
Effective Degradation of Free Gossypol in Defatted Cottonseed Meal by Bacterial Laccases: Performance and Toxicity Analysis
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Expression and Purification of Laccases
2.3. Characterization of FG Degradation Capacity of Laccases
2.4. Quantification of FG by HPLC
2.5. Molecular Docking Study
2.6. Cytotoxicity Assessment of FG Degradation Products
2.7. In Vivo Toxicity Assessment of FG Degradation Products
2.8. Application of Laccase to Degrade FG in Defatted Cottonseed Meal
2.9. Statistical Analysis
3. Results and Discussion
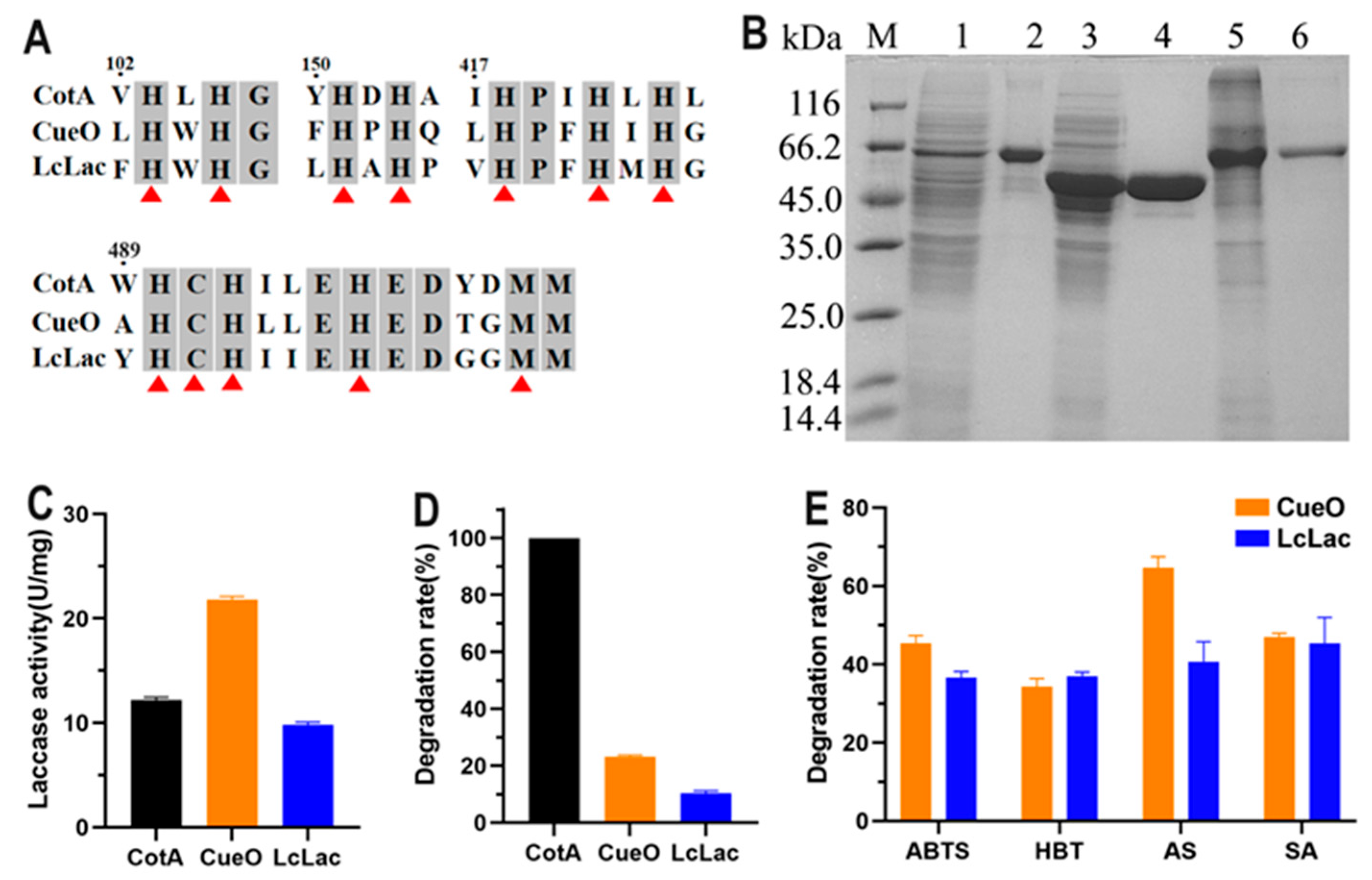
3.1. Expression of Three Bacterial Laccases and Investigation of their FG-Degrading Capacity
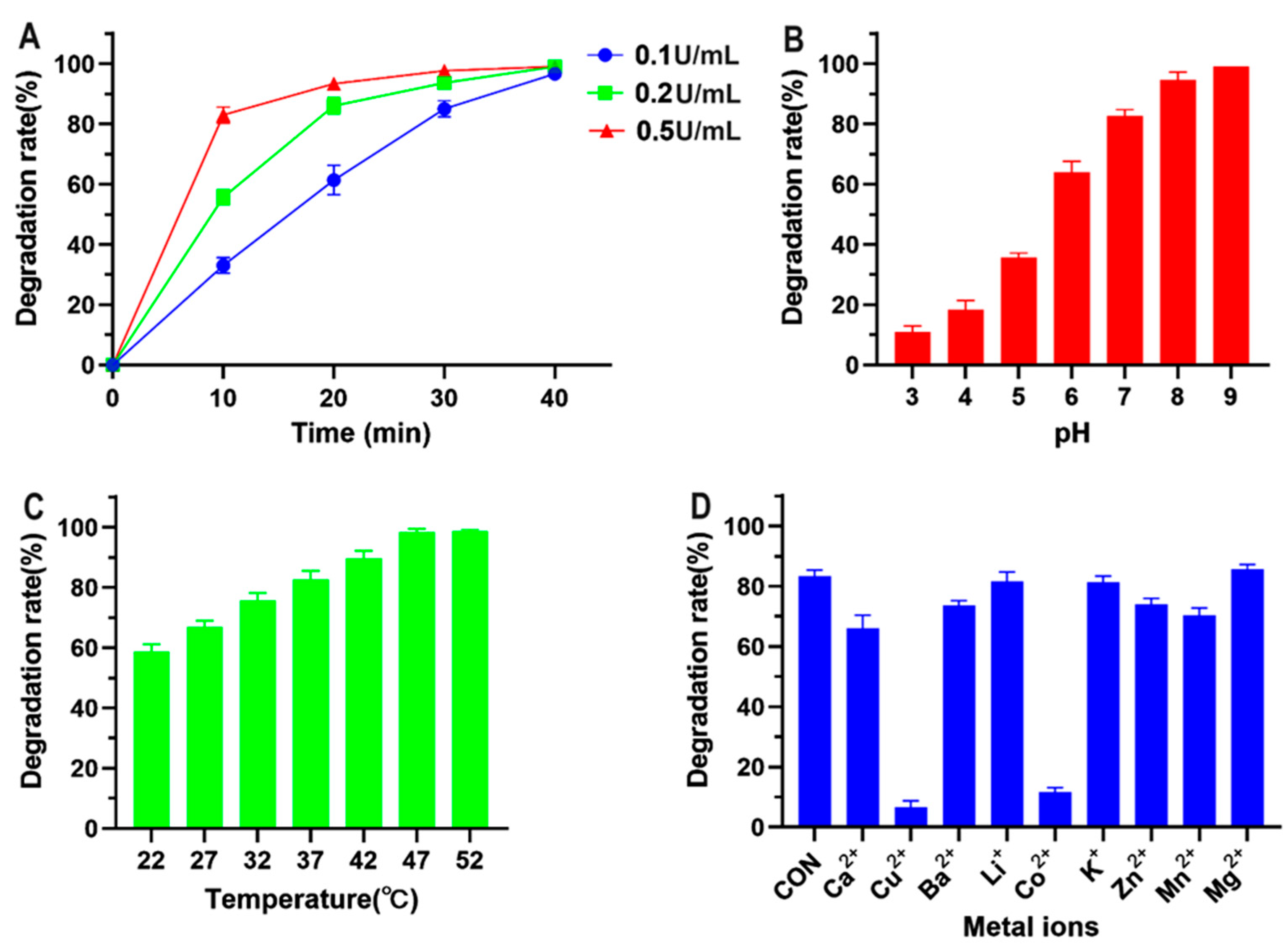
3.2. Enzymatic Characterization of CotA Laccase for Directly Oxidizing FG
3.3. Molecular Docking of FG with CotA Laccase
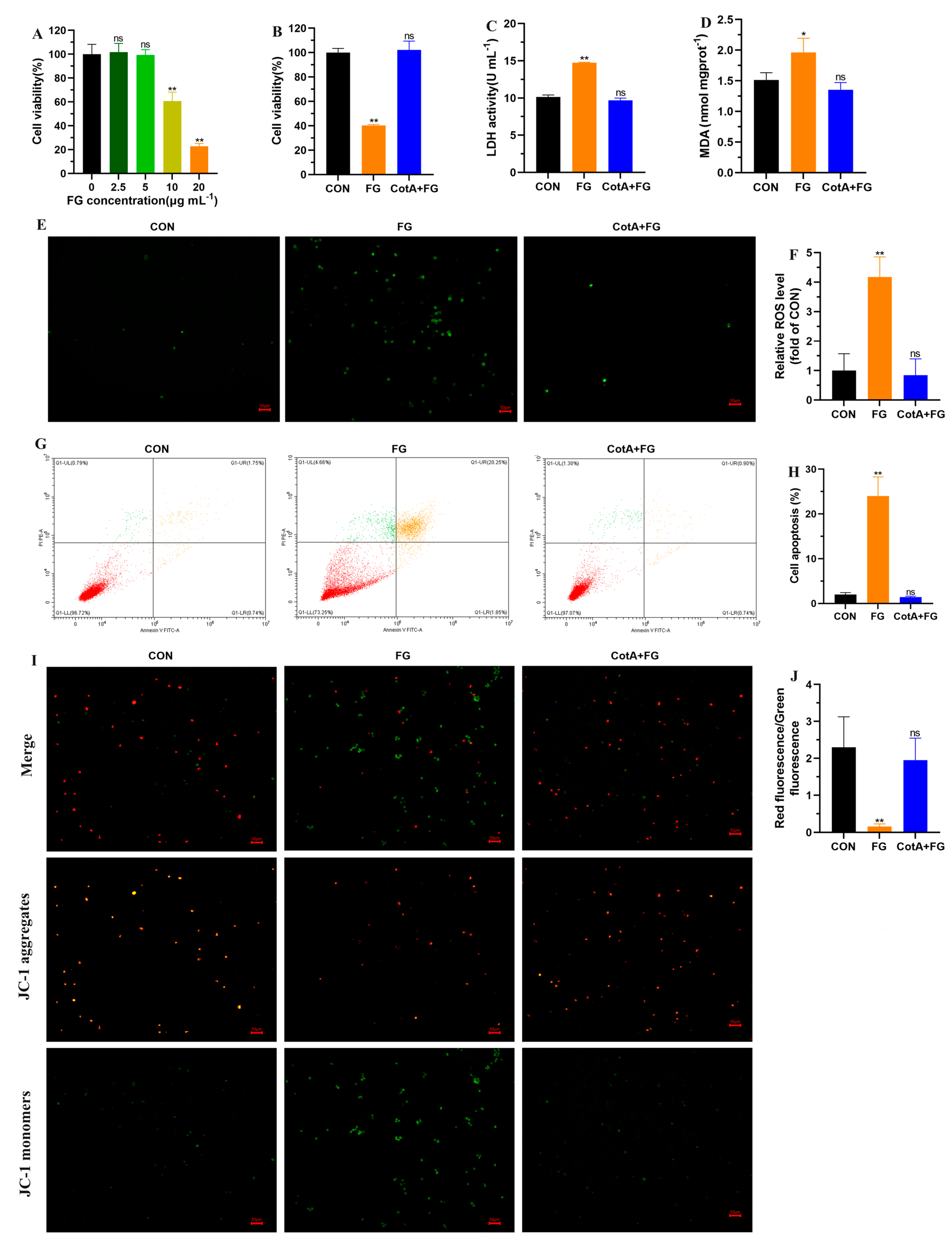
3.4. Cytotoxicity Assessment of FG Degradation Products
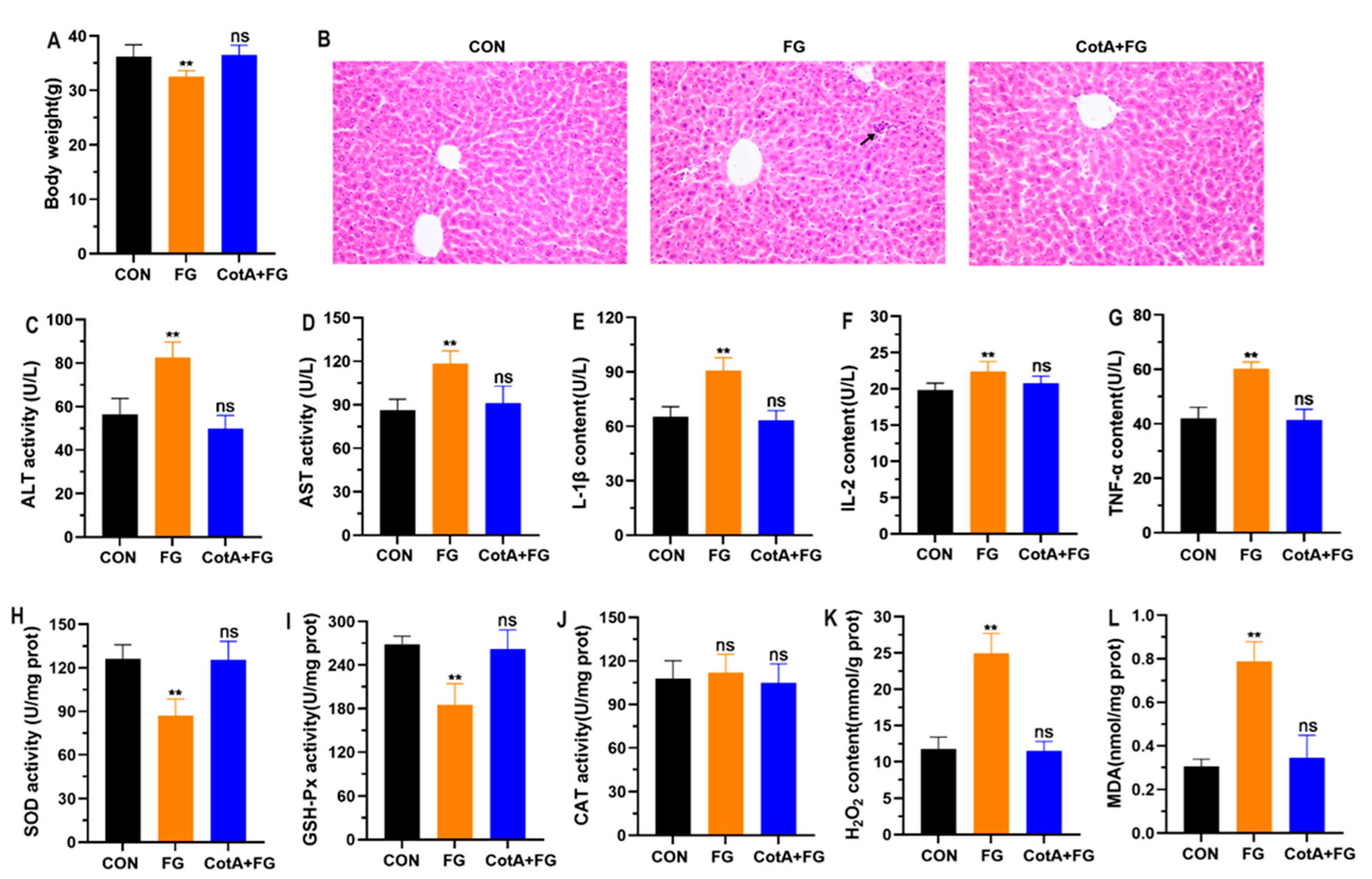
3.5. In Vivo Toxicity Assessment of FG Degradation Products
3.6. Application of CotA Laccase to Degrade FG in Defatted Cottonseed Meal
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Langyan, S.; Yadava, P.; Khan, F.N.; Dar, Z.A.; Singh, R.; Kumar, A. Sustaining protein nutrition through plant-based foods. Front. Nutr. 2021, 8, 772573. [Google Scholar] [CrossRef] [PubMed]
- Ninkuu, V.; Liu, Z.X.; Zhou, Y.P.; Sun, X.W. The nutritional and industrial significance of cottonseeds and genetic techniques in gossypol detoxification. New Phytol. 2023, 10, 10433. [Google Scholar] [CrossRef]
- He, Z.; Cheng, H.N.; He, J. Initial formulation of novel peanut butter-like products from glandless cottonseed. Foods 2023, 12, 378. [Google Scholar] [CrossRef]
- He, Z.; Rogers, S.I.; Nam, S.; Klasson, K.T. The effects of oil content on the structural and textural properties of cottonseed butter/spread products. Foods 2023, 12, 4158. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.K.; Yang, H.J.; Wang, Y.L.; Yang, K.L.; Jiang, L.S.; Li, S.L. Gossypol detoxification in the rumen and Helicoverpa armigera larvae: A review. Anim. Nutr. 2021, 7, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, I.C.; Fonseca, N.B.; Oloris, S.C.; Melo, M.M.; Soto-Blanco, B. Gossypol toxicity from cottonseed products. Sci. World J. 2014, 2014, 231635. [Google Scholar] [CrossRef]
- Hatamoto-Zervoudakis, L.K.; Duarte Júnior, M.F.; Zervoudakis, J.T.; Motheo, T.F.; Silva-Marques, R.P.; Tsuneda, P.P.; Nichi, M.; Santo, B.S.E.; Almeida, R.D. Free gossypol supplementation frequency and reproductive toxicity in young bulls. Theriogenology 2018, 110, 153–157. [Google Scholar] [CrossRef]
- Ding, Z.M.; Chen, Y.W.; Wang, Y.S.; Ahmad, M.J.; Yang, S.J.; Duan, Z.Q.; Liu, M.; Yang, C.X.; Xiong, J.J.; Liang, A.X.; et al. Gossypol exposure induces mitochondrial dysfunction and oxidative stress during mouse oocyte in vitro maturation. Chem. Biol. Interact. 2021, 348, 109642. [Google Scholar] [CrossRef]
- Villaseñor, M.; Coscioni, A.C.; Galvão, K.N.; Chebel, R.C.; Santos, J.E. Gossypol disrupts embryo development in heifers. J. Dairy Sci. 2008, 91, 3015–3024. [Google Scholar] [CrossRef]
- Kumar, M.; Tomar, M.; Punia, S.; Grasso, S.; Arrutia, F.; Choudhary, J.; Singh, S.; Verma, P.; Mahapatra, A.; Patil, S.; et al. Cottonseed: A sustainable contributor to global protein requirements. Trends Food Sci. Technol. 2021, 111, 100–113. [Google Scholar] [CrossRef]
- Jiang, Y.; Du, X.; Xu, Q.; Yin, C.; Zhang, H.; Liu, Y.; Liu, X.; Yan, H. Biodegradation of gossypol by Aspergillus terreus-YJ01. Microorganisms 2023, 11, 2148. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.K.; Li, W.J.; Wu, Q.C.; Wang, Y.L.; Li, S.L.; Yang, H.J. Isolation and identification of a rumen Lactobacillus bacteria and its degradation potential of gossypol in cottonseed meal during solid-state fermentation. Microorganisms 2021, 9, 2200. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, W.; Zhang, H.; Cui, Y.; Wang, F.; Wu, J.; Chao, H.; Yan, D. The complete genome sequence of a gossypol-degrading bacterial strain, Raoultella sp. YL01. Curr. Microbiol. 2023, 80, 163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, D.; Liu, L.; Chang, Z.; Peng, N. Effective gossypol removal from cottonseed meal through optimized solid-state fermentation by Bacillus coagulans. Microb. Cell Factories 2022, 21, 252. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gao, T.; Hao, Z.; Guo, X.; Zhu, B. Anaerobic solid-state fermentation with Bacillus subtilis for digesting free gossypol and improving nutritional quality in cottonseed meal. Front. Nutr. 2022, 9, 1017637. [Google Scholar] [CrossRef]
- Chen, C.; Pi, W.; Zhang, Y.; Nie, C.X.; Liang, J.; Ma, X.; Wang, Y.; Ge, W.; Zhang, W.J. Effect of a functional recombinant cytochrome P450 enzyme of Helicoverpa armigera on gossypol metabolism co-expressed with NADPH-cytochrome P450 reductase in Pichia pastoris. Pestic. Biochem. Physiol. 2019, 155, 15–25. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, X.; Huang, R.; Nie, C.; Niu, J.; Chen, C.; Zhang, W. Biodegradation of free gossypol by Helicoverpa armigera carboxylesterase expressed in Pichia pastoris. Toxins 2022, 14, 816. [Google Scholar] [CrossRef]
- Yang, J.; Li, W.; Ng, T.B.; Deng, X.; Lin, J.; Ye, X. Laccases: Production, expression regulation, and applications in pharmaceutical biodegradation. Front. Microbiol. 2017, 8, 832. [Google Scholar] [CrossRef]
- Martins, L.O.; Durão, P.; Brissos, V.; Lindley, P.F. Laccases of prokaryotic origin: Enzymes at the interface of protein science and protein technology. Cell. Mol. Life Sci. 2015, 72, 911–922. [Google Scholar] [CrossRef]
- Guo, Y.; Qin, X.; Tang, Y.; Ma, Q.; Zhang, J.; Zhao, L. CotA laccase, a novel aflatoxin oxidase from Bacillus licheniformis, transforms aflatoxin B1 to aflatoxin Q1 and epi-aflatoxin Q1. Food Chem. 2020, 325, 126877. [Google Scholar] [CrossRef]
- Callejón, S.; Sendra, R.; Ferrer, S.; Pardo, I. Cloning and characterization of a new laccase from Lactobacillus plantarum J16 CECT 8944 catalyzing biogenic amines degradation. Appl. Microbiol. Biotechnol. 2016, 100, 3113–3124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X.; Hao, Z.; Xi, R.; Cai, Y.; Liao, X. Hydrogen peroxide-resistant CotA and YjqC of Bacillus altitudinis spores are a promising biocatalyst for catalyzing reduction of sinapic acid and sinapine in rapeseed meal. PLoS ONE 2016, 11, e0158351. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, M.; Luo, X.; Fan, Y.; Zheng, Z.; He, Z.; Yin, R.; Meng, T.; Xu, S.; Pan, Y.; et al. Intramolecular annulation of gossypol by laccase to produce safe cottonseed protein. Front. Chem. 2020, 8, 583176. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, A.; Rencoret, J.; Ibarra, D.; Molina, S.; Camarero, S.; Romero, J.; Del Río, J.C.; Martínez, A.T. Removal of lipophilic extractives from paper pulp by laccase and lignin-derived phenols as natural mediators. Environ. Sci. Technol. 2007, 41, 4124–4129. [Google Scholar] [CrossRef]
- Lin, H.; Yu, Z.J.; Wang, Q.; Liu, Y.J.; Jiang, L.; Xu, C.; Xian, M. Application of laccase catalysis in bond formation and breakage: A review. Catalysts 2023, 13, 750. [Google Scholar] [CrossRef]
- Guo, Y.P.; Wang, Y.A.; Liu, Y.R.; Ma, Q.G.; Ji, C.; Zhao, L.H. Detoxification of the mycoestrogen zearalenone by spore CotA laccase and application of immobilized laccase in contaminated corn meal. LWT 2022, 163, 113548. [Google Scholar] [CrossRef]
- Khan, S.I.; Sahinkaya, M.; Colak, D.N.; Zada, N.S.; Uzuner, U.; Belduz, A.O.; Çanakçi, S.; Khan, A.Z.; Khan, S.; Badshah, M.; et al. Production and characterization of novel thermostable CotA-laccase from Bacillus altitudinis SL7 and its application for lignin degradation. Enzym. Microb. Technol. 2024, 172, 110329. [Google Scholar] [CrossRef]
- Cheng, C.M.; Patel, A.K.; Singhania, R.R.; Tsai, C.H.; Chen, S.Y.; Chen, C.W.; Dong, C.D. Heterologous expression of bacterial CotA-laccase, characterization and its application for biodegradation of malachite green. Bioresour. Technol. 2021, 340, 125708. [Google Scholar] [CrossRef]
- Wang, H.; Huang, L.; Li, Y.; Ma, J.; Wang, S.; Zhang, Y.; Ge, X.; Wang, N.; Lu, F.; Liu, Y. Characterization and application of a novel laccase derived from Bacillus amyloliquefaciens. Int. J. Biol. Macromol. 2020, 150, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Yu, D.Z.; Zhou, H.Y.; Lin, H.K.; Yan, Z.; Wu, A.B. CotA laccase from Bacillus licheniformis ZOM-1 effectively degrades zearalenone, aflatoxin B1 and alternariol. Food Control 2023, 145, 109472. [Google Scholar] [CrossRef]
- Fonseca, N.B.; Gadelha, I.C.; Oloris, S.C.; Soto-Blanco, B. Effectiveness of albumin-conjugated gossypol as an immunogen to prevent gossypol-associated acute hepatotoxicity in rats. Food Chem. Toxicol. 2013, 56, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Blevins, S.; Siegel, P.B.; Blodgett, D.J.; Ehrich, M.; Saunders, G.K.; Lewis, R.M. Effects of silymarin on gossypol toxicosis in divergent lines of chickens. Poult. Sci. 2010, 89, 1878–1886. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jin, L.; Li, X.; Deng, H.; Chen, Y.; Lian, Q.; Ge, R.; Deng, H. Gossypol induces apoptosis in ovarian cancer cells through oxidative stress. Mol. Biosyst. 2013, 9, 1489–1497. [Google Scholar] [CrossRef] [PubMed]
- El-Sharaky, A.S.; Newairy, A.A.; Elguindy, N.M.; Elwafa, A.A. Spermatotoxicity, biochemical changes and histological alteration induced by gossypol in testicular and hepatic tissues of male rats. Food Chem. Toxicol. 2010, 48, 3354–3361. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Y.; Pi, W.; Yang, W.; Nie, C.; Liang, J.; Ma, X.; Zhang, W.J. Optimization of the process parameters for reduction of gossypol levels in cottonseed meal by functional recombinant NADPH-cytochrome P450 reductase and cytochrome P450 CYP9A12 of Helicoverpa armigera. AMB Express 2019, 9, 98. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zheng, H.; Zhang, X.; Chen, X.; Liu, Y.; Tang, Y.; Zhang, W.; Wang, Z.; Zhao, L.; Guo, Y. Effective Degradation of Free Gossypol in Defatted Cottonseed Meal by Bacterial Laccases: Performance and Toxicity Analysis. Foods 2024, 13, 566. https://doi.org/10.3390/foods13040566
Zhang L, Zheng H, Zhang X, Chen X, Liu Y, Tang Y, Zhang W, Wang Z, Zhao L, Guo Y. Effective Degradation of Free Gossypol in Defatted Cottonseed Meal by Bacterial Laccases: Performance and Toxicity Analysis. Foods. 2024; 13(4):566. https://doi.org/10.3390/foods13040566
Chicago/Turabian StyleZhang, Liangyu, Hao Zheng, Xingke Zhang, Xiaoxue Chen, Yanrong Liu, Yu Tang, Wei Zhang, Zhixiang Wang, Lihong Zhao, and Yongpeng Guo. 2024. "Effective Degradation of Free Gossypol in Defatted Cottonseed Meal by Bacterial Laccases: Performance and Toxicity Analysis" Foods 13, no. 4: 566. https://doi.org/10.3390/foods13040566