
Non-Covalent Interaction of Folic Acid and 5-Methyltetrahydrofolate with Caseinates Improves the Folates Stability Studied by Multi-Spectroscopic Analysis and Molecular Docking
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Fluorescence Spectroscopy
2.4. Surface Hydrophobicity
2.5. Fourier Transform Infrared (FTIR) Spectroscopy
2.6. Molecular Docking
2.7. Storage Stability
2.7.1. Sample Storage
2.7.2. Physical Stability
2.7.3. Chemical Stability
2.8. Differencial Scanning Calorimetry (DSC)
2.9. Statistical Analysis
3. Results and Discussion
3.1. Quenching Mechanism of NaCas by Folate
3.2. Thermodynamic Parameters of the NaCas-Folate System
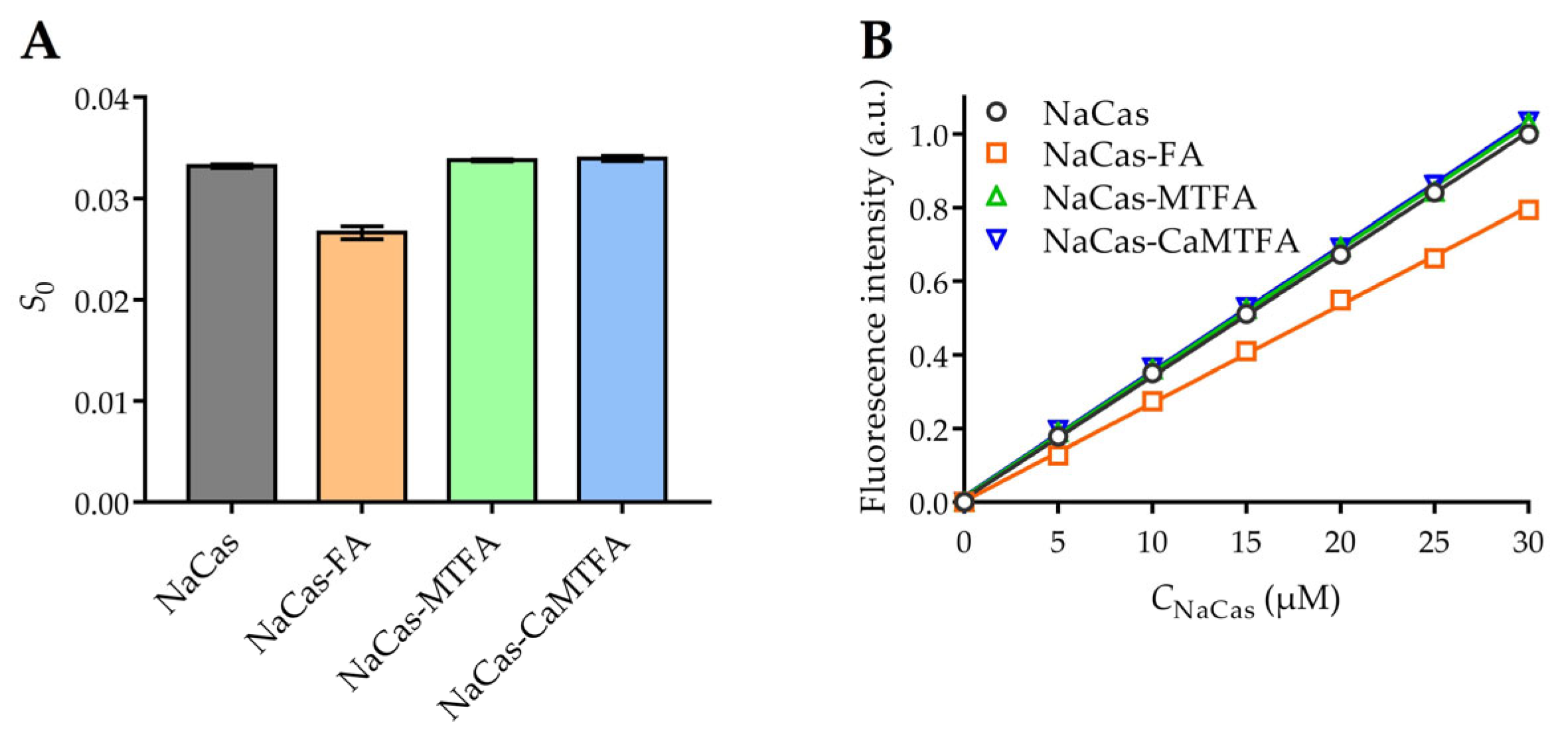
3.3. Surface Hydrophobicity
3.4. Fourier Transform Infrared (FTIR) Spectra
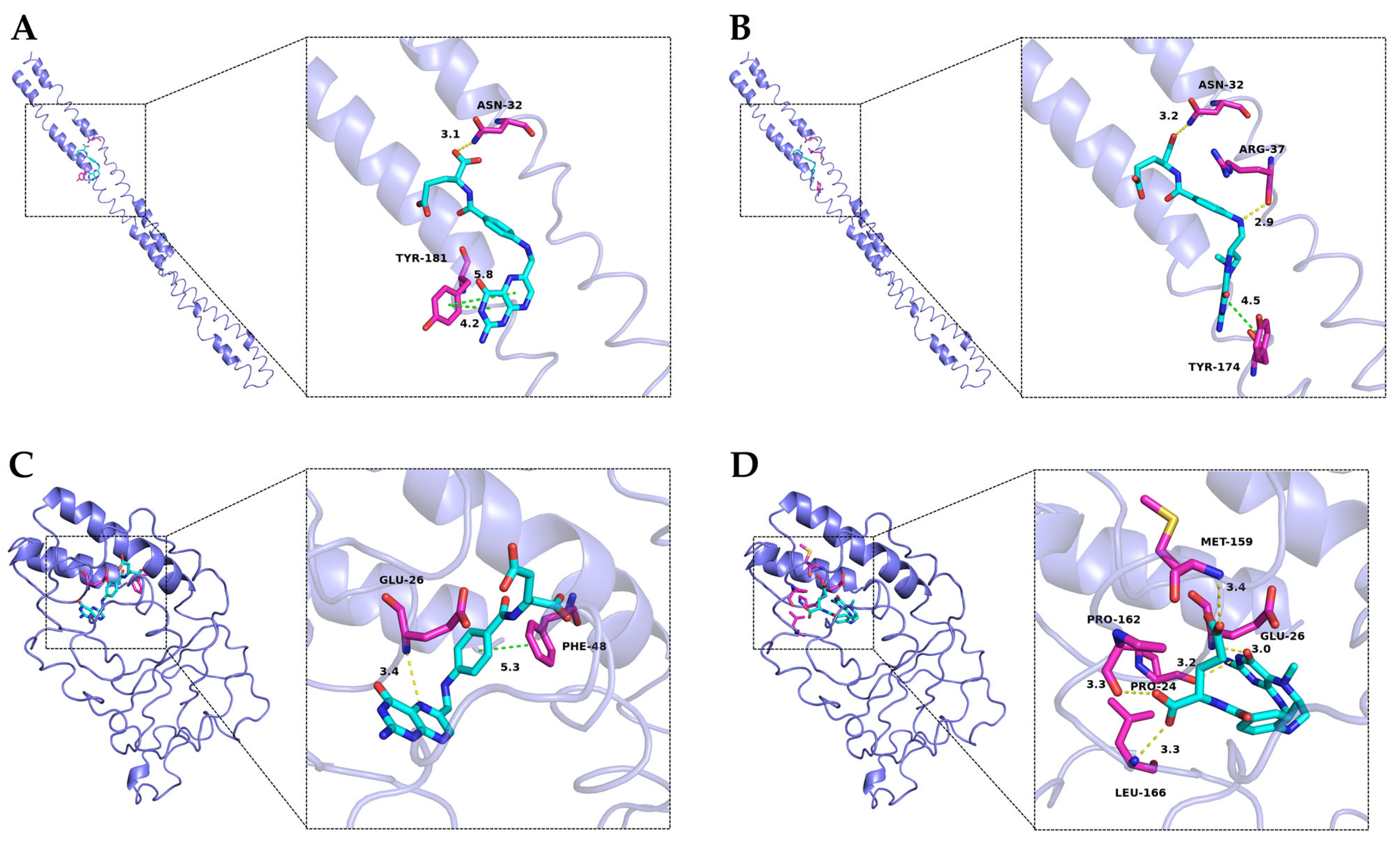
3.5. Molecular Docking Study
3.6. Physical Stability of Complexes
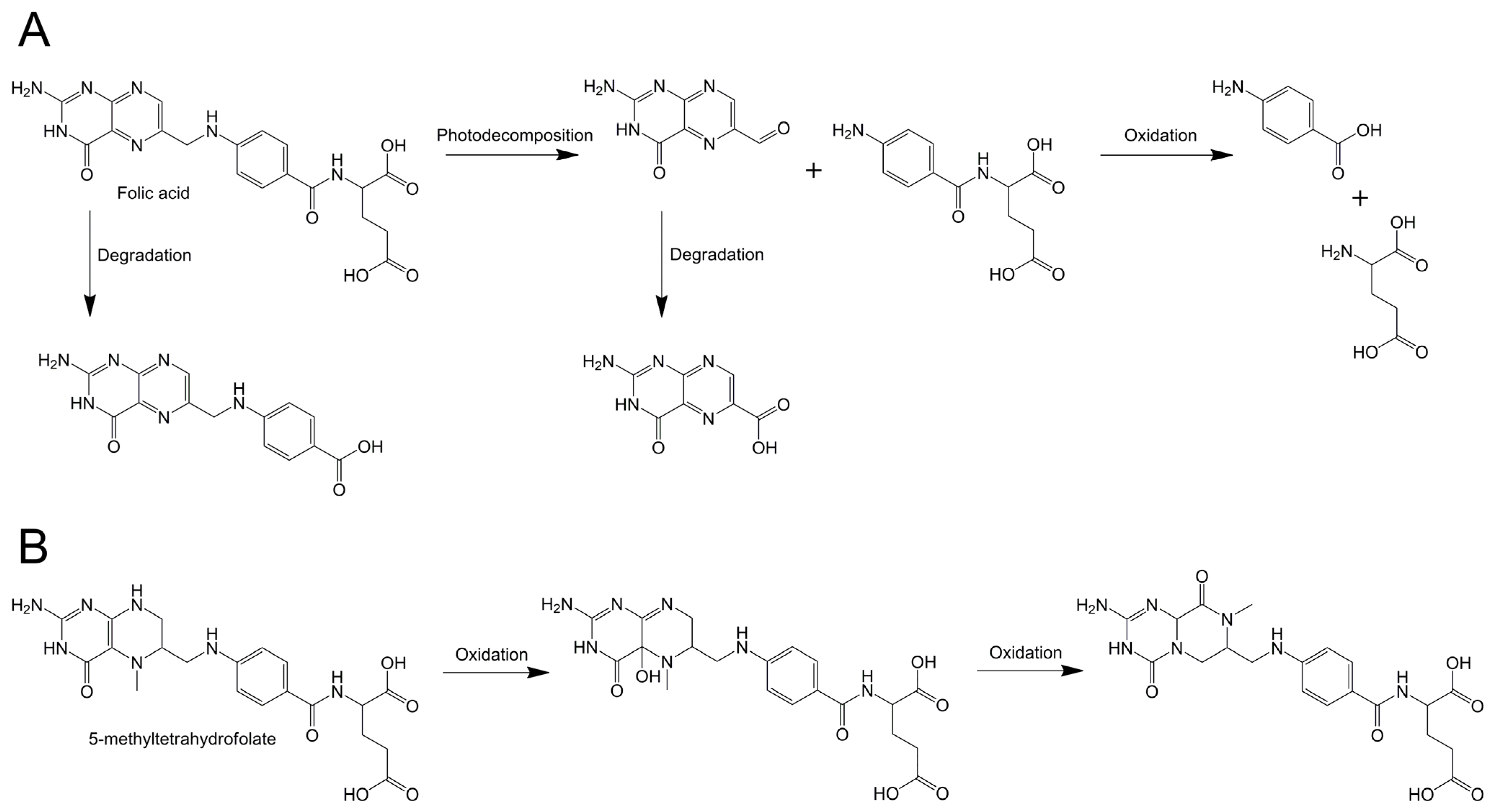
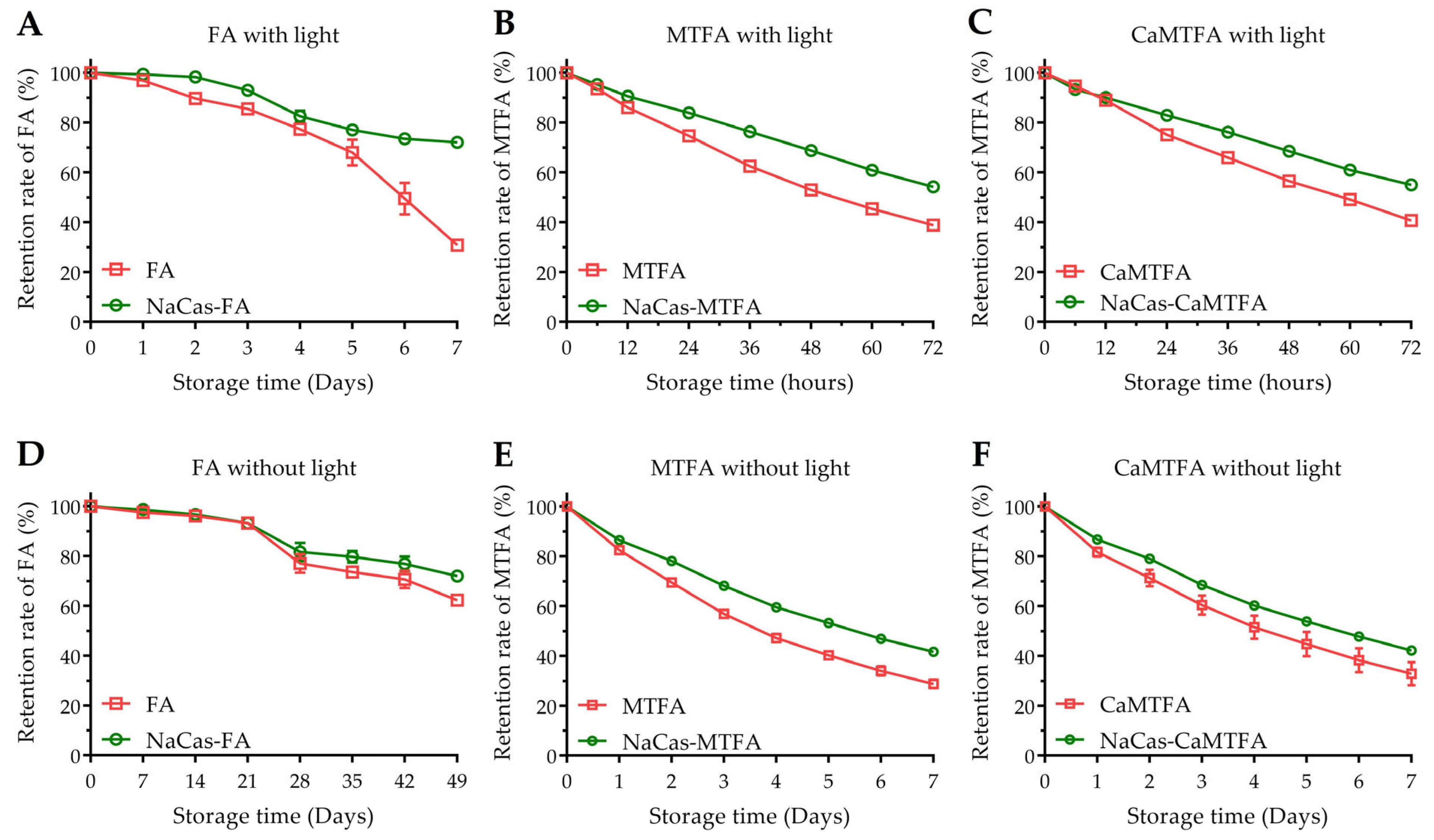
3.7. Chemical Stability of Folates
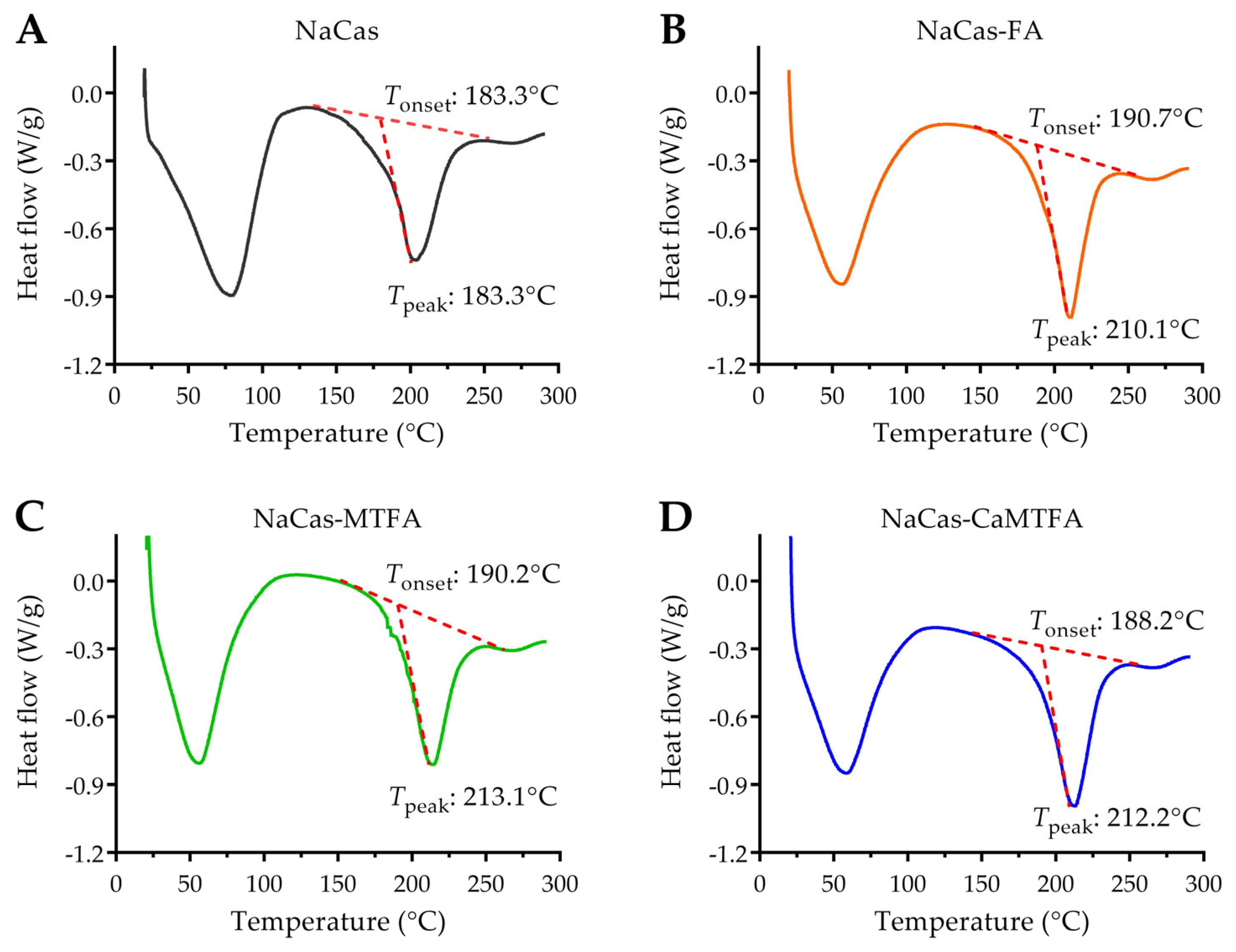
3.8. Thermal Stability of Protein by DSC
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wusigale; Liang, L. Folates: Stability and interaction with biological molecules. J. Agric. Food Res. 2020, 2, 100039. [Google Scholar] [CrossRef]
- Saini, R.K.; Nile, S.H.; Keum, Y.S. Folates: Chemistry, analysis, occurrence, biofortification and bioavailability. Food Res. Int. 2016, 8, 1–13. [Google Scholar] [CrossRef]
- Liu, M.; Chen, Q.; Sun, Y.; Zeng, L.; Wu, H.; Gu, Q.; Li, P. Probiotic potential of a folate-producing strain Latilactobacillus sakei LZ217 and its modulation effects on human gut microbiota. Foods 2020, 11, 234. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Edelmann, M.; Piironen, V.; Kariluoto, S. 5-methyltetrahydrofolate is a crucial factor in determining the bioaccessibility of folate in bread. J. Agric. Food Chem. 2022, 70, 13379–13390. [Google Scholar] [CrossRef] [PubMed]
- Lian, Z.; Wu, Z.; Gu, R.; Wang, Y.; Wu, C.; Cheng, Z.; He, M.; Wang, Y.; Cheng, Y.; Gu, H.F. Evaluation of cardiovascular toxicity of folic acid and 6S-5-methyltetrahy-drofolate-calcium in early embryonic development. Cells 2022, 11, 3946. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, K.; Bouhallab, S.; Croguennec, T.; Renard, D.; Lechevalier, V. Application of high-pressure and ultrasound technologies for legume proteins as wall material in microencapsulation: New insights and advances. Trends Food Sci. Technol. 2022, 127, 49–62. [Google Scholar] [CrossRef]
- Fathi, M.; Donsi, F.; McClements, D.J. Protein-based delivery systems for the nanoencapsulation of food ingredients. Compr. Rev. Food Sci. Food Saf. 2018, 17, 920–936. [Google Scholar] [CrossRef]
- Tang, C.H. Assembly of food proteins for nano-encapsulation and delivery of nutraceuticals (a mini-review). Food Hydrocoll. 2021, 117, 106710. [Google Scholar] [CrossRef]
- Wusigale; Liang, L.; Luo, Y. Casein and pectin: Structures, interactions, and applications. Trends Food Sci. Technol. 2020, 97, 391–403. [Google Scholar] [CrossRef]
- Rehan, F.; Ahemad, N.; Gupta, M. Casein nanomicelle as an emerging biomaterial—A comprehensive review. Colloids Surf. B 2019, 179, 280–292. [Google Scholar] [CrossRef]
- Casanova, F.; Nascimento, L.G.L.; Silva, N.F.N.; de Carvalho, A.F.; Gaucheron, F. Interactions between caseins and food-derived bioactive molecules: A review. Food Chem. 2021, 359, 129820. [Google Scholar] [CrossRef] [PubMed]
- Rashidinejad, A.; Jameson, G.B.; Singh, H. The Effect of pH and Sodium Caseinate on the Aqueous Solubility, Stability, and Crystallinity of Rutin towards Concentrated Colloidally Stable Particles for the Incorporation into Functional Foods. Molecules 2022, 27, 534. [Google Scholar] [CrossRef]
- Ghayour, N.; Hosseini, S.M.H.; Eskandari, M.H.; Esteghlal, S.; Nekoei, A.R.; Gahruie, H.H.; Tatar, M.; Naghibalhossaini, F. Nanoencapsulation of quercetin and curcumin in casein-based delivery systems. Food Hydrocoll. 2019, 87, 394–403. [Google Scholar] [CrossRef]
- Penalva, R.; Esparza, I.; Agüeros, M.; Gonzalez-Navarro, C.J.; Gonzalez-Ferrero, C.; Irache, J.M. Casein nanoparticles as carriers for the oral delivery of folic acid. Food Hydrocoll. 2015, 44, 399–406. [Google Scholar] [CrossRef]
- Malekhosseini, P.; Alami, M.; Khomeiri, M.; Esteghlal, S.; Nekoei, A.; Hosseini, S.M.H. Development of casein-based nanoencapsulation systems for delivery of epigallocatechin gallate and folic acid. Food Sci. Nutr. 2019, 7, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, P.; Tajmir-Riahi, H.A. Locating the binding sites of folic acid with milk α- and β-caseins. J. Phys. Chem. B 2012, 116, 513–519. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Liu, X.; Li, Y.; Yin, X.; Subirade, M.; Zhou, P.; Liang, L. The folic acid/β-casein complex: Characteristics and physicochemical implications. Food Res. Int. 2014, 57, 162–167. [Google Scholar] [CrossRef]
- Li, M.; O’Mahony, J.A.; Kelly, A.L.; Brodkorb, A. The influence of temperature- and divalent-cation-mediated aggregation of β-casein on the physical and microstructural properties of β-casein-stabilised emulsions. Colloids Surf. B 2020, 187, 110620. [Google Scholar] [CrossRef] [PubMed]
- Lelis, C.A.; Galvan, D.; Conte-Junior, C.A. Nanocarriers for β-carotene based on milk protein. Food Bioprocess Technol. 2023, 16, 43–67. [Google Scholar] [CrossRef]
- Dickinson, E.; Casanova, H. A thermoreversible emulsion gel based on sodium caseinate. Food Hydrocoll. 1999, 13, 285–289. [Google Scholar] [CrossRef]
- Mantovani, R.A.; Hamon, P.; Rousseau, F.; Tavares, G.M.; Mercadante, A.Z.; Croguennec, T.; Bouhallab, S. Unraveling the molecular mechanisms underlying interactions between caseins and lutein. Food Res. Int. 2020, 138, 109781. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yang, X.; Chen, F.; Zeng, G.; Zhou, L.; Li, X.; Tuo, X. Spectroscopic and in silico insight into the interaction between dicofol and human serum albumin. Spectrochim. Acta A 2022, 264, 120277. [Google Scholar] [CrossRef]
- Jia, J.; Gao, X.; Hao, M.; Tang, L. Comparison of binding interaction between β-lactoglobulin and three common polyphenols using multi-spectroscopy and modeling methods. Food Chem. 2017, 228, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, N.; He, Q.; Sun, Q.; Zeng, W.C. Effects of casein on the stability, antioxidant activity, and bioavailability of lotus anthocyanins. J. Food Biochem. 2022, 46, e14288. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Liu, Y.; Li, J.; Sun, X.; Gu, J.; He, Y.; Ci, H.; Wen, L.; Yu, H.; Xu, X. Effect of soybean protein isolate-7s on delphinidin-3-o-glucoside from purple corn stability and their interactional characterization. Foods 2022, 11, 895. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.T.; Callis, P.R. Mechanisms of tryptophan fluorescence shifts in proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef]
- Hu, Y.J.; Liu, Y.; Xiao, X.H. Investigation of the interaction between berberine and human serum albumin. Biomacromolecules 2009, 10, 517–521. [Google Scholar] [CrossRef]
- Andersen, C.M.; Mortensen, G. Fluorescence spectroscopy: A rapid tool for analyzing dairy products. J. Agric. Food Chem. 2008, 56, 720–729. [Google Scholar] [CrossRef]
- Zhang, Q.A.; Fu, X.Z.; García Martín, J.F. Effect of ultrasound on the interaction between (−)-epicatechin gallate and bovine serum albumin in a model wine. Ultrason. Sonochem. 2017, 37, 405–413. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Zhang, L. Study on the mechanism of non-covalent interaction between rose anthocyanin extracts and whey protein isolate under different pH conditions. Food Chem. 2022, 384, 132492. [Google Scholar] [CrossRef]
- Tang, L.; Li, S.; Bi, H.; Gao, X. Interaction of cyanidin-3-O-glucoside with three proteins. Food Chem. 2016, 196, 550–559. [Google Scholar] [CrossRef]
- Chilom, C.G.; David, M.; Florescu, M. Monitoring biomolecular interaction between folic acid and bovine serum albumin. Spectrochim. Acta A 2020, 230, 118074. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Xu, M.; Zeng, M.; Qin, F.; Chen, J. Interactions of milk α-and β-casein with malvidin-3-O-glucoside and their effects on the stability of grape skin anthocyanin extracts. Food Chem. 2016, 199, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, M.; Yan, H.; Cai, C.; Guo, Q.; Pei, W.; Zhang, R.; Wang, Z.; Han, J. Mutual influence of piceatannol and bisphenol F on their interaction with pepsin: Insights from spectroscopic, isothermal titration calorimetry and molecular modeling studies. Spectrochim. Acta A 2019, 206, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, P.; Hasni, I.; Tajmir-Riahi, H.A. Folic acid complexes with human and bovine serum albumins. Food Chem. 2011, 129, 1148–1155. [Google Scholar] [CrossRef]
- Li, T.; Hu, P.; Dai, T.; Li, P.; Ye, X.; Chen, J.; Liu, C. Comparing the binding interaction between β-lactoglobulin and flavonoids with different structure by multi-spectroscopy analysis and molecular docking. Spectrochim. Acta A 2018, 201, 197–206. [Google Scholar] [CrossRef]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef]
- Ren, C.; Xiong, W.; Li, J.; Li, B. Comparison of binding interactions of cyanidin-3-O-glucoside to β-conglycinin and glycinin using multi-spectroscopic and thermodynamic methods. Food Hydrocoll. 2019, 92, 155–162. [Google Scholar] [CrossRef]
- Holt, C. A quantitative calcium phosphate nanocluster model of the casein micelle: The average size, size distribution and surface properties. Eur. Biophys. J. 2021, 50, 847–866. [Google Scholar] [CrossRef]
- Holt, C.; Carver, J. Quantitative multivalent binding model of the structure, size distribution and composition of the casein micelles of cow milk. Int. Dairy J. 2022, 126, 105292. [Google Scholar] [CrossRef]
- Wang, T.; Yadong Li, Y.; De Witte, F.; Rebry, F.; Li, H.; Vermeir, P.; Dewettinck, K.; Van der Meeren, P. Influence of calcium concentration on the re-assembly of sodium caseinate into casein micelles and on their renneting behavior. Food Res. Int. 2024, 180, 113991. [Google Scholar] [CrossRef]
- Zhan, F.; Li, J.; Wang, Y.; Shi, M.; Li, B.; Sheng, F. Bulk, foam, and interfacial properties of tannic acid/sodium caseinate nanocomplexes. J. Agric. Food Chem. 2018, 66, 6832–6839. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Yang, W.; Fan, R.; Yuan, F.; Gao, Y. Evaluation of structural and functional properties of protein–EGCG complexes and their ability of stabilizing a model β-carotene emulsion. Food Hydrocoll. 2015, 45, 337–350. [Google Scholar] [CrossRef]
- Qie, X.; Chen, W.; Wu, R.; Wang, Z.; Zeng, M.; Chen, J.; Goff, H.D.; He, Z. The effects of β-lactoglobulin on cyanidin-3-O-glucoside antioxidant activity and bioaccessibility after heat treatment. Food Res. Int. 2022, 157, 111494. [Google Scholar] [CrossRef]
- Dai, S.; Lian, Z.; Qi, W.; Chen, Y.; Tong, X.; Tian, T.; Lyu, B.; Wang, M.; Wang, H.; Jiang, L. Non-covalent interaction of soy protein isolate and catechin: Mechanism and effects on protein conformation. Food Chem. 2022, 384, 132507. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Ma, T.; Shi, L.; Wang, Q.; Zhang, L.; Zhang, F.; Wang, Z.; Zhou, Z. The formation of phycocyanin-EGCG complex for improving the color protection stability exposing to light. Food Chem. 2022, 370, 130985. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Q.; Lei, L.; Li, F.; Zhao, J.; Zhang, Y.; Li, L.; Wang, Q.; Ming, J. Molecular interaction of soybean glycinin and β-conglycinin with (−)-epigallocatechin gallate induced by pH changes. Food Hydrocoll. 2020, 108, 106010. [Google Scholar] [CrossRef]
- Abdollahi, K.; Condict, L.; Hung, A.; Kasapis, S. Binding parameters and molecular dynamics of β-lactoglobulin-vanillic acid complexation as a function of pH—Part A: Acidic pH. Food Chem. 2021, 360, 130059. [Google Scholar] [CrossRef]
- Nascimento, L.G.L.; Casanova, F.; Silva, N.F.N.; de Carvalho Teixeira, A.V.N.; de Carvalho, A.F. Casein-based hydrogels: A mini-review. Food Chem. 2020, 314, 126063. [Google Scholar] [CrossRef]
- Yi, J.; Fan, Y.; Yokoyama, W.; Zhang, Y.; Zhao, L. Characterization of milk proteins–lutein complexes and the impact on lutein chemical stability. Food Chem. 2016, 200, 91–97. [Google Scholar] [CrossRef]
- Zhao, R.; Qin, X.; Zhong, J. Interaction between curcumin and β-casein: Multi-spectroscopic and molecular dynamics simulation methods. Molecules 2021, 26, 5092. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Kang, S.; Zhang, J.; Jiang, L.; Liu, Y.; Yang, M.; Cao, X.; Zheng, Y.; Shao, J.; Yue, X. The non-covalent interactions between whey protein and various food functional ingredients. Food Chem. 2022, 394, 133455. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Tan, W.; Liu, W.; Wei, C.; Chen, J.; Zhao, Z.; Tian, J. Inhibited the walnut oil oxidation through the microcapsules that consisted of (−)-Epigallocatechin gallate and sodium caseinate. Food Biosci. 2024, 61, 104601. [Google Scholar] [CrossRef]
- Wang, T.; Li, H.; Denon, Q.; To, C.M.; Van der Meeren, P. Investigating the effect of diluents and fat globules on the size measurement of casein micelles by dynamic light scattering. Int. Dairy J. 2023, 142, 105640. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, R. pH-dependent structures and properties of casein micelles. Biophys. Chem. 2008, 136, 67–73. [Google Scholar] [CrossRef]
- Liu, Y.; Tomiuk, S.; Rozoy, E.; Simard, S.; Bazinet, L.; Green, T.; Kitts, D.D. Thermal oxidation studies on reduced folate, L-5-methyltetrahydrofolic acid (L-5-MTHF) and strategies for stabilization using food matrices. J. Food Sci. 2012, 77, C236–C243. [Google Scholar] [CrossRef]
- Liang, L.; Zhang, J.; Zhou, P.; Subirade, M. Protective effect of ligand-binding proteins against folic acid loss due to photodecomposition. Food Chem. 2013, 141, 754–761. [Google Scholar] [CrossRef]
- Cheng, H.; Dong, H.; Wusigale; Liang, L. A comparison of β-casein complexes and micelles as vehicles for trans-/cis-resveratrol. Food Chem. 2020, 330, 127209. [Google Scholar] [CrossRef] [PubMed]
- Off, M.K.; Steindal, A.E.; Porojnicu, A.C.; Juzeniene, A.; Vorobey, A.; Johnsson, A.; Moan, J. Ultraviolet photodegradation of folic acid. J. Photochem. Photobiol. B 2005, 80, 47–55. [Google Scholar] [CrossRef]
- An, J.; Zhang, Y.; Ying, Z.; Li, H.; Liu, W.; Wang, J.; Liu, X. The Formation, Structural Characteristics, Absorption Pathways and Bioavailability of Calcium–Peptide Chelates. Foods 2022, 11, 2762. [Google Scholar] [CrossRef]
- Zhou, Z.; Xu, J.; Liu, Y.; Meng, D.; Sun, X.; Yi, H.; Gao, Y.; Sun, G.; Strappe, P.; Blanchard, C.; et al. Thermal stability improvement of rice bran albumin protein incorporated with epigallocatechin gallate. J. Food Sci. 2017, 82, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhong, Q. Encapsulation of bixin in sodium caseinate to deliver the colorant in transparent dispersions. Food Hydrocoll. 2013, 33, 1–9. [Google Scholar] [CrossRef]
- Liu, M.; Liu, T.; Shi, Y.; Zhao, Y.; Yan, H.; Sun, B.; Wang, Q.; Wang, Z.; Han, J. Comparative study on the interaction of oxyresveratrol and piceatannol with trypsin and lysozyme: Binding ability, activity and stability. Food Funct. 2019, 10, 8182–8194. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | T (K) | Kq (×1012 L·mol−1·s−1) | Ka (×104 M−1) | n | ΔG (kJ·mol−1) | ΔH (kJ·mol−1) | ΔS (J·mol−1 K−1) |
|---|---|---|---|---|---|---|---|
| FA | 298.2 | 4.87 ± 0.03 | 7.84 ± 0.09 | 1.02 ± 0.01 | −27.92 ± 0.03 | 21.65 ± 0.49 | 166.30 ± 1.62 |
| 303.2 | 4.66 ± 0.01 | 9.00 ± 0.13 | 1.04 ± 0.02 | −28.74 ± 0.03 | |||
| 308.2 | 4.97 ± 0.02 | 10.41 ± 0.17 | 1.05 ± 0.01 | −29.58 ± 0.04 | |||
| MTFA | 298.2 | 5.09 ± 0.02 | 13.70 ± 0.23 | 1.07 ± 0.01 | −29.30 ± 0.04 | −45.62 ± 1.30 | −54.79 ± 4.29 |
| 303.2 | 4.25 ± 0.02 | 9.96 ± 0.16 | 1.06 ± 0.01 | −28.99 ± 0.04 | |||
| 308.2 | 3.90 ± 0.02 | 7.53 ± 0.05 | 1.04 ± 0.01 | −28.76 ± 0.02 | |||
| CaMTFA | 298.2 | 5.27 ± 0.02 | 13.68 ± 0.02 | 1.06 ± 0.01 | −29.30 ± 0.01 | −46.09 ± 1.96 | −56.28 ± 6.47 |
| 303.2 | 4.24 ± 0.03 | 10.29 ± 0.05 | 1.06 ± 0.01 | −29.07 ± 0.01 | |||
| 308.2 | 3.76 ± 0.02 | 7.48 ± 0.06 | 1.04 ± 0.01 | −28.73 ± 0.02 |
| Formulation | α-Helix (%) | β-Sheet (%) | β-Turn (%) | Random Coil (%) |
|---|---|---|---|---|
| NaCas | 19.60 ± 0.14 | 25.71 ± 1.04 | 36.06% ± 1.27 | 18.63 ± 0.31 |
| NaCas-FA | 19.52 ± 0.10 | 26.36 ± 0.01 | 35.45% ± 0.26 | 18.67 ± 0.23 |
| NaCas-MTFA | 19.80 ± 0.27 | 25.59 ± 1.15 | 35.93% ± 1.15 | 18.68 ± 0.27 |
| NaCas-CaMTFA | 19.42 ± 0.07 | 26.09 ± 1.29 | 35.93% ± 1.34 | 18.56 ± 0.12 |
| Condition | NaCas | NaCas + FA | ||
|---|---|---|---|---|
| Particle Size (nm) | ζ-Potential (mV) | Particle Size (nm) | ζ-Potential (mV) | |
| 0 days | 168.6 ± 7.0 | −16.4 ± 1.2 | 165.6 ± 6.1 | −17.4 ± 0.8 |
| 7 days in light | 168.0 ± 0.4 | −17.8 ± 0.8 | 137.4 ± 5.4 | −21.7 ± 1.6 |
| 49 days in dark | 167.7 ± 3.8 | −16.7 ± 1.7 | 144.0 ± 4.4 | −19.1 ± 1.2 |
| Condition | NaCas + MTFA | NaCas + CaMTFA | ||
| Particle size (nm) | ζ-potential (mV) | Particle size (nm) | ζ-potential (mV) | |
| 0 days | 172.6 ± 5.1 | −16.3 ± 0.9 | 174.6 ± 0.3 | −16.3 ± 0.2 |
| 3 days in light | 139.2 ± 2.3 | −26.2 ± 0.5 | 137.8 ± 2.0 | −26.4 ± 1.4 |
| 7 days in dark | 142.8 ± 6.2 | −23.6 ± 0.3 | 144.9 ± 1.6 | −23.1 ± 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Yan, Y.; Zhang, G.; Zhao, Y.; Zhao, F.; Ding, Z.; Wang, Z. Non-Covalent Interaction of Folic Acid and 5-Methyltetrahydrofolate with Caseinates Improves the Folates Stability Studied by Multi-Spectroscopic Analysis and Molecular Docking. Foods 2024, 13, 2756. https://doi.org/10.3390/foods13172756
He L, Yan Y, Zhang G, Zhao Y, Zhao F, Ding Z, Wang Z. Non-Covalent Interaction of Folic Acid and 5-Methyltetrahydrofolate with Caseinates Improves the Folates Stability Studied by Multi-Spectroscopic Analysis and Molecular Docking. Foods. 2024; 13(17):2756. https://doi.org/10.3390/foods13172756
Chicago/Turabian StyleHe, Linlin, Yuqian Yan, Gang Zhang, Yanna Zhao, Fa Zhao, Zhuang Ding, and Zhengping Wang. 2024. "Non-Covalent Interaction of Folic Acid and 5-Methyltetrahydrofolate with Caseinates Improves the Folates Stability Studied by Multi-Spectroscopic Analysis and Molecular Docking" Foods 13, no. 17: 2756. https://doi.org/10.3390/foods13172756
APA StyleHe, L., Yan, Y., Zhang, G., Zhao, Y., Zhao, F., Ding, Z., & Wang, Z. (2024). Non-Covalent Interaction of Folic Acid and 5-Methyltetrahydrofolate with Caseinates Improves the Folates Stability Studied by Multi-Spectroscopic Analysis and Molecular Docking. Foods, 13(17), 2756. https://doi.org/10.3390/foods13172756