. Methanolic Extraction
2.7. Polyphenols Profile Composition
2.8. Analysis of B Vitamins
2.9. Antioxidant Activity
2.10. Statistical Analysis
3. Results and Discussion
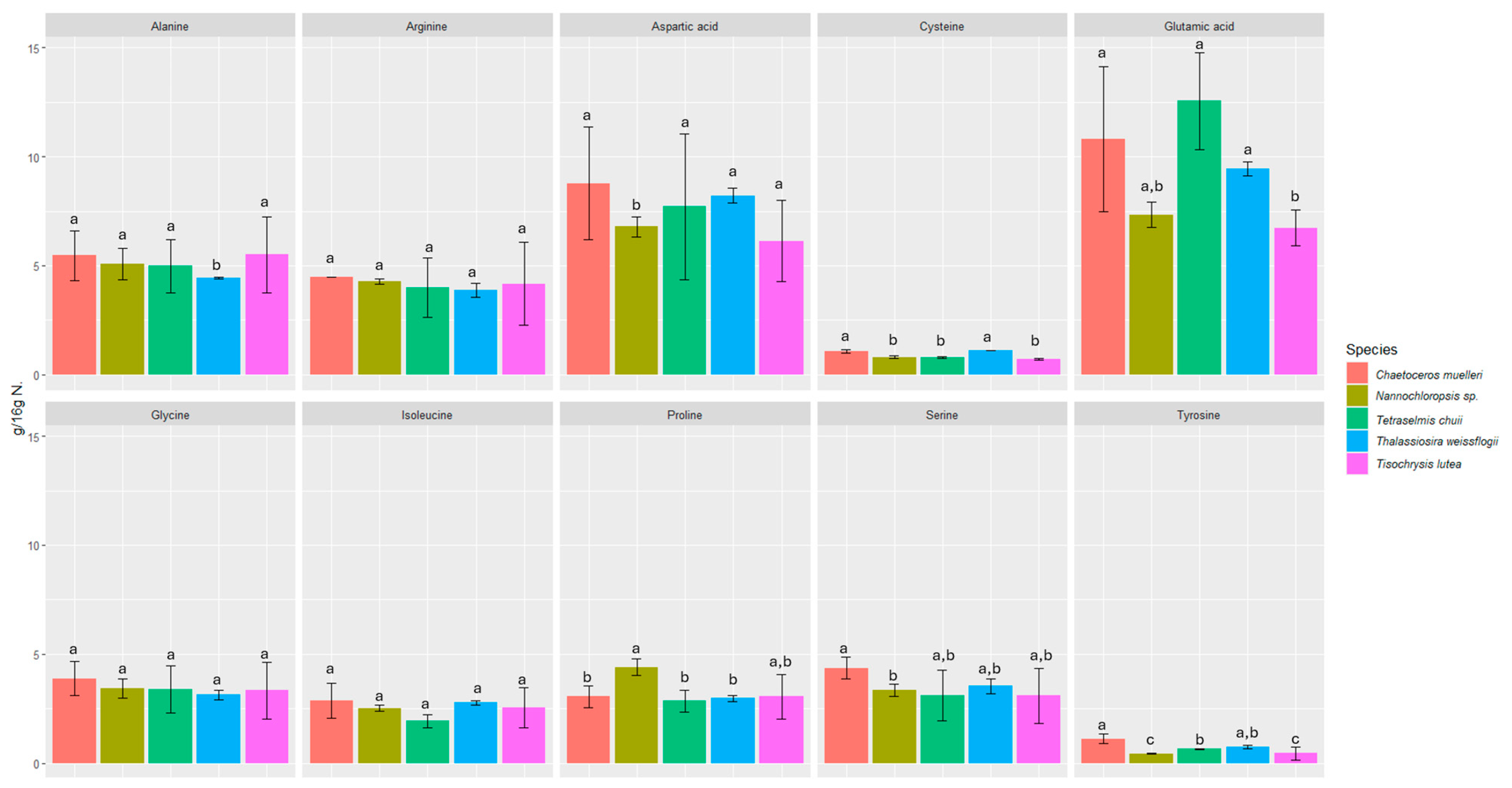
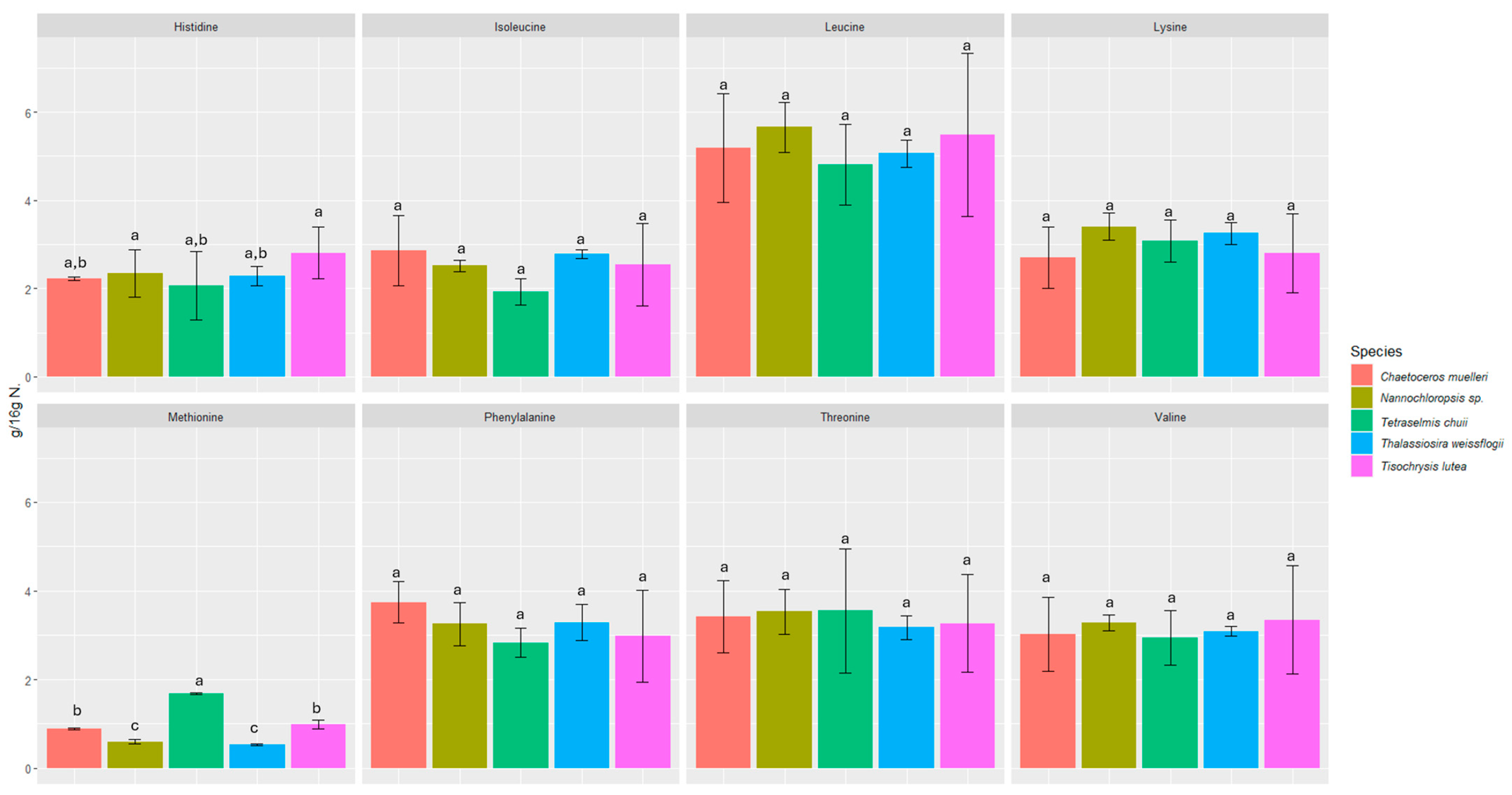
3.1. Protein and Amino Acid Content
3.2. Mineral and Trace Element Content
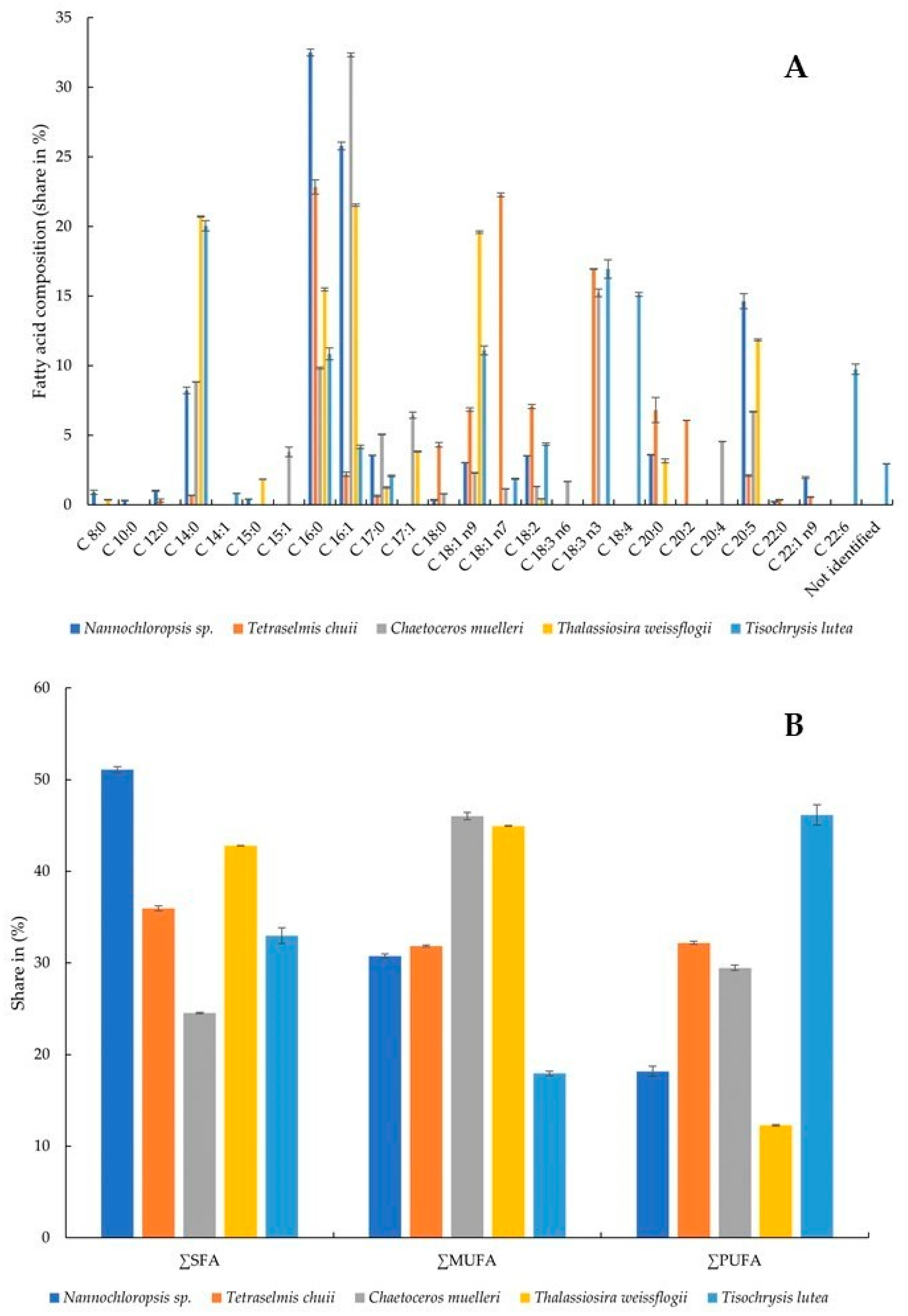
3.3. Fatty Acid Composition
3.4. Composition of Polyphenols
3.5. Content of B Vitamins
3.6. Antioxidant Activity
3.6.1. DPPH Assay
3.6.2. ABTS Assay
3.7. Nutritional Use of Algae in the Food Industry
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a Future Food Source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef] [PubMed]
- UNICEF. The State of Food Security and Nutrition in the World 2019: Safeguarding Against Economic Slowdowns and Downturns. 2019. Available online: https://www.sidalc.net/search/Record/unfao:852712/Description (accessed on 3 April 2024).
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as Nutritional and Functional Food Sources: Revisiting Our Understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Zanella, L.; Vianello, F. Microalgae of the Genus Nannochloropsis: Chemical Composition and Functional Implications for Human Nutrition. J. Funct. Foods 2020, 68, 103919. [Google Scholar] [CrossRef]
- Wijffels, R.H.; Barbosa, M.J.; Eppink, M.H.M. Microalgae for the Production of Bulk Chemicals and Biofuels. Biofuels Bioprod. Biorefining 2010, 4, 287–295. [Google Scholar] [CrossRef]
- Wijffels, R.H.; Kruse, O.; Hellingwerf, K.J. Potential of Industrial Biotechnology with Cyanobacteria and Eukaryotic Microalgae. Curr. Opin. Biotechnol. 2013, 24, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A Novel Ingredient in Nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- Bleakley, S.; Hayes, M. Algal Proteins: Extraction, Application, and Challenges Concerning Production. Foods 2017, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Draaisma, R.B.; Wijffels, R.H.; Slegers, P.E.; Brentner, L.B.; Roy, A.; Barbosa, M.J. Food Commodities from Microalgae. Curr. Opin. Biotechnol. 2013, 24, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Delgado, C.L. Rising Consumption of Meat and Milk in Developing Countries Has Created a New Food Revolution. J. Nutr. 2003, 133, 3907S–3910S. [Google Scholar] [CrossRef]
- Wang, Y.; Tibbetts, S.M.; McGinn, P.J. Microalgae as Sources of High-Quality Protein for Human Food and Protein Supplements. Foods 2021, 10, 3002. [Google Scholar] [CrossRef]
- García, J.L.; De Vicente, M.; Galán, B. Microalgae, Old Sustainable Food and Fashion Nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef]
- Matos, Â.P. Chapter 3—Microalgae as a Potential Source of Proteins. In Proteins: Sustainable Source, Processing and Applications; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 63–96. ISBN 978-0-12-816695-6. [Google Scholar]
- Matos, Â.P. The Impact of Microalgae in Food Science and Technology. J. Am. Oil Chem. Soc. 2017, 94, 1333–1350. [Google Scholar] [CrossRef]
- FAO; WHO; UNU. Energy and Protein Requirements: Report of a Joint FAO/WHO/UNU Expert Consultation; World Health Organization: Geneva, Switzerland, 1973; ISBN 92-4-120522-9. [Google Scholar]
- Becker, E.W. Microalgae for human and animal nutrition. In Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 2nd ed.; Richmond, A., Hu, Q., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 461–503. [Google Scholar]
- Santiago-Díaz, P.; Rivero, A.; Rico, M.; Gómez-Pinchetti, J.L. Characterization of Novel Selected Microalgae for Antioxidant Activity and Polyphenols, Amino Acids, and Carbohydrates. Mar. Drugs 2022, 20, 40. [Google Scholar] [CrossRef]
- Rupérez, P. Mineral Content of Edible Marine Seaweeds. Food Chem. 2002, 79, 23–26. [Google Scholar] [CrossRef]
- Mabeau, S.; Fleurence, J. Seaweed in Food Products: Biochemical and Nutritional Aspects. Trends Food Sci. Technol. 1993, 4, 103–107. [Google Scholar] [CrossRef]
- Ortega-Calvo, J.J.; Mazuelos, C.; Hermosin, B.; Sáiz-Jiménez, C. Chemical Composition of Spirulina and Eukaryotic Algae Food Products Marketed in Spain. J. Appl. Phycol. 1993, 5, 425–435. [Google Scholar] [CrossRef]
- Ito, K.; Hori, K. Seaweed: Chemical Composition and Potential Food Uses. Food Rev. Int. 1989, 5, 101–144. [Google Scholar] [CrossRef]
- Nisizawa, K.; Noda, H.; Kikuchi, R.; Watanabe, T. The Main Seaweed Foods in Japan. Hydrobiologia 1987, 151, 5–29. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K.; Saeid, A. Plant Growth Biostimulants, Dietary Feed Supplements and Cosmetics Formulated with Supercritical CO2 Algal Extracts. Molecules 2017, 22, 66. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algal Extracts: Technology and Advances. Eng. Life Sci. 2014, 14, 581–591. [Google Scholar] [CrossRef]
- Rao, A.R.; Sarada, R.; Baskaran, V.; Ravishankar, G.A. Antioxidant Activity of Botryococcus Braunii Extract Elucidated In Vitro Models. J. Agric. Food Chem. 2006, 54, 4593–4599. [Google Scholar] [CrossRef]
- Jahnke, L.S. Massive Carotenoid Accumulation in Dunaliella Bardawil Induced by Ultraviolet-A Radiation. J. Photochem. Photobiol. B 1999, 48, 68–74. [Google Scholar] [CrossRef]
- Natrah, F.; Yusoff, F.; Shariff, M.; Abas, F.; Mariana, N. Screening of Malaysian Indigenous Microalgae for Antioxidant Properties and Nutritional Value. J. Appl. Phycol. 2007, 19, 711–718. [Google Scholar] [CrossRef]
- Safafar, H.; Van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, Phenolic Compounds and Tocopherols Contribute to the Antioxidative Properties of Some Microalgae Species Grown on Industrial Wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef]
- Del Mondo, A.; Smerilli, A.; Sané, E.; Sansone, C.; Brunet, C. Challenging Microalgal Vitamins for Human Health. Microb. Cell Factories 2020, 19, 201. [Google Scholar] [CrossRef]
- Smith, A.G.; Croft, M.T.; Moulin, M.; Webb, M.E. Plants Need Their Vitamins Too. Curr. Opin. Plant Biol. 2007, 10, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Udayan, A.; Arumugam, M.; Pandey, A. Nutraceuticals from Algae and Cyanobacteria. In Algal Green Chemistry; Elsevier: Amsterdam, The Netherlands, 2017; pp. 65–89. [Google Scholar]
- Grusak, M.A.; DellaPenna, D. Improving the Nutrient Composition of Plants to Enhance Human Nutrition And Health. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 133–161. [Google Scholar] [CrossRef] [PubMed]
- Day, L. Proteins from Land Plants—Potential Resources for Human Nutrition and Food Security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]
- Yaqoob, P. Role of Lipids in Human Nutrition. In Handbook of Olive Oil: Analysis and Properties; Aparicio, R., Harwood, J., Eds.; Springer: Boston, MA, USA, 2013; pp. 655–675. ISBN 978-1-4614-7777-8. [Google Scholar]
- Gharibzahedi, S.M.T.; Jafari, S.M. The Importance of Minerals in Human Nutrition: Bioavailability, Food Fortification, Processing Effects and Nanoencapsulation. Trends Food Sci. Technol. 2017, 62, 119–132. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An Overview of Plant Phenolic Compounds and Their Importance in Human Nutrition and Management of Type 2 Diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Wan, Z.; Zheng, J.; Zhu, Z.; Sang, L.; Zhu, J.; Luo, S.; Zhao, Y.; Wang, R.; Zhang, Y.; Hao, K.; et al. Intermediate Role of Gut Microbiota in Vitamin B Nutrition and Its Influences on Human Health. Front. Nutr. 2022, 9, 1031502. [Google Scholar] [CrossRef] [PubMed]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar. Drugs 2021, 19, 549. [Google Scholar] [CrossRef]
- ISO 20483:2013; Cereals and Pulses—Determination of the Nitrogen Content and Calculation of the Crude Protein Content—Kjeldahl Method. International Organization for Standardization: Geneva, Switzerland, 2013. Available online: https://www.iso.org/standard/59162.html (accessed on 4 March 2024).
- AOAC Official Method 994.12 Amino Acids in Feeds|PDF|Amino Acid|Ph. Available online: https://www.scribd.com/document/609975831/Aoac-Official-Method-994-12-Amino-Acids-in-Feeds-1 (accessed on 4 March 2024).
- Tomczak, A.; Zielińska-Dawidziak, M.; Piasecka-Kwiatkowska, D.; Lampart-Szczapa, E. Blue Lupine Seeds Protein Content and Amino Acids Composition. Plant Soil Environ. 2018, 64, 147–155. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for The Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- American Oil Chemists’ Society. AOCS Official Method Ce 1h-05: Determination of Cis-, Trans-, Saturated, Monounsaturated and Polyunsaturated Fatty Acids in Vegetable or Non-ruminant Animal Oils and Fats by Capillary GLC. In Official Methods and Recommended Practices of the AOCS; AOCS: Urbana, IL, USA, 2005. [Google Scholar]
- Kowalczewski, P.Ł.; Gumienna, M.; Rybicka, I.; Górna, B.; Sarbak, P.; Dziedzic, K.; Kmiecik, D. Nutritional Value and Biological Activity of Gluten-Free Bread Enriched with Cricket Powder. Molecules 2021, 26, 1184. [Google Scholar] [CrossRef] [PubMed]
- Drożdżyńska, A.; Dzidzic, K.; Kośmider, A.; Leja, K.; Czaczyk, K.; Górecka, D. Application of Fast Liquid Chromatography for Antioxidants Analysis. Acta Sci. Pol. Technol. Aliment. 2012, 11, 19–25. [Google Scholar] [PubMed]
- Li, H.-B.; Chen, F. Simultaneous Determination of Nine Water-Soluble Vitamins in Pharmaceutical Preparations by High-Performance Liquid Chromatography with Diode Array Detection. J. Sep. Sci. 2001, 24, 271–274. [Google Scholar] [CrossRef]
- Kačániová, M.; Vukovic, N.L.; Čmiková, N.; Galovičová, L.; Schwarzová, M.; Šimora, V.; Kowalczewski, P.Ł.; Kluz, M.I.; Puchalski, C.; Bakay, L.; et al. Salvia Sclarea Essential Oil Chemical Composition and Biological Activities. Int. J. Mol. Sci. 2023, 24, 5179. [Google Scholar] [CrossRef]
- Andreeva, A.; Budenkova, E.; Babich, O.; Sukhikh, S.; Ulrikh, E.; Ivanova, S.; Prosekov, A.; Dolganyuk, V. Production, Purification, and Study of the Amino Acid Composition of Microalgae Proteins. Molecules 2021, 26, 2767. [Google Scholar] [CrossRef]
- Hulatt, C.J.; Wijffels, R.H.; Bolla, S.; Kiron, V. Production of Fatty Acids and Protein by Nannochloropsis in Flat-Plate Photobioreactors. PLoS ONE 2017, 12, e0170440. [Google Scholar] [CrossRef]
- Rebolloso-Fuentes, M.M.; Navarro-Pérez, A.; García-Camacho, F.; Ramos-Miras, J.J.; Guil-Guerrero, J.L. Biomass Nutrient Profiles of the Microalga Nannochloropsis. J. Agric. Food Chem. 2001, 49, 2966–2972. [Google Scholar] [CrossRef]
- Khatoon, H.; Haris, H.; Rahman, N.A.; Zakaria, M.N.; Begum, H.; Mian, S. Growth, Proximate Composition and Pigment Production of Tetraselmis Chuii Cultured with Aquaculture Wastewater. J. Ocean Univ. China 2018, 17, 641–646. [Google Scholar] [CrossRef]
- Brown, M.R. The Amino-Acid and Sugar Composition of 16 Species of Microalgae Used in Mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- Wang, X.-W.; Liang, J.-R.; Luo, C.-S.; Chen, C.-P.; Gao, Y.-H. Biomass, Total Lipid Production, and Fatty Acid Composition of the Marine Diatom Chaetoceros Muelleri in Response to Different CO2 Levels. Bioresour. Technol. 2014, 161, 124–130. [Google Scholar] [CrossRef]
- Iqbal, A.; Khalil, I.A.; Ateeq, N.; Sayyar Khan, M. Nutritional Quality of Important Food Legumes. Food Chem. 2006, 97, 331–335. [Google Scholar] [CrossRef]
- Cho, K.S. Chemical Compositions in the Various Plant Types of Peanut (Arachis Hypogaea L.). J. Korean Soc. Food Nutr. Korea Repub. 1993, 22, 576–580. [Google Scholar]
- Brown, M.R.; Jeffrey, S.W. Biochemical Composition of Microalgae from the Green Algal Classes Chlorophyceae and Prasinophyceae. 1. Amino Acids, Sugars and Pigments. J. Exp. Mar. Biol. Ecol. 1992, 161, 91–113. [Google Scholar] [CrossRef]
- Araya, M.; García, S.; Rengel, J.; Pizarro, S.; Álvarez, G. Determination of Free and Protein Amino Acid Content in Microalgae by HPLC-DAD with Pre-Column Derivatization and Pressure Hydrolysis. Mar. Chem. 2021, 234, 103999. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical Composition and Nutritional Properties of Freshwater and Marine Microalgal Biomass Cultured in Photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef]
- Tokuşoglu, Ö.; Üunal, M.K. Biomass Nutrient Profiles of Three Microalgae: Spirulina Platensis, Chlorella Vulgaris, and Isochrisis Galbana. J. Food Sci. 2003, 68, 1144–1148. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K. Fatty Acids of Microalgae: Diversity and Applications. Rev. Environ. Sci. Biotechnol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Servel, M.-O.; Claire, C.; Derrien, A.; Coiffard, L.; De Roeck-Holtzhauer, Y. Fatty Acid Composition of Some Marine Microalgae. Phytochemistry 1994, 36, 691–693. [Google Scholar] [CrossRef]
- Sahu, A.; Pancha, I.; Jain, D.; Paliwal, C.; Ghosh, T.; Patidar, S.; Bhattacharya, S.; Mishra, S. Fatty Acids as Biomarkers of Microalgae. Phytochemistry 2013, 89, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, L.; Oliveira, A.C. Microalgae as a Raw Material for Biofuels Production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Zolotariova, Y.K.; Mokrosnop, V.M.; Stepanov, S.S. Polyphenol Compounds of Macroscopic and Microscopic Algae. Int. J. Algae 2019, 21, 3–29. [Google Scholar] [CrossRef]
- Scaglioni, P.T.; Quadros, L.; de Paula, M.; Furlong, V.B.; Abreu, P.C.; Badiale-Furlong, E. Inhibition of Enzymatic and Oxidative Processes by Phenolic Extracts from Spirulina Sp. and Nannochloropsis Sp. Food Technol. Biotechnol. 2018, 56, 344. [Google Scholar] [CrossRef] [PubMed]
- Haoujar, I.; Cacciola, F.; Abrini, J.; Mangraviti, D.; Giuffrida, D.; Oulad El Majdoub, Y.; Kounnoun, A.; Miceli, N.; Fernanda Taviano, M.; Mondello, L.; et al. The Contribution of Carotenoids, Phenolic Compounds, and Flavonoids to the Antioxidative Properties of Marine Microalgae Isolated from Mediterranean Morocco. Molecules 2019, 24, 4037. [Google Scholar] [CrossRef]
- Yıldırım, Z.; İlk, Y.; Yıldırım, M. The Synergist Effect of P-Hydroxybenzoic Acid and Propyl-Paraben on the Antibacterial Activity of Enterocin KP. Turk. J. Agric. Food Sci. Technol. 2014, 2, 1–5. [Google Scholar] [CrossRef]
- Zhao, Z.; Xue, W.; Wang, J.; Zhang, C.; Zhou, D. The Role of Trace P-Hydroxybenzoic Acid to Chlorella for Advanced Wastewater Treatment: Mitigating Bacterial Contamination and Boosting Biomass Recovery. Resour. Conserv. Recycl. 2023, 199, 107229. [Google Scholar] [CrossRef]
- Edelmann, M.; Aalto, S.; Chamlagain, B.; Kariluoto, S.; Piironen, V. Riboflavin, Niacin, Folate and Vitamin B12 in Commercial Microalgae Powders. J. Food Compos. Anal. 2019, 82, 103226. [Google Scholar] [CrossRef]
- Bishop, W.M.; Zubeck, H.M. Evaluation of Microalgae for Use as Nutraceuticals and Nutritional Supplements. J. Nutr. Food Sci. 2012, 2, 1000147. [Google Scholar] [CrossRef]
- Brown, M.R.; Mular, M.; Miller, I.; Farmer, C.; Trenerry, C. The Vitamin Content of Microalgae Used in Aquaculture. J. Appl. Phycol. 1999, 11, 247–255. [Google Scholar] [CrossRef]
- Paterson, S.; Gómez-Cortés, P.; de la Fuente, M.A.; Hernández-Ledesma, B. Bioactivity and Digestibility of Microalgae Tetraselmis Sp. and Nannochloropsis Sp. as Basis of Their Potential as Novel Functional Foods. Nutrients 2023, 15, 477. [Google Scholar] [CrossRef] [PubMed]
- Choochote, W.; Suklampoo, L.; Ochaikul, D. Evaluation of Antioxidant Capacities of Green Microalgae. J. Appl. Phycol. 2014, 26, 43–48. [Google Scholar] [CrossRef]
- Hajimahmoodi, M.; Faramarzi, M.A.; Mohammadi, N.; Soltani, N.; Oveisi, M.R.; Nafissi-Varcheh, N. Evaluation of Antioxidant Properties and Total Phenolic Contents of Some Strains of Microalgae. J. Appl. Phycol. 2010, 22, 43–50. [Google Scholar] [CrossRef]
- Sousa, I.; Gouveia, L.; Batista, A.P.; Raymundo, A.; Bandarra, N.M. Microalgae in Novel Food Products. Food Chem. Res. Dev. 2008, 75, 75–112. [Google Scholar]
- Batista, A.P.; Raymundo, A.; Sousa, I.; Empis, J. Rheological Characterization of Coloured Oil-in-Water Food Emulsions with Lutein and Phycocyanin Added to the Oil and Aqueous Phases. Food Hydrocoll. 2006, 20, 44–52. [Google Scholar] [CrossRef]
- Ferreira, A.; Guerra, I.; Costa, M.; Silva, J.; Gouveia, L. Chapter 15—Future Perspectives of Microalgae in the Food Industry. In Cultured Microalgae for the Food Industry; Lafarga, T., Acién, G., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 387–433. ISBN 978-0-12-821080-2. [Google Scholar]
- Saharan, V.; Jood, S. Vitamins, Minerals, Protein Digestibility and Antioxidant Activity of Bread Enriched with Spirulina Platensis Powder. Int. J. Agric. Sci. 2017, 9, 3917–3919. [Google Scholar]
- Ak, B.; Avsaroglu, E.; Isik, O.; Özyurt, G.; Kafkas, E.; Etyemez, M. Nutritional and Physicochemical Characteristics of Bread Enriched with Microalgae Spirulina Platensis. Int. J. Eng. Res. Appl. 2016, 6, 30–38. [Google Scholar]
- Khemiri, S.; Khelifi, N.; Nunes, M.C.; Ferreira, A.; Gouveia, L.; Smaali, I.; Raymundo, A. Microalgae Biomass as an Additional Ingredient of Gluten-Free Bread: Dough Rheology, Texture Quality and Nutritional Properties. Algal Res. 2020, 50, 101998. [Google Scholar] [CrossRef]
- Selmo, M.S.; Salas-Mellado, M.M. Technological Quality of Bread from Rice Flour with Spirulina. Int. Food Res. J. 2014, 21, 1523–1528. [Google Scholar]
- Lafarga, T.; Mayre, E.; Echeverria, G.; Viñas, I.; Villaró, S.; Acién-Fernández, F.G.; Castellari, M.; Aguiló-Aguayo, I. Potential of the Microalgae Nannochloropsis and Tetraselmis for Being Used as Innovative Ingredients in Baked Goods. LWT 2019, 115, 108439. [Google Scholar] [CrossRef]
- Fradique, M.; Batista, A.P.; Nunes, M.C.; Gouveia, L.; Bandarra, N.M.; Raymundo, A. Incorporation of Chlorella Vulgaris and Spirulina Maxima Biomass in Pasta Products. Part 1: Preparation and Evaluation. J. Sci. Food Agric. 2010, 90, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Acién-Fernández, F.G.; Castellari, M.; Villaró, S.; Bobo, G.; Aguiló-Aguayo, I. Effect of Microalgae Incorporation on the Physicochemical, Nutritional, and Sensorial Properties of an Innovative Broccoli Soup. LWT 2019, 111, 167–174. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Microalgae | Content (%) |
|---|---|
| Nannochloropsis sp. | 41.68 ± 0.58 ab |
| Tetraselmis chuii | 40.38 ± 0.27 b |
| Chaetoceros muelleri | 34.09 ± 0.39 c |
| Thalassiosira weissflogii | 42.45 ± 0.18 a |
| Tisochrysis lutea | 39.48 ± 1.80 b |
| Mineral | Nannochloropsis sp. | Tetraselmis Chuii | Chaetoceros muelleri | Thalassiosira weissflogii | Tisochrysis lutea |
|---|---|---|---|---|---|
| Ca | 3350.00 ± 270.00 c | 54,900.00 ± 4390.00 a | 3340.00 ± 270.00 c | 2550.00 ± 200.00 d | 4840.00 ± 390.00 b |
| Mg | 3300.00 ± 260.00 c | 7350.00 ± 590.00 a | 7560.00 ± 600.00 a | 4180.00 ± 330.00 bc | 5210.00 ± 420.00 b |
| K | 10,100.00 ± 800.00 c | 12,600.00 ± 1000.00 c | 21,200.00 ± 1700.00 b | 34,700.00 ± 2800.00 a | 11,400.00 ± 900.00 c |
| Na | 13,900.00 ± 1100.00 c | 65,700.00 ± 5300.00 ab | 73,000.00 ± 5800.00 a | 49,900.00 ± 400.00 b | 54,800.00 ± 4400.00 b |
| Cu | 45.10 ± 3.60 c | 78.40 ± 6.20 a | 73.60 ± 5.90 ab | 68.80 ± 5.50 b | 78.50 ± 6.30 a |
| Fe | 605.00 ± 48.00 b | 898.00 ± 72.00 a | 969.00 ± 78.00 a | 580.00 ± 46.00 b | 307.00 ± 25.00 c |
| Mn | 94.50 ± 7.60 b | 139.00 ± 11.00 a | 93.80 ± 7.50 b | 106.00 ± 8.00 ab | 78.90 ± 6.30 c |
| Zn | 93.90 ± 7.50 bc | 106.00 ± 8.50 b | 103.00 ± 8.20 b | 83.5 ± 6.70 c | 181.0 ± 14.00 a |
| Pb | 56.90 ± 4.60 b | 69.50 ± 5.60 b | 94.80 ± 7.60 a | 94.8 ± 7.60 a | 108.0 ± 9.00 a |
| Sample | Nannochloropsis sp. | Tetraselmis chuii | Chaetoceros muelleri | Thalassiosira weissflogii | Tisochrysis lutea |
|---|---|---|---|---|---|
| Kaempferol | 1.47 ± 0.02 b | 3.35 ± 0.05 a | ND | 0.99 ± 0.01 c | ND |
| Vitexin | 1.26 ± 0.03 | ND | ND | ND | ND |
| Rutin | ND | 1.03 ± 0.013 a | ND | ND | 1.07 ± 0.05 a |
| p-Coumaric acid | ND | ND | ND | ND | 0.36 ± 0.06 |
| Catechin | ND | ND | ND | ND | 0.79 ± 0.05 |
| Chlorogenic acid | ND | ND | ND | ND | ND |
| Gallic acid | ND | ND | ND | ND | ND |
| p-Hydroxybenzoic acid | 2.94 ± 0.07 c | 1.61 ± 0.10 d | ND | 10.96 ± 0.11 a | 7.28 ± 0.12 b |
| Sample | Nannochloropsis sp. | Tetraselmis chuii | Chaetoceros muelleri | Thalassiosira weissflogii | Tisochrysis lutea |
|---|---|---|---|---|---|
| Niacin (B3) | 97.81 ± 1.21 d | 161.66 ± 2.04 c | 358.39 ± 2.42 b | 57.57 ± 1.70 e | 514.82 ± 2.95 a |
| Riboflavin (B2) | 2.44 ± 0.12 d | 8.27 ± 0.28 b | 6.94 ± 0.26 c | ND | 20.44 ± 1.96 a |
| Folic acid (B9) | ND | ND | 10.53 ± 0.92 | ND | ND |
| Microalgae | IC50 (mg/mL) | TEAC Equivalent |
|---|---|---|
| Nannochloropsis sp. | 2.01 ± 0.04 a | 0.001475 a |
| Tetraselmis chuii | 1.69 ± 0.02 a | 0.001748 a |
| Chaetoceros muelleri | 0.44 ± 0.01 a | 0.006784 a |
| Thalassiosira weissflogii | 0.92 ± 0.02 a | 0.003229 a |
| Tisochrysis lutea | 1.43 ± 0.02 a | 0.002076 a |
| Microalgae | IC50 (mg/mL) | TEAC Equivalent |
|---|---|---|
| Nannochloropsis sp. | 0.39 ± 0.02 a | 0.006225 a |
| Tetraselmis chuii | 0.17 ± 0.01 b | 0.014891 b |
| Chaetoceros muelleri | 0.21 ± 0.01 b | 0.011897 c |
| Thalassiosira weissflogii | 0.34 ± 0.00 c | 0.007317 d |
| Tisochrysis lutea | 0.38 ± 0.03 a,c | 0.006588 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čmiková, N.; Kowalczewski, P.Ł.; Kmiecik, D.; Tomczak, A.; Drożdżyńska, A.; Ślachciński, M.; Królak, J.; Kačániová, M. Characterization of Selected Microalgae Species as Potential Sources of Nutrients and Antioxidants. Foods 2024, 13, 2160. https://doi.org/10.3390/foods13132160
Čmiková N, Kowalczewski PŁ, Kmiecik D, Tomczak A, Drożdżyńska A, Ślachciński M, Królak J, Kačániová M. Characterization of Selected Microalgae Species as Potential Sources of Nutrients and Antioxidants. Foods. 2024; 13(13):2160. https://doi.org/10.3390/foods13132160
Chicago/Turabian StyleČmiková, Natália, Przemysław Łukasz Kowalczewski, Dominik Kmiecik, Aneta Tomczak, Agnieszka Drożdżyńska, Mariusz Ślachciński, Jakub Królak, and Miroslava Kačániová. 2024. "Characterization of Selected Microalgae Species as Potential Sources of Nutrients and Antioxidants" Foods 13, no. 13: 2160. https://doi.org/10.3390/foods13132160
APA StyleČmiková, N., Kowalczewski, P. Ł., Kmiecik, D., Tomczak, A., Drożdżyńska, A., Ślachciński, M., Królak, J., & Kačániová, M. (2024). Characterization of Selected Microalgae Species as Potential Sources of Nutrients and Antioxidants. Foods, 13(13), 2160. https://doi.org/10.3390/foods13132160