
Non-Targeted Metabolomics Analysis of γ–Aminobutyric Acid Enrichment in Germinated Maize Induced by Pulsed Light


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Surface Sterilization of Seeds
2.3. Pulse Light Treatment
2.4. Germination
2.5. Macromolecule and γ–Aminobutyric Acid Quantification
2.6. Determination of Glutamate Decarboxylase Activity
2.7. Determination of γ–Aminobutyric Transaminase Activity
2.8. Metabolomic Sample Extraction
2.9. HPLC–MS/MS Analysis
2.10. Data Processing and Annotation
3. Results and Discussion
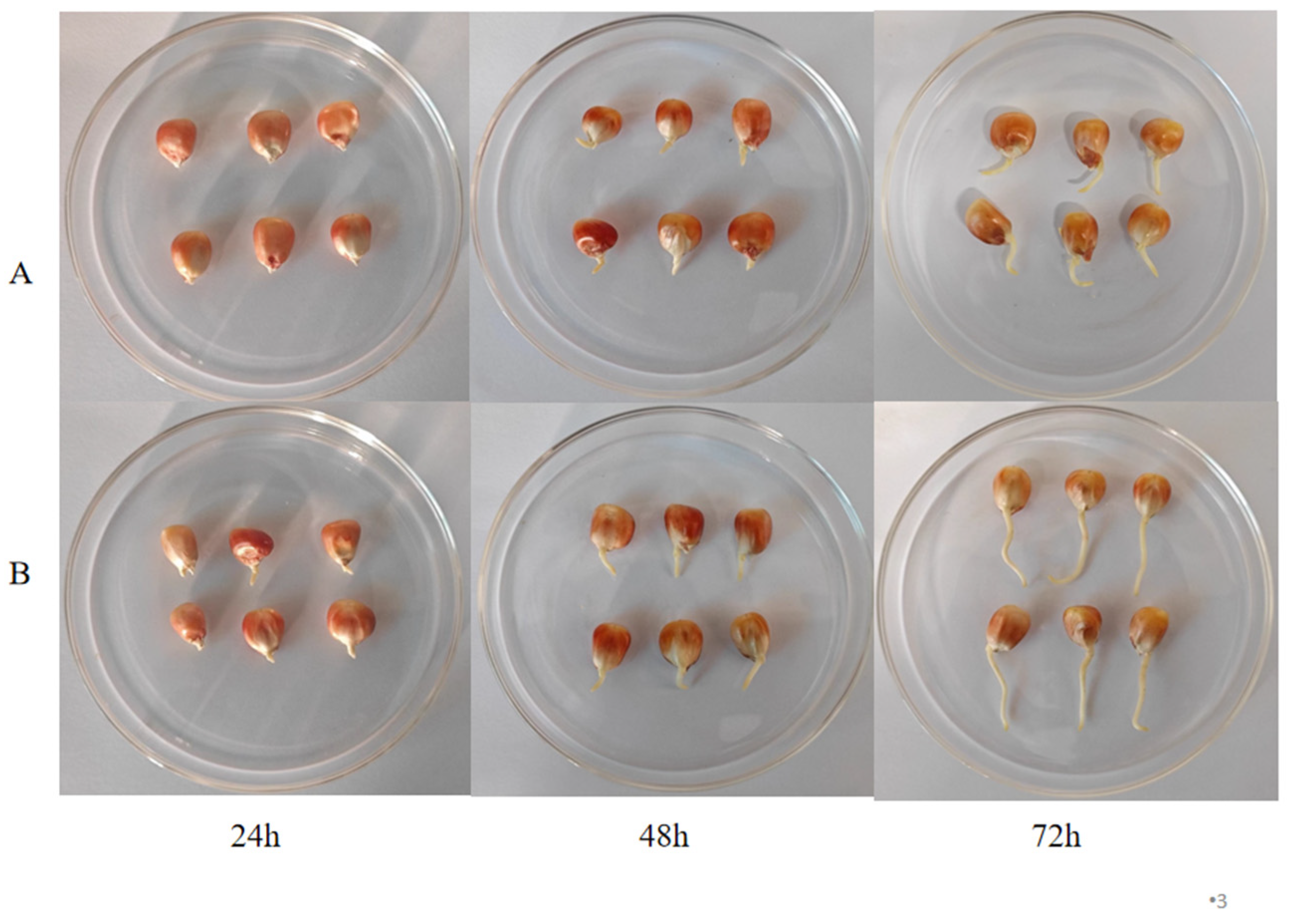
3.1. Effect of Pulsed Light Treatment on Morphological Characteristics and Basal Nutrients in Germinated Maize
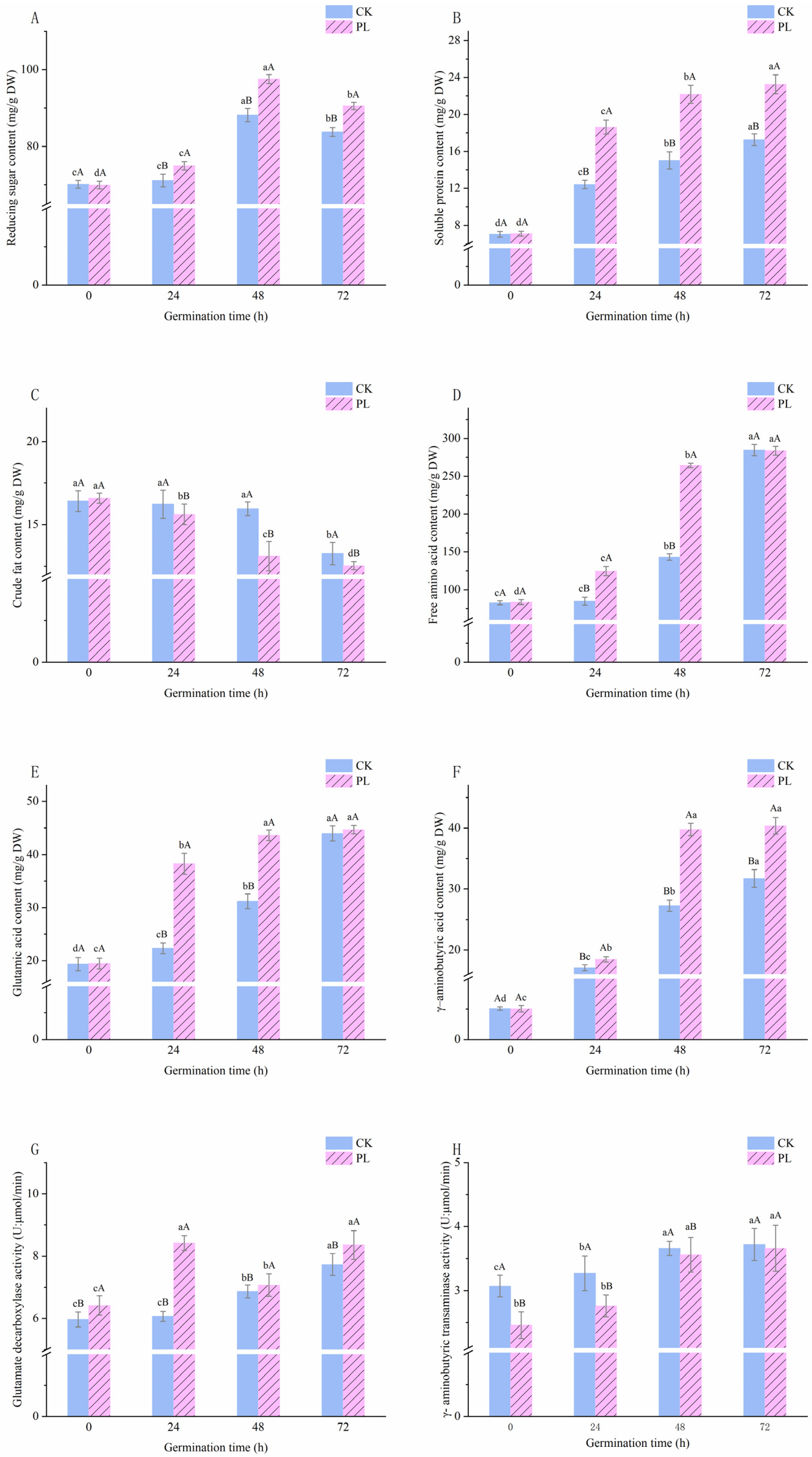
3.2. Effect of Pulsed Light Treatment on Free Amino Acids and Glutamic Acid Content in Germinated Maize
3.3. Effect of Pulsed Light Treatment on Glutamic Acid Decarboxylase Activity, γ–Aminobutyric Transaminase, and γ–Aminobutyric Acid Content in Germinated Maize
3.4. Metabolomics Analysis of Pulsed Light Treatment on γ–Aminobutyric Acid Enrichment in Germinated Maize
3.4.1. Liquid Chromatography Mass Spectrometry/Mass Spectrometry analysis
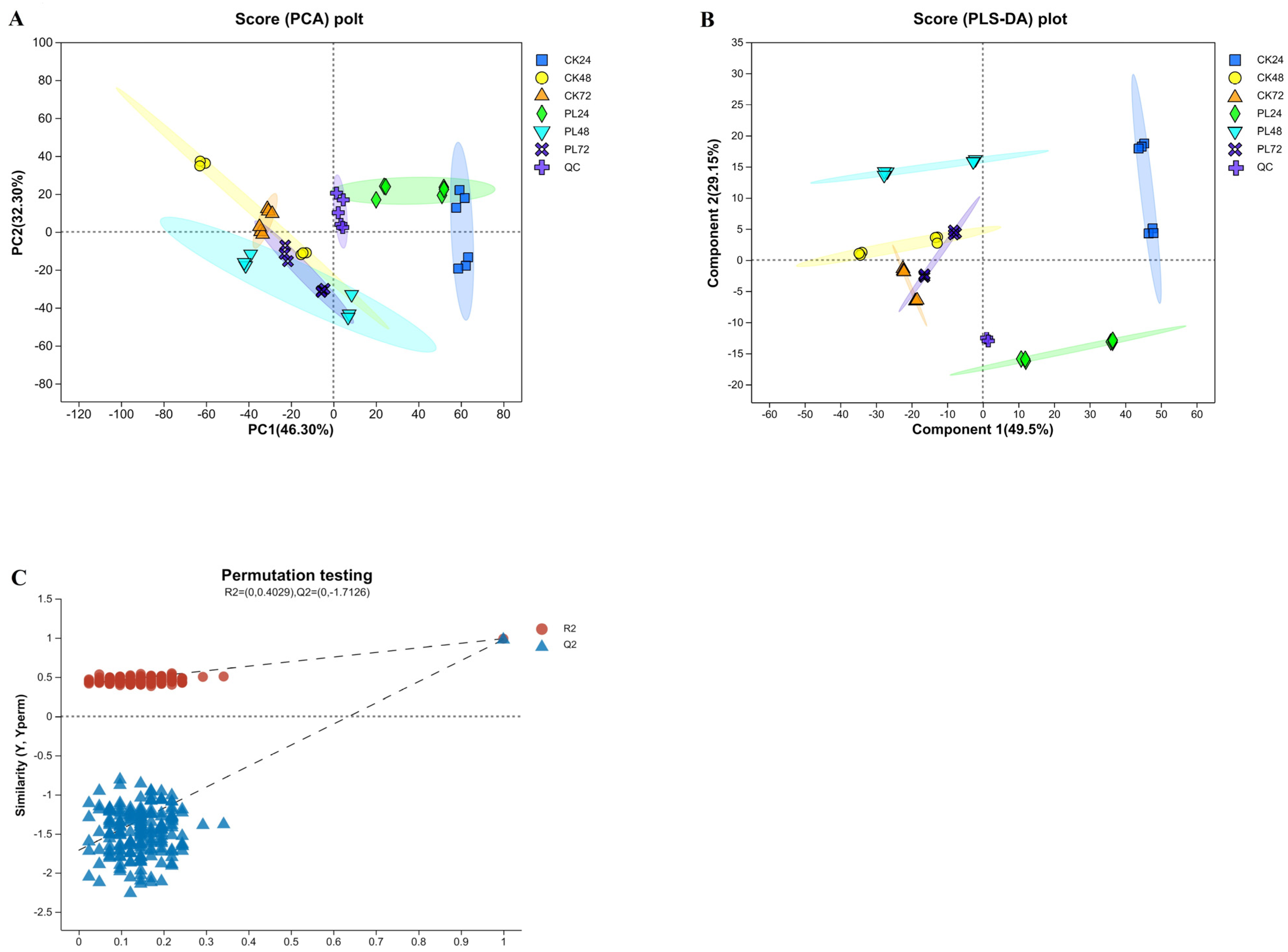
3.4.2. Principal Components Analysis and Partial Least Squares Discriminant Analysis of the Metabolites
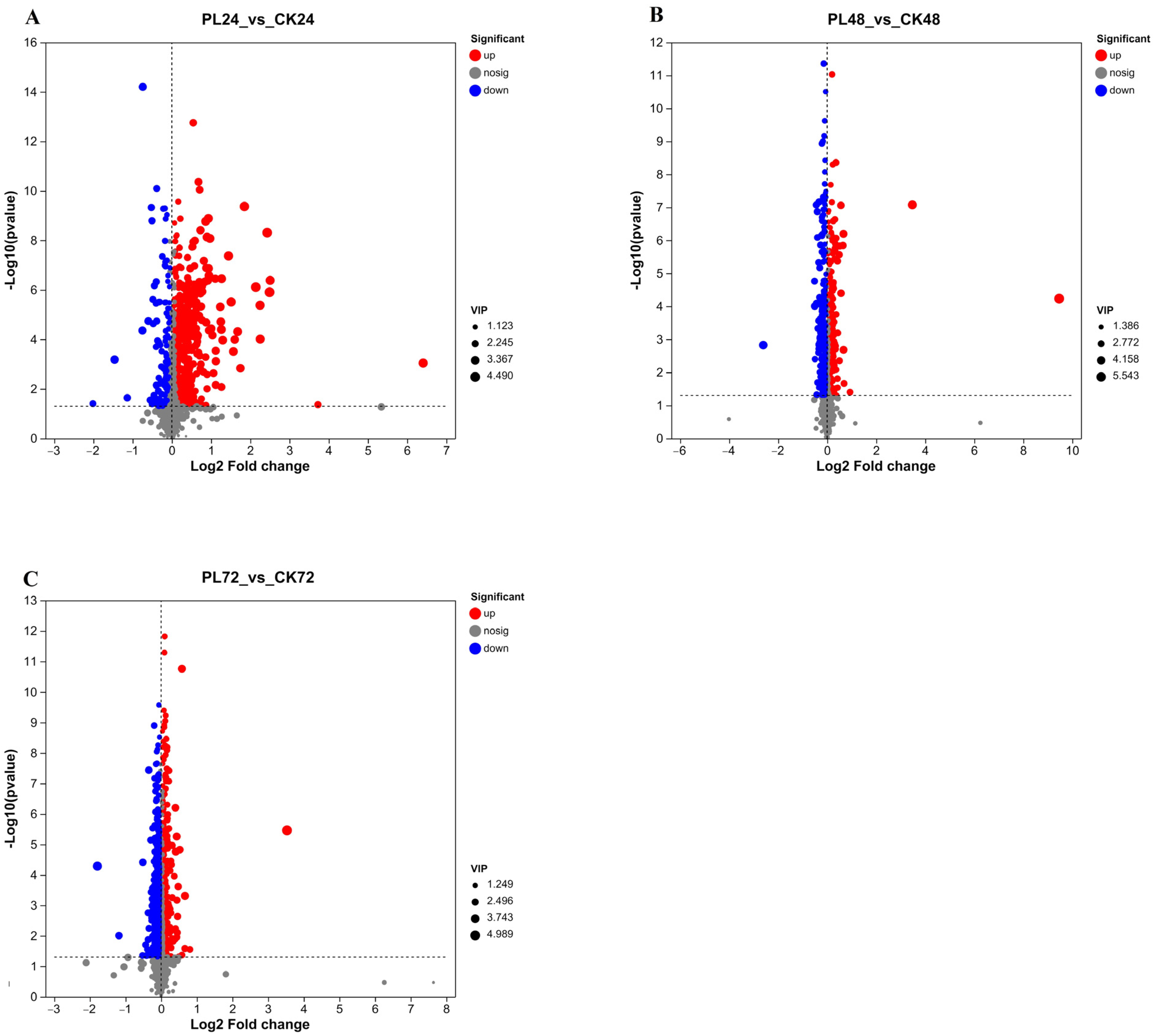
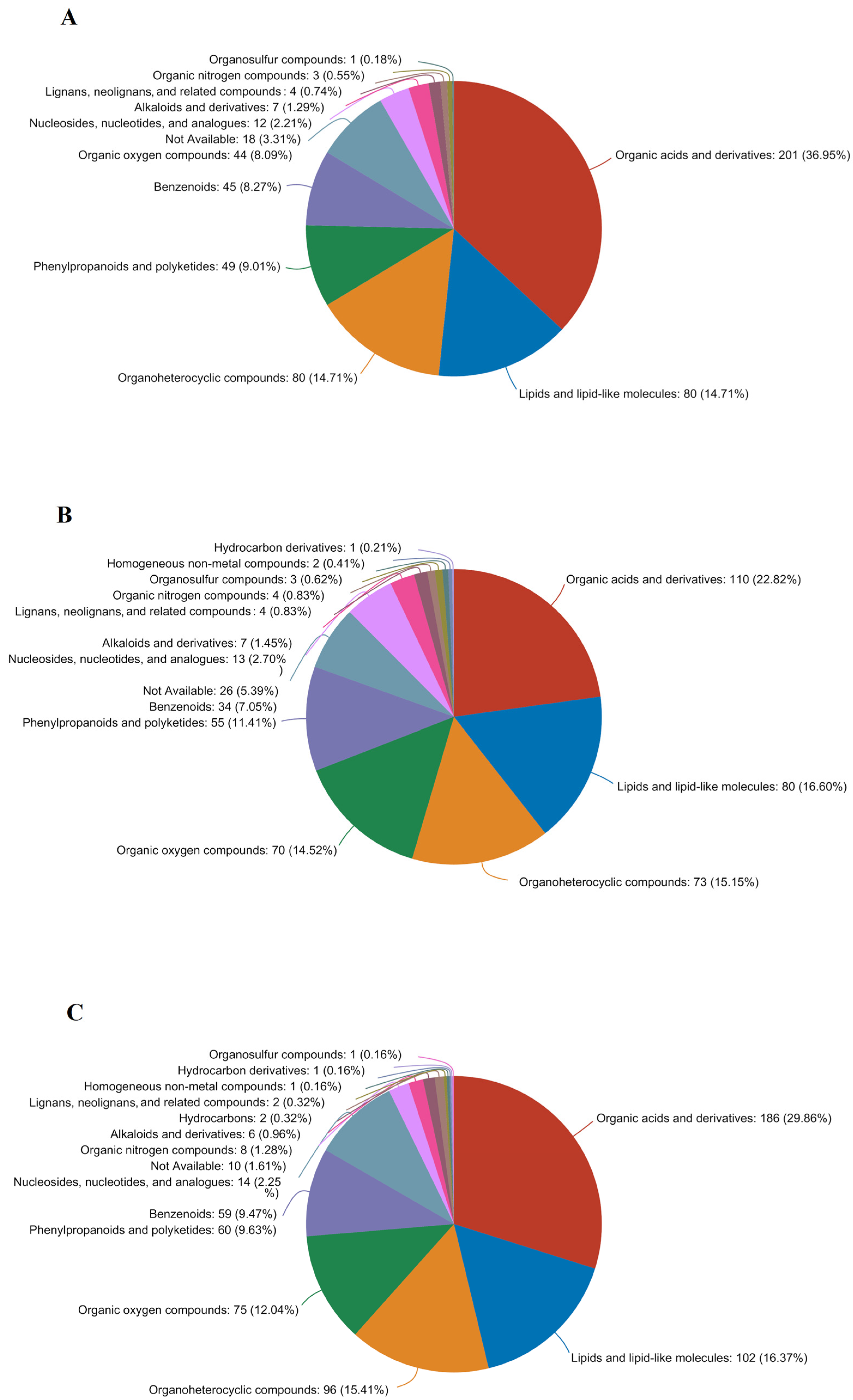
3.4.3. Analysis of Differential Metabolites in Germinated Maize
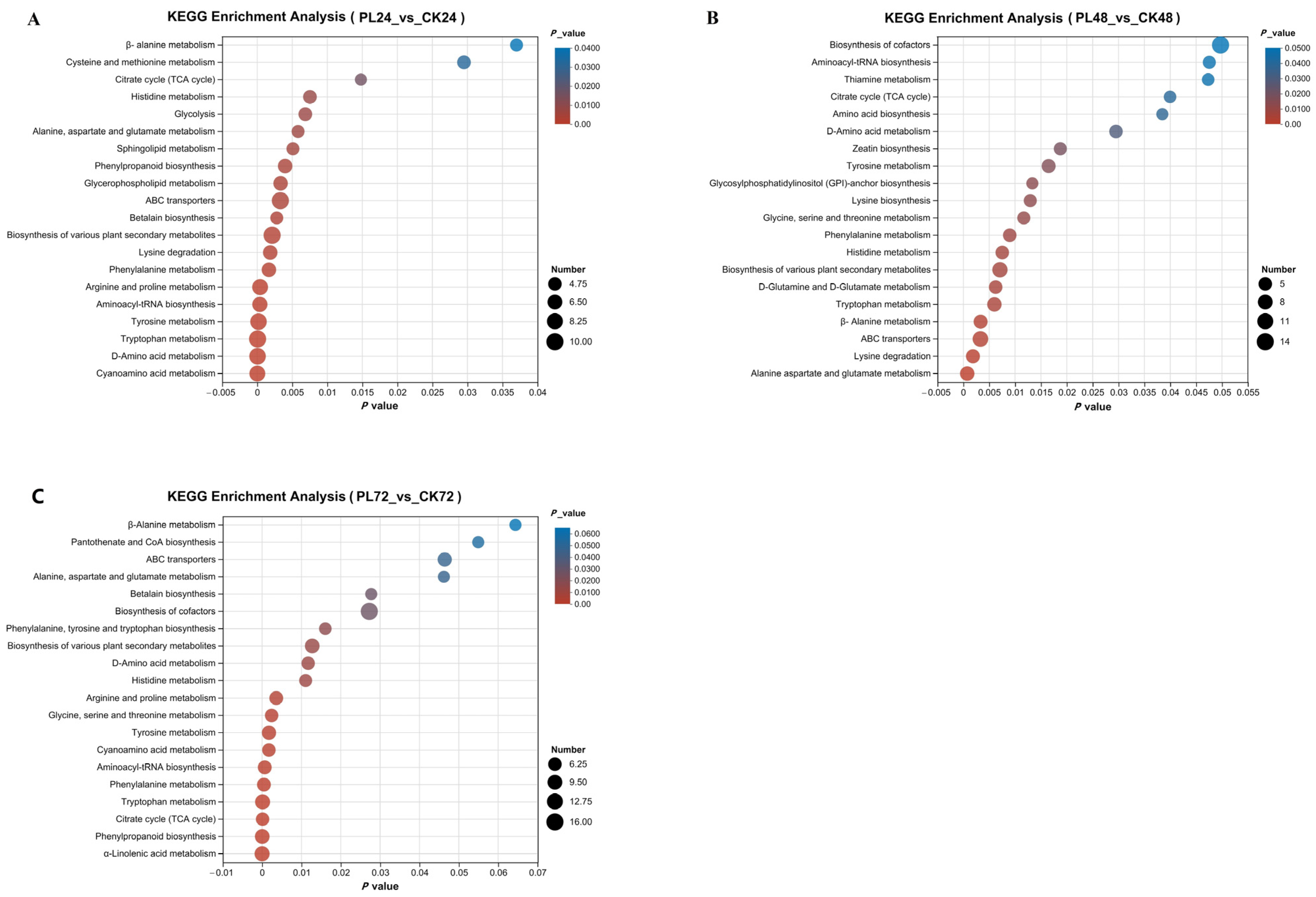
3.4.4. Kyoto Encyclopedia of Genes and Genomes Metabolic Pathway Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef]
- He, W.; Wang, Y.; Dai, Z.; Dai, Z.; Liu, C.; Xiao, Y.; Wei, Q.; Song, J.; Li, D. Effect of UV-B radiation and a supplement of CaCl2 on carotenoid biosynthesis in germinated corn kernels. Food Chem. 2019, 278, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, H.; Wang, S. Application of ultrasound, microwaves, and magnetic fields techniques in the germination of cereals. Food Sci. Technol. Res. 2019, 25, 489–497. [Google Scholar] [CrossRef]
- Nikmaram, N.; Dar, B.; Roohinejad, S.; Koubaa, M.; Barba, F.J.; Greiner, R.; Johnson, S.K. Recent advances in γ-aminobutyric acid (GABA) properties in pulses: An overview. J. Sci. Food Agr. 2017, 97, 2681–2689. [Google Scholar] [CrossRef]
- Suwanmanon, K.; Hsieh, P.-C.; Analysis, D. Effect of γ-aminobutyric acid and nattokinase-enriched fermented beans on the blood pressure of spontaneously hypertensive and normotensive Wistar-Kyoto rats. J. Food Drug. Anal. 2014, 22, 485–491. [Google Scholar] [CrossRef]
- Wu, C.; Huang, Y.; Lai, X.; Lai, R.; Zhao, W.; Zhang, M.; Zhao, W. Study on quality components and sleep-promoting effect of GABA Maoyecha tea. J. Funct. Foods 2014, 7, 180–190. [Google Scholar] [CrossRef]
- Han, J.; Zhao, X.; Zhao, X.; Wang, Q.; Li, P.; Gu, Q. Microbial-Derived γ-Aminobutyric Acid: Synthesis, Purification, Physiological Function, and Applications. J. Agric. Food Chem. 2023, 71, 14931–14946. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Tao, H.; Li, Y.; Pan, D.; Cao, J.; Liu, L.; Zhou, X.; Barba, F.J. Characterizing physicochemical, nutritional and quality attributes of wholegrain Oryza sativa L. subjected to high intensity ultrasound-stimulated pre–germination. Food Control 2020, 108, 106827. [Google Scholar] [CrossRef]
- Yin, Y.; Yang, R.; Gu, Z. Calcium regulating growth and GABA metabolism pathways in germinating soybean (Glycine max L.) under NaCl stress. Eur. Food Res. Technol. 2014, 239, 149–156. [Google Scholar] [CrossRef]
- Youn, Y.-S.; Park, J.-K.; Jang, H.-D.; Rhee, Y.-W. Sequential hydration with anaerobic and heat treatment increases GABA (γ-aminobutyric acid) content in wheat. Food Chem. 2011, 129, 1631–1635. [Google Scholar] [CrossRef]
- Heinrich, V.; Zunabovic, M.; Bergmair, J.; Kneifel, W.; Jäger, H. Post-packaging application of pulsed light for microbial decontamination of solid foods: A review. Innov. Food Sci. Emerg. 2015, 30, 145–156. [Google Scholar] [CrossRef]
- Chen, B.Y.; Lung, H.M.; Yang, B.B.; Wang, C.Y. Pulsed light sterilization of packaging materials. Food Packag. Shelf 2015, 5, 1–9. [Google Scholar] [CrossRef]
- Xia, Q.; Green, B.D.; Zhu, Z.; Li, Y.; Gharibzahedi, S.M.T.; Roohinejad, S.; Barba, F.J. Innovative processing techniques for altering the physicochemical properties of wholegrain brown rice (Oryza sativa L.)—Opportunities for enhancing food quality and health attributes. Crit. Rev. Food Sci. 2019, 59, 3349–3370. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Gui, Z.; Huang, Y.; Yang, H.; Luo, J.; Zeng, X. Integrated transcriptomics and metabolomics analyses provide insights into qingke in response to cold stress. J. Agr. Food Chem. 2023, 71, 18345–18358. [Google Scholar] [CrossRef]
- Wang, Z.; Ahmad, W.; Zhu, A.; Geng, W.; Kang, W.; Ouyang, Q.; Chen, Q. Identification of volatile compounds and metabolic pathway during ultrasound-assisted kombucha fermentation by HS–SPME–GC/MS combined with metabolomic analysis. Ultrason. Sonochem. 2023, 94, 106339. [Google Scholar] [CrossRef]
- Villate, A.; San Nicolas, M.; Gallastegi, M.; Aulas, P.; Olivares, M.; Usobiaga, A.; Etxebarria, N.; Aizpurua–Olaizola, O. Metabolomics as a prediction tool for plants performance under environmental stress. Plant Sci. 2021, 303, 110789. [Google Scholar] [CrossRef]
- Hiran, P.; Kerdchoechuen, O.; Laohakunjit, N. Combined effects of fermentation and germination on nutritional compositions, functional properties and volatiles of maize seeds. J. Cereal Sci. 2016, 71, 207–216. [Google Scholar] [CrossRef]
- Saqib, A.A.N.; Whitney, P.J. Differential behaviour of the dinitrosalicylic acid (DNS) reagent towards mono–and di–saccharide sugars. Biomass Bioenergy 2011, 35, 4748–4750. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 71, 248–254. [Google Scholar] [CrossRef]
- Cheung, P.C.; Leung, A.Y.; Ang, P.O. Comparison of supercritical carbon dioxide and Soxhlet extraction of lipids from a brown seaweed, Sargassum hemiphyllum (Turn.) C. Ag. J. Agr. Food Chem. 1998, 46, 4228–4232. [Google Scholar] [CrossRef]
- Chen, H.H.; Chang, H.C.; Chen, Y.K.; Hung, C.L.; Lin, S.Y.; Chen, Y.S. An improved process for high nutrition of germinated brown rice production: Low-pressure plasma. Food Chem. 2016, 191, 120–127. [Google Scholar] [CrossRef]
- Khwanchai, P.; Chinprahast, N.; Pichyangkura, R.; Chaiwanichsiri, S. Gamma-aminobutyric acid and glutamic acid contents, and the GAD activity in germinated brown rice (Oryza sativa L.): Effect of rice cultivars. Food Sci. Biotechnol. 2014, 23, 373–379. [Google Scholar] [CrossRef]
- Huang, X.-J.; Jian, S.-F.; Wan, S.; Miao, J.-H.; Zhong, C. Exogenous γ-aminobutyric acid (GABA) alleviates nitrogen deficiency by mediating nitrate uptake and assimilation in Andrographis paniculata seedlings. Plant Physiol. Bioch. 2023, 198, 107700. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Al-Dalali, S.; Zhou, H.; Xu, B. Influence of curing on the metabolite profile of water-boiled salted duck. Food Chem. 2022, 397, 133752. [Google Scholar] [CrossRef]
- Lee, S.-W.; Seo, J.M.; Lee, M.-K.; Chun, J.-H.; Antonisamy, P.; Arasu, M.V.; Suzuki, T.; Al-Dhabi, N.A.; Kim, S.-J. Influence of different LED lamps on the production of phenolic compounds in common and Tartary buckwheat sprouts. Ind. Crop. Prod. 2014, 54, 320–326. [Google Scholar] [CrossRef]
- Lu, T.-C.; Meng, L.-B.; Yang, C.-P.; Liu, G.-F.; Liu, G.-J.; Ma, W.; Wang, B.-C. A shotgun phosphoproteomics analysis of embryos in germinated maize seeds. Planta 2008, 228, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Gołda, A.; Szyniarowski, P.; Ostrowska, K.; Kozik, A.; Rąpała-Kozik, M. Thiamine binding and metabolism in germinating seeds of selected cereals and legumes. Plant. Physiol. Bioch. 2004, 42, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Mackerness, S.A.H. Plant responses to ultraviolet-B (UV-B: 280–320 nm) stress: What are the key regulators? Plant growth regul. 2000, 32, 27–39. [Google Scholar] [CrossRef]
- Xia, Q.; Wang, L.; Li, Y. Exploring high hydrostatic pressure-mediated germination to enhance functionality and quality attributes of wholegrain brown rice. Food Chem. 2018, 249, 104–110. [Google Scholar] [CrossRef]
- Ding, J.; Ulanov, A.V.; Dong, M.; Yang, T. Enhancement of gama–aminobutyric acid (GABA) and other health-related metabolites in germinated red rice (Oryza sativa L.) by ultrasonication. Ultrason. Sonochem. 2018, 40, 791–797. [Google Scholar] [CrossRef]
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (gaba) and plant responses to stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Scott-Taggart, C.P.; Van Cauwenberghe, O.R.; Mclean, M.D.; Shelp, B.J. Regulation of γ-aminobutyric acid synthesis in situ by glutamate availability. Physiol. Plantarum 1999, 106, 363–369. [Google Scholar] [CrossRef]
- Dennison, K.L.; Spalding, E.P. Glutamate-gated calcium fluxes in Arabidopsis. Plant Physiol. 2000, 124, 1511–1514. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-H.; Dong, C.-D.; Kumar Patel, A.; Rani Singhania, R.; Yang, M.-J.; Guo, H.-R.; Kuo, J.-M. Characterization of waste cell biomass derived glutamate decarboxylase for in vitro gamma-aminobutyric acid production and value-addition. Bioresour. Technol. 2021, 337, 125423. [Google Scholar] [CrossRef] [PubMed]
- Mae, N.; Makino, Y.; Oshita, S.; Kawagoe, Y.; Tanaka, A.; Aoki, K.; Kurabayashi, A.; Akihiro, T.; Akama, K.; Koike, S.; et al. Accumulation mechanism of γ-aminobutyric acid in tomatoes (Solanum lycopersicum L.) under low O2 with and without CO2. J. Agr. Food Chem. 2012, 60, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Zhen, S.; Zhu, G.; Bian, Y.; Yan, Y. Comparative metabolome analysis of wheat embryo and endosperm reveals the dynamic changes of metabolites during seed germination. Plant Physiol. Bioch. 2017, 115, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Cai, Z.; Deng, Y.; Pan, D.; Zhou, C.; Cao, J.; Chen, X.; Xia, Q. An untargeted metabolomic insight into the high-pressure stress effect on the germination of wholegrain Oryza sativa L. Food Res. Int. 2021, 140, 109984. [Google Scholar] [CrossRef]
- Girija, A.; Yadav, R.; Corke, F.; Doonan, J.; Mur, L.A. Untargeted metabolomic profiling reveals variation in metabolites associated with nutritional values in tef accessions. Plant Food Hum. Nutr. 2021, 76, 536–539. [Google Scholar] [CrossRef]
- Xiong, Q.; Zhang, J.; Sun, C.; Wang, R.; Wei, H.; He, H.; Zhou, D.; Zhang, H.; Zhu, J. Metabolomics revealed metabolite biomarkers of antioxidant properties and flavonoid metabolite accumulation in purple rice after grain filling. Food Chem. X 2023, 18, 100720. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.G.; Hu, Q.P.; Duan, J.L.; Tian, C.R. Dynamic changes in γ-aminobutyric acid and glutamate decarboxylase activity in oats (Avena nuda L.) during steeping and germination. J. Agr. Food Chem. 2010, 58, 9759–9763. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Devireddy, A.R.; Azad, R.K.; Shulaev, V.; Mittler, R. Rapid accumulation of glutathione during light stress in Arabidopsis. Plant Cell Physiol. 2018, 59, 1817–1826. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chen, H.; Gu, Z. Factors influencing diamine oxidase activity and γ-aminobutyric acid content of fava bean (Vicia faba L.) during germination. J. Agr. Food Chem. 2011, 59, 11616–11620. [Google Scholar] [CrossRef]
- Ma, M.; Zhang, H.; Xie, Y.; Yang, M.; Tang, J.; Wang, P.; Yang, R.; Gu, Z. Response of nutritional and functional composition, anti-nutritional factors and antioxidant activity in germinated soybean under UV-B radiation. LWT Food Sci. Technol. 2020, 118, 108709. [Google Scholar] [CrossRef]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Guan, D.; Fan, B.; Sun, Y.; Wang, F. Untargeted mass spectrometry-based metabolomics approach unveils molecular changes in heat-damaged and normal soybean. LWT Food Sci. Technol. 2022, 171, 114136. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, X.; Li, A.; Wang, R.; Ni, X.; Hu, J.; Wei, H.; Zhang, H.; Xiong, Q. The main nutritional components in colored rice grains. LWT Food Sci. Technol. 2024, 191, 115663. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Liu, X.; Xu, L.; Xie, M.; Yu, M. Non-Targeted Metabolomics Analysis of γ–Aminobutyric Acid Enrichment in Germinated Maize Induced by Pulsed Light. Foods 2024, 13, 2675. https://doi.org/10.3390/foods13172675
Zhang L, Liu X, Xu L, Xie M, Yu M. Non-Targeted Metabolomics Analysis of γ–Aminobutyric Acid Enrichment in Germinated Maize Induced by Pulsed Light. Foods. 2024; 13(17):2675. https://doi.org/10.3390/foods13172675
Chicago/Turabian StyleZhang, Liangchen, Xiaojing Liu, Liwei Xu, Mengxi Xie, and Miao Yu. 2024. "Non-Targeted Metabolomics Analysis of γ–Aminobutyric Acid Enrichment in Germinated Maize Induced by Pulsed Light" Foods 13, no. 17: 2675. https://doi.org/10.3390/foods13172675
APA StyleZhang, L., Liu, X., Xu, L., Xie, M., & Yu, M. (2024). Non-Targeted Metabolomics Analysis of γ–Aminobutyric Acid Enrichment in Germinated Maize Induced by Pulsed Light. Foods, 13(17), 2675. https://doi.org/10.3390/foods13172675