
Co-Immobilization of Alcalase/Dispase for Production of Selenium-Enriched Peptide from Cardamine violifolia
 ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Se-Enriched Protein from CV
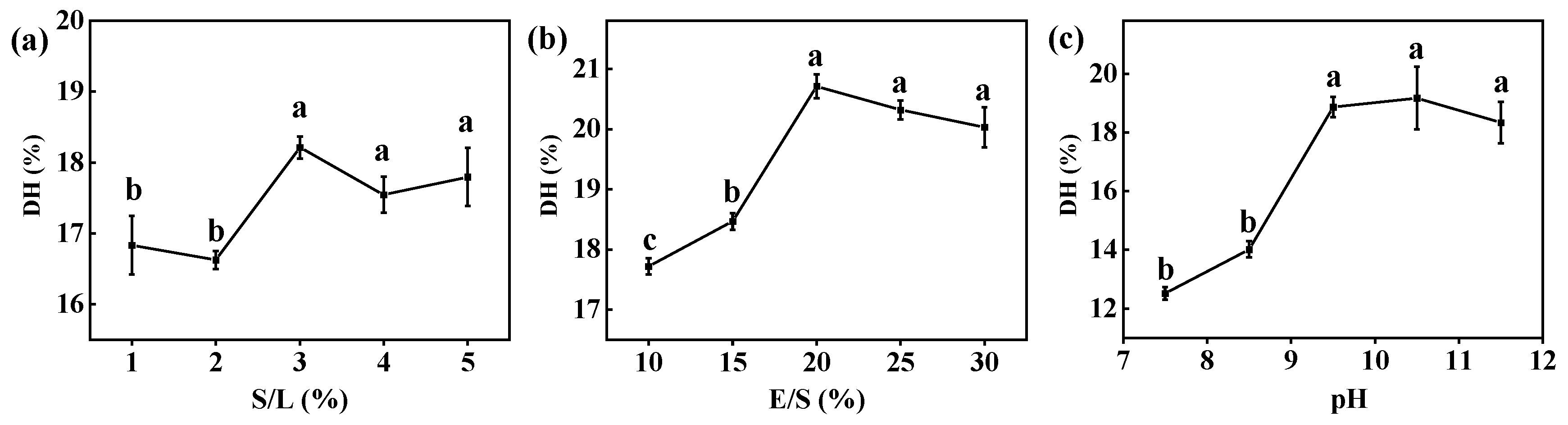
2.3. Optimizing Enzymatic Hydrolysis of CV Protein by Free Alcalase
2.4. Preparation of Co-Immobilized Enzyme
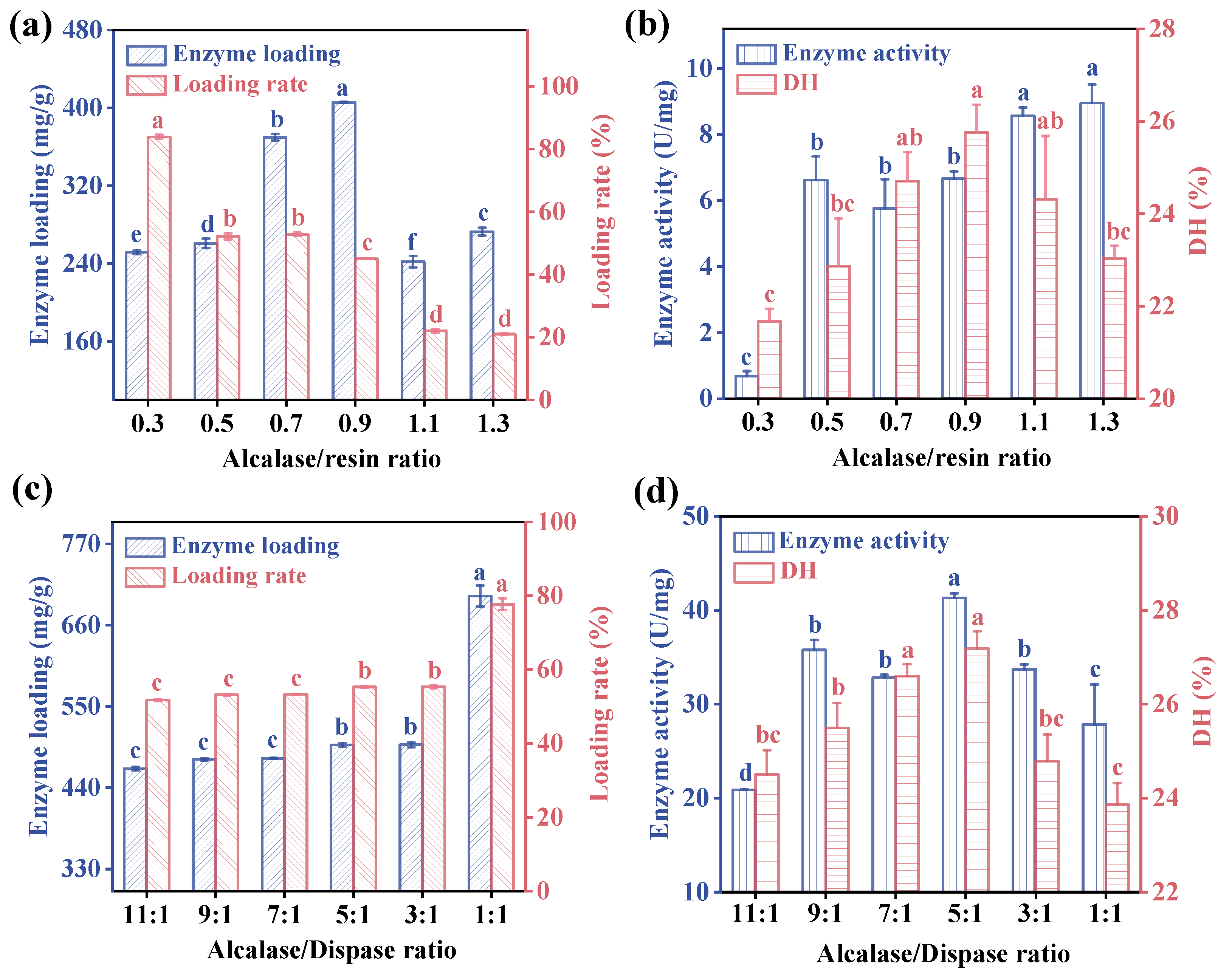
2.4.1. Determination of the Optimal Alcalase/Resin Ratio
2.4.2. Determination of the Optimal Ratio of Two Proteases
2.5. Characterization
2.6. Enzymatic Properties Assay of Free and Co-Immobilized Enzyme
2.6.1. Examination of the Stability
2.6.2. Kinetic Parameters of Enzyme
2.7. Analysis of Hydrolysis Products
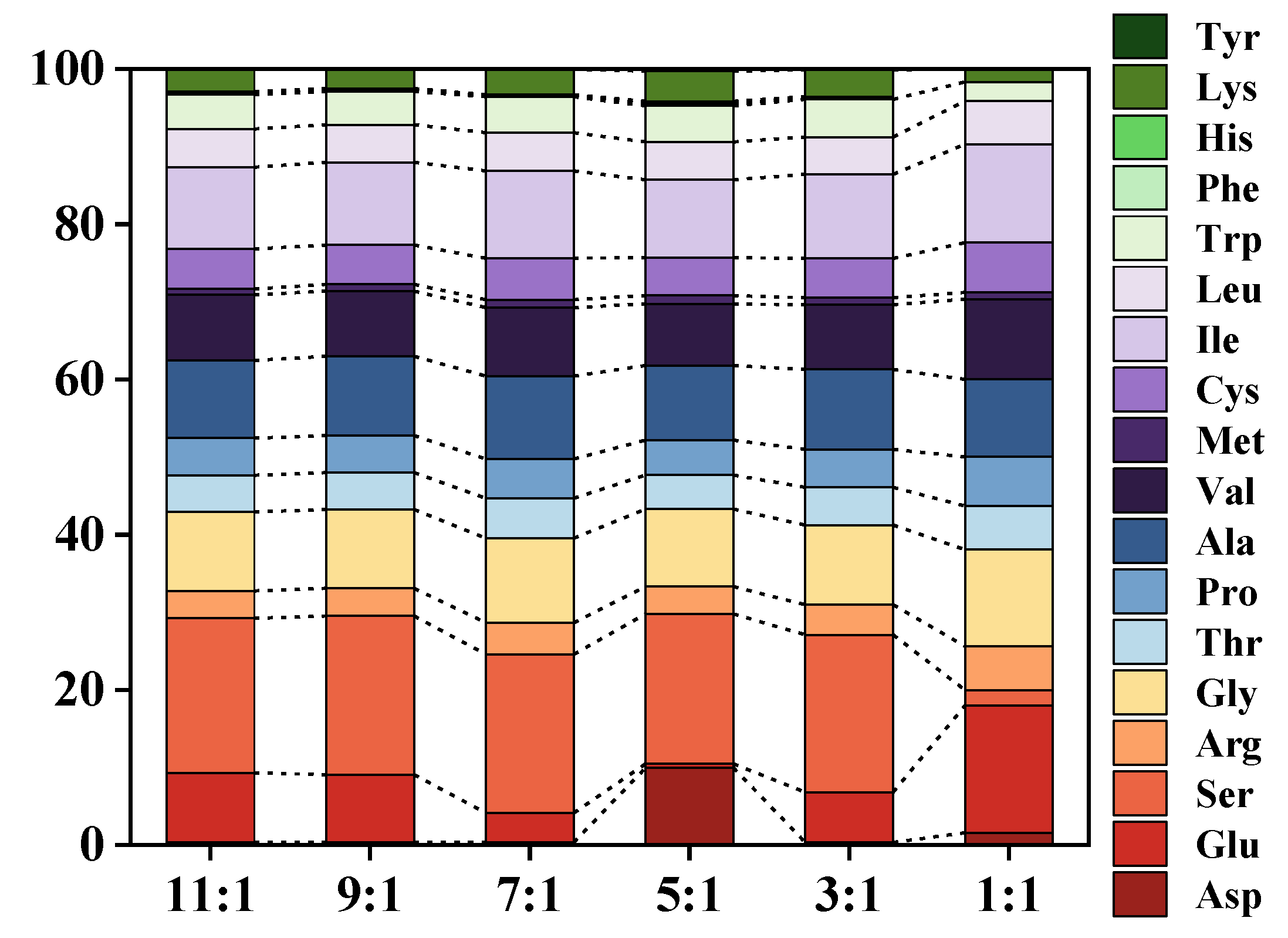
2.7.1. Measurement of Amino Acid Fraction
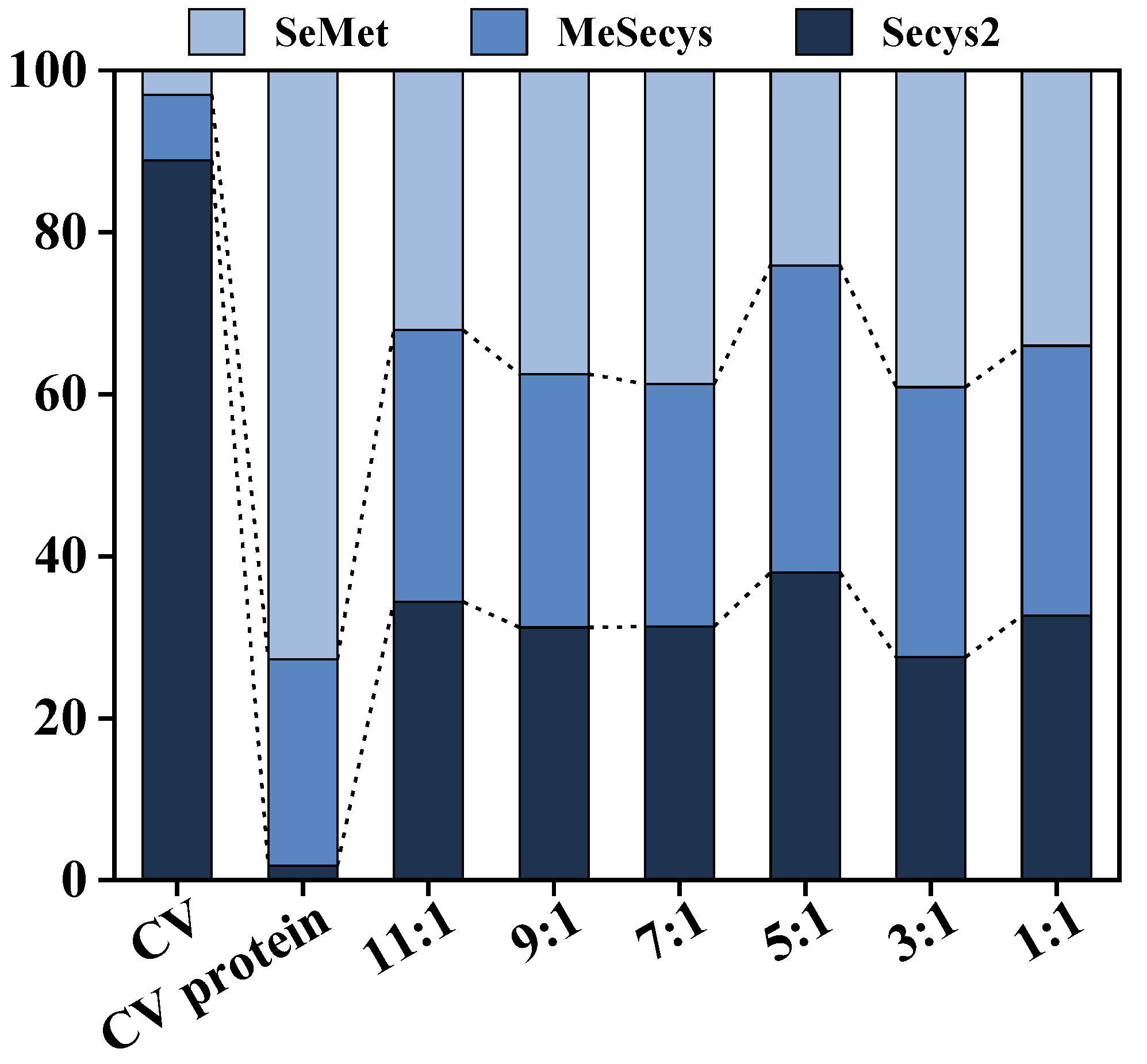
2.7.2. Determination of Se Content
2.7.3. Identification of Se-Enriched Peptide Sequences
2.8. Statistical Analysis
3. Results and Discussion
3.1. Enzyme Immobilization
3.1.1. Single-Factor Experiment of Free Alcalase
3.1.2. Determination of the Optimal Alcalase/Resin and Alcalase/Dispase Ratio
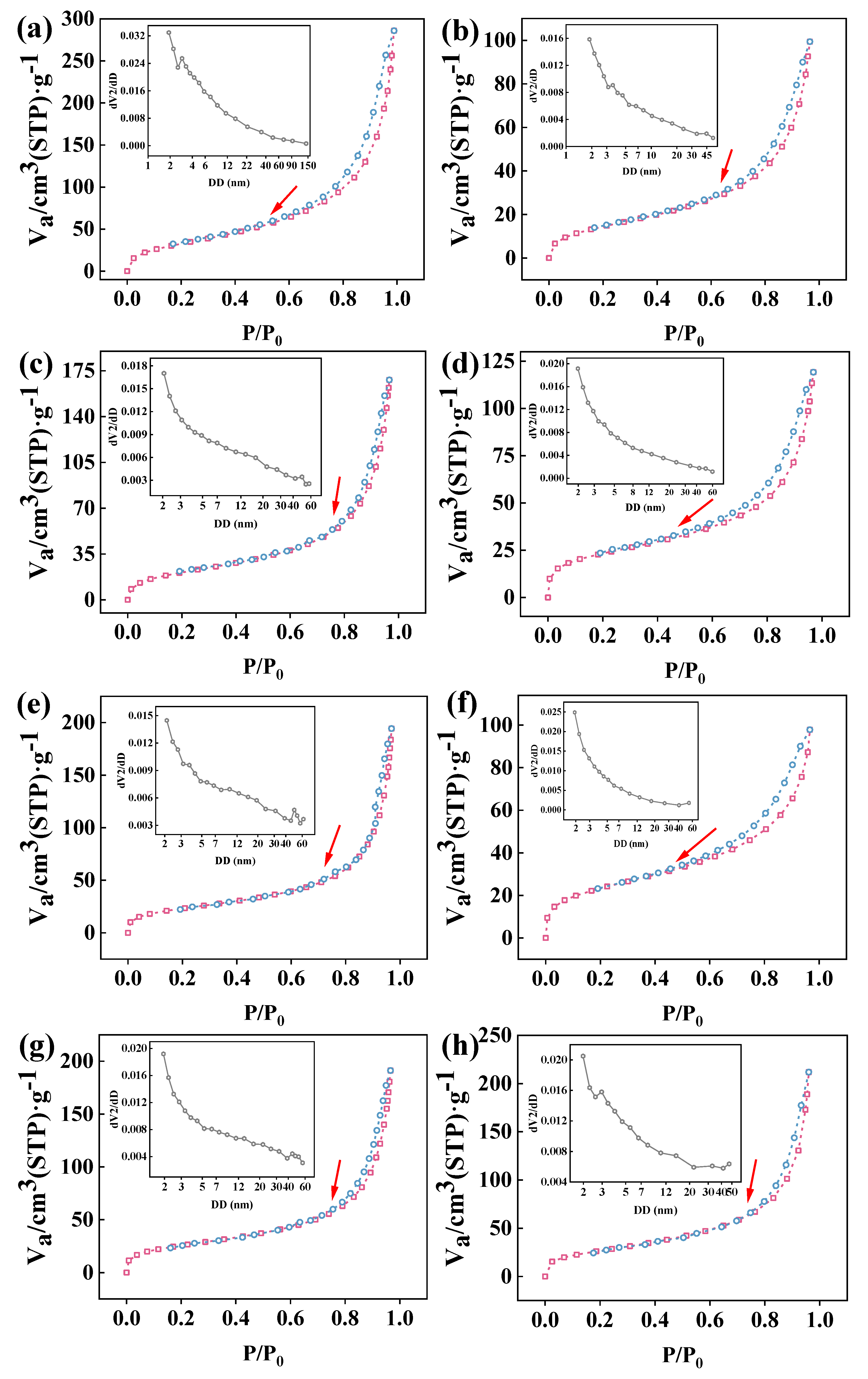
3.2. N2 Adsorption–Desorption and Pore Size Distribution of Co-Immobilized Enzyme
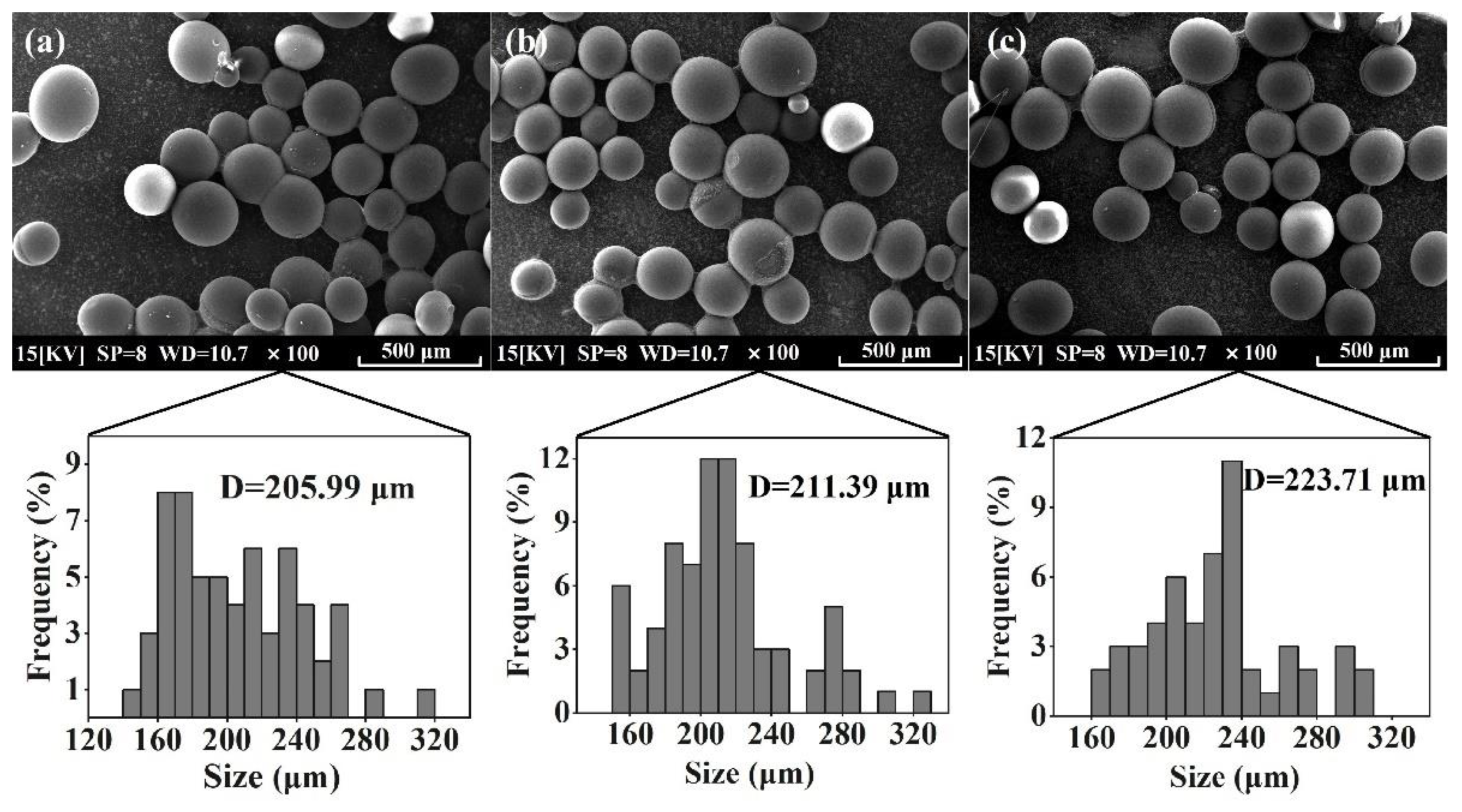
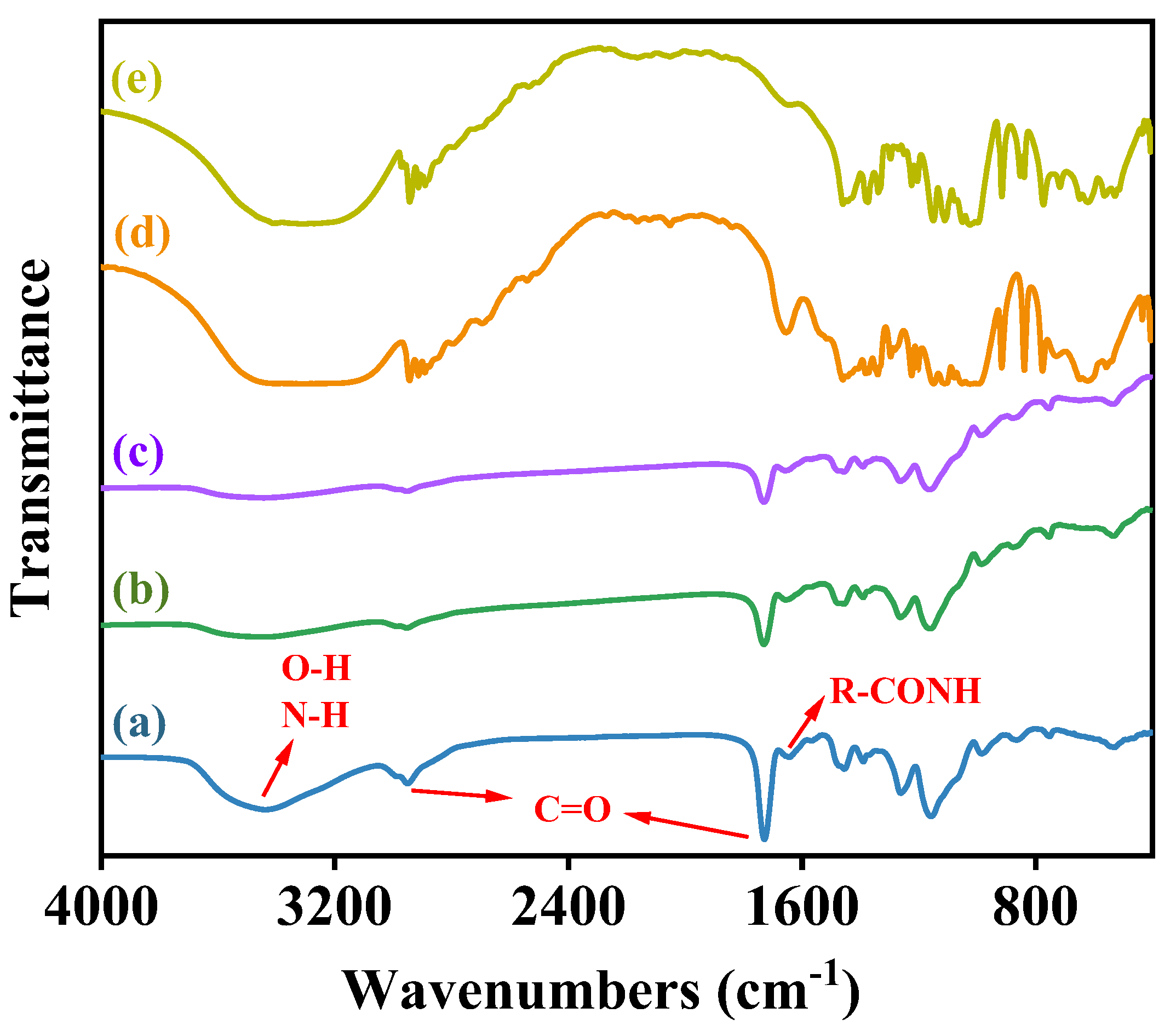
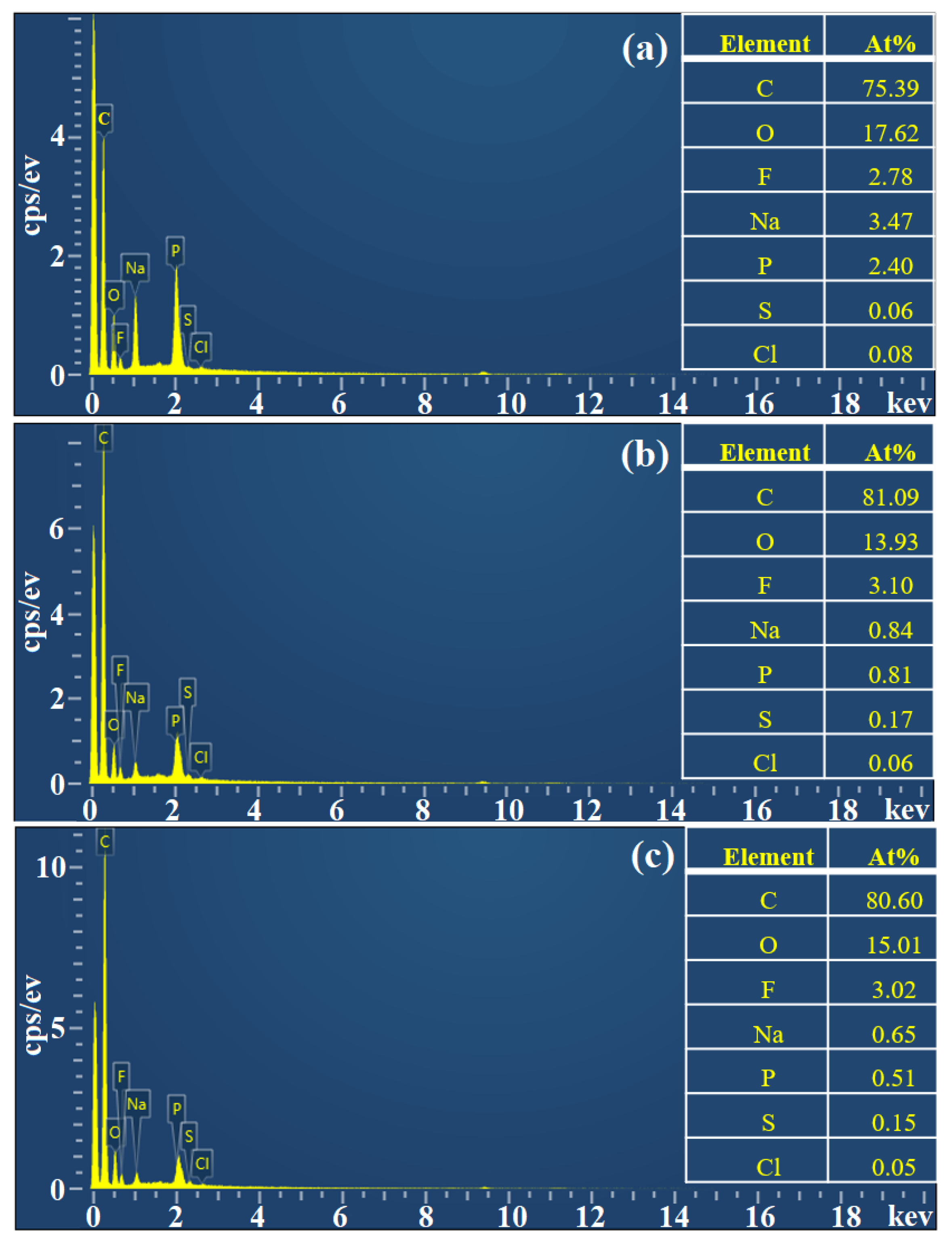
3.3. Characterization of Co-Immobilized Enzyme
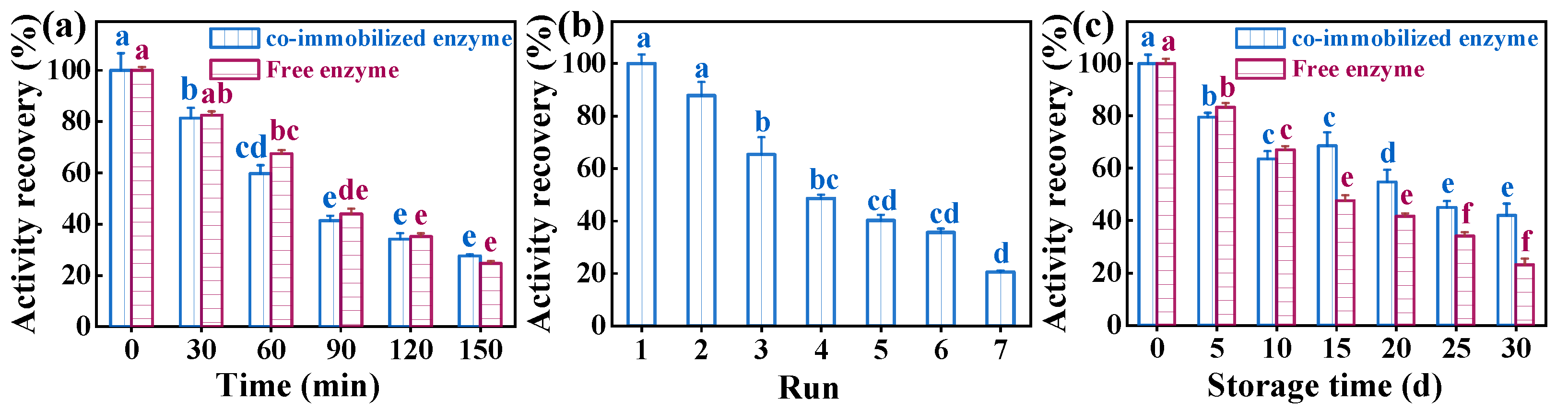
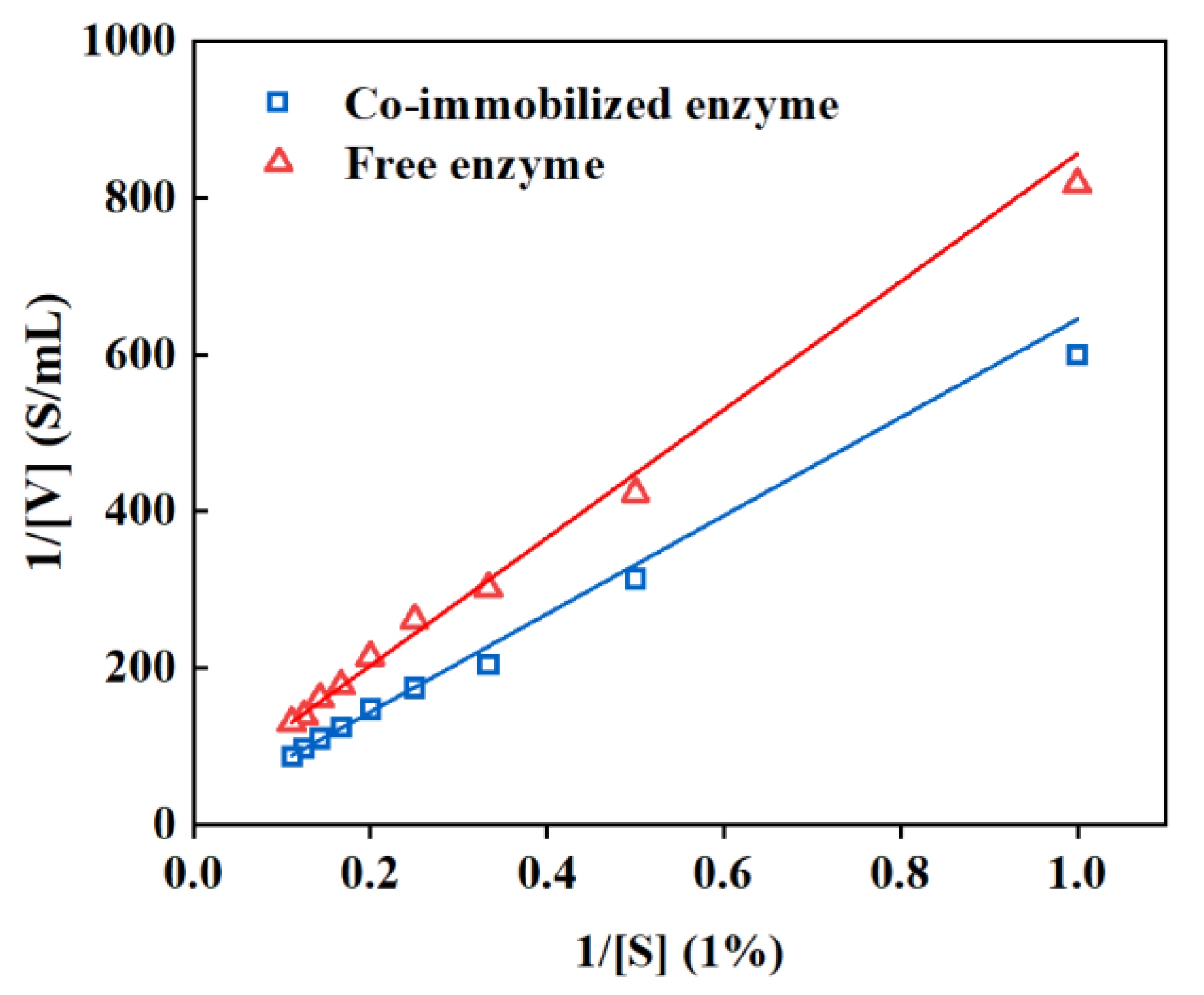
3.4. Examination of the Performance of Co-Immobilized Enzyme
3.5. Determination of Enzymatic Digestion Products
3.6. Comparison of the Efficiency of Different Immobilized Enzymes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ermakov, V.; Jovanović, L. Selenium deficiency as a consequence of human activity and its correction. J. Geochem. Explor. 2010, 107, 193–199. [Google Scholar] [CrossRef]
- Tangjaidee, P.; Swedlund, P.; Xiang, J.; Yin, H.; Quek, S.Y. Selenium-enriched plant foods: Selenium accumulation, speciation, and health functionality. Front. Nutr. 2023, 9, 962312–962330. [Google Scholar] [CrossRef] [PubMed]
- Estevez, H.; Garcia-Calvo, E.; Rivera-Torres, J.; Vallet-Regí, M.; González, B.; Luque-Garcia, J.L. Transcriptome Analysis Identifies Novel Mechanisms Associated with the Antitumor Effect of Chitosan-Stabilized Selenium Nanoparticles. Pharmaceutics 2021, 13, 356–375. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wei, W.; Wang, J.; Chen, T. Theranostic applications of selenium nanomedicines against lung cancer. J. Nanobiotechnol. 2023, 21, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, M.; Szczyglowska, M.; Konieczka, P.; Namiesnik, J. Methods of Selenium Supplementation: Bioavailability and Determination of Selenium Compounds. Crit. Rev. Food Sci. 2014, 56, 36–55. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.G.; Evenson, J.K.; Thompson, K.M.; Sunde, R.A. Glutathione peroxidase and phospholipid hydroperoxide glutathione peroxidase are differentially regulated in rats by dietary selenium. J. Nutr. 1995, 125, 1438–1446. [Google Scholar] [PubMed]
- Nasim, M.J.; Zuraik, M.M.; Abdin, A.Y.; Ney, Y.; Jacob, C. Selenomethionine: A Pink Trojan Redox Horse with Implications in Aging and Various Age-Related Diseases. Antioxidants 2021, 10, 882. [Google Scholar] [CrossRef]
- Liu, C.; Xu, L.; Tian, H.; Yao, H.; Elding, L.I.; Shi, T. Kinetics and mechanism for reduction of Pt(IV) anticancer model compounds by Se-methyl L-selenocysteine. Comparison with L-selenomethionine. J. Mol. Liq. 2018, 271, 838–843. [Google Scholar] [CrossRef]
- Rao, S.; Yu, T.; Cong, X.; Zhang, W.; Zhu, Z.; Liao, Y.; Ye, J.; Cheng, S.; Xu, F. Effects of selenate applied at two growth stages on the nutrient quality of Cardamine violifolia. Sci. Hortic. 2021, 288, 110352. [Google Scholar] [CrossRef]
- Geng, Z.; Wang, L.; Feng, Y.; Sun, Y.; Wang, S.; Shen, L. Isolation, Identification and Selenium Tolerance Assay of Endophytic Microbes from Selenium Hyperaccumulator Cardamine violifolia. J. Agric. Biotechnol. 2021, 29, 1795–1807. (In Chinese) [Google Scholar]
- Du, C.; Wang, P.; Li, Y.; Cong, X.; Huang, D.; Chen, S.; Zhu, S. Investigation of selenium and selenium species in Cardamine violifolia using in vitro digestion coupled with a Caco-2 cell monolayer model. Food Chem. 2024, 444, 138675–138684. [Google Scholar] [CrossRef] [PubMed]
- Both, E.B.; Shao, S.; Xiang, J.; Jókai, Z.; Yin, H.; Liu, Y.; Magyar, A.; Dernovics, M. Selenolanthionine is the major water-soluble selenium compound in the selenium tolerant plant Cardamine violifolia. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 2354–2362. [Google Scholar] [CrossRef] [PubMed]
- Lv, R.; Dong, Y.; Bao, Z.; Zhang, S.; Lin, S.; Sun, N. Advances in the activity evaluation and cellular regulation pathways of food-derived antioxidant peptides. Trends Food Sci. Tech. 2022, 122, 171–186. [Google Scholar]
- Ling, R.; Du, C.; Li, Y.; Wang, S.; Cong, X.; Huang, D.; Chen, S.; Zhu, S. Protective Effect of Selenium-enriched Peptide from Cardamine violifolia on Ethanol-induced L-02 Hepatocyte Injury. Biol. Trace Elem. Res. 2024, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; McClements, D.J.; Li, J.; Chang, C.; Su, Y.; Gu, L.; Yang, Y. Egg Yolk Selenopeptides: Preparation, Characterization, and Immunomodulatory Activity. J. Agric. Food Chem. 2024, 72, 5237–5246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gao, S.; Li, H.; Cao, M.; Li, W.; Liu, X. Immunomodulatory effects of selenium-enriched peptides from soybean in cyclophosphamide-induced immunosuppressed mice. Food Sci. Nutr. 2021, 9, 6322–6334. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Du, M.; Wu, M.; Yue, P.; Yang, X.; Wei, X.; Wang, Y. Preparation, physicochemical characterization and identification of two novel mixed ACE-inhibiting peptides from two distinct tea alkali-soluble protein. Eur. Food Res. Technol. 2020, 246, 1483–1494. [Google Scholar] [CrossRef]
- Chen, B.; Miao, J.; Ye, H.; Xia, Z.; Huang, W.; Guo, J.; Liang, X.; Yin, Y.; Zheng, Y.; Cao, Y. Purification, Identification, and Mechanistic Investigation of Novel Selenium-Enriched Antioxidant Peptides from Moringa oleifera Seeds. J. Agric. Food Chem. 2023, 71, 4625–4637. [Google Scholar] [CrossRef]
- Rybarczyk, A.; Smułek, W.; Grzywaczyk, A.; Kaczorek, E.; Jesionowski, T.; Nghiem, L.D.; Zdarta, J. 3D printed polylactide scaffolding for laccase immobilization to improve enzyme stability and estrogen removal from wastewater. Bioresour. Technol. 2023, 381, 129144–129153. [Google Scholar] [CrossRef]
- Guimarães, J.R.; Carballares, D.; Rocha-Martin, J.; Tardioli, P.W.; Fernandez-Lafuente, R. The immobilization protocol greatly alters the effects of metal phosphate modification on the activity/stability of immobilized lipases. Int. J. Biol. Macromol. 2022, 222, 2452–2466. [Google Scholar] [CrossRef]
- Anwar, N.; Yuxia, W.; Chao, Z.; Liguo, Y.; Sherif, M.A. Fractionation and purification of antioxidant peptides from Chinese sturgeon (Acipenser sinensis) protein hydrolysates prepared using papain and alcalase 2.4 L. Arab. J. Chem. 2022, 15, 104368–104377. [Google Scholar]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.-H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [PubMed]
- Zhang, X.; He, H.; Xiang, J.; Li, B.; Zhao, M.; Hou, T. Selenium-containing soybean antioxidant peptides: Preparation and comprehensive comparison of different selenium supplements. Food Chem. 2021, 358, 129888–129897. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, H.; Wang, J.; Xing, L. Preparation of active corn peptides from zein through double enzymes immobilized with calcium alginate–chitosan beads. Process Biochem. 2014, 49, 1682–1690. [Google Scholar] [CrossRef]
- Wu, M.; Zhu, Z.; Li, S.; Cai, J.; Cong, X.; Yu, T.; Yang, W.; He, J.; Cheng, S. Green recovery of Se-rich protein and antioxidant peptides from Cardamine Violifolia: Composition and bioactivity. Food Biosci. 2020, 38, 100743–100751. [Google Scholar] [CrossRef]
- Du, C.; Yu, T.; Cong, X.; Chen, S.; Zhu, S. Preparation of Cardamine Protein and Its Antioxidant Activity. J. Food Sci. Biotechnol. 2020, 39, 68–75. (In Chinese) [Google Scholar]
- Zhu, S.; Du, C.; Yu, T.; Cong, X.; Liu, Y.; Chen, S.; Li, Y. Antioxidant Activity of Selenium-Enriched Peptides from the Protein Hydrolysate of Cardamine violifolia. J. Food Sci. 2019, 84, 3504–3511. [Google Scholar] [CrossRef] [PubMed]
- Rhee, K.C. Determination of total nitrogen. In Current Protocols in Food Analytical Chemistry; Wiley: Hoboken, NJ, USA, 2001; pp. B1.2.1–B1.2.9. [Google Scholar]
- Mateo, C.; Fernández-Lorente, G.; Abian, O.; Fernández-Lafuente, R.; Guisán, J.M. Multifunctional epoxy supports: A new tool to improve the covalent immobilization of proteins. The promotion of physical adsorptions of proteins on the supports before their covalent linkage. Biomacromolecules 2001, 1, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Duan, Y.; Cheng, H. Preparation of immobilized pepsin for extraction of collagen from bovine hide. RSC Adv. 2022, 12, 34548–34556. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, S.; Zhang, H.; Wang, F.; Marszałek, K.; Zhu, Z. Multiple effects driven by pulsed electric field to enhance the catalytic efficiency of the mussel-inspired proteolytic membrane in protein hydrolysis. Chem. Eng. J. 2023, 453, 139792. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, J.; Li, S.; Wei, Y.; Wan, Y. Biocatalytic membrane based on polydopamine coating: A platform for studying immobilization mechanisms. Langmuir 2018, 34, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, J.-L.; Xu, H.-J.; Yuan, Z.-H.; Guo, Y. Cellulase deactivation based kinetic modeling of enzymatic hydrolysis of steam-exploded wheat straw. Bioresour. Technol. 2010, 101, 8261–8266. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Cong, X.; Sun, Z.; Chen, Z.; Chen, X.; Zhu, Z.; Li, S.; Cheng, S. Production of Cardamine violifolia selenium-enriched peptide using immobilized Alcalase on Fe3O4 modified by tannic acid and polyethyleneimine. RSC Adv. 2022, 12, 22082–22090. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.-D.; Zeng, H.-Y.; Alain, G.B.F.C.; Feng, B. Heat-pretreatment and enzymolysis behavior of the lotus seed protein. Food Chem. 2016, 201, 230–236. [Google Scholar] [CrossRef] [PubMed]
- AL-Bukhaiti, W.Q.; Al-Dalali, S.; Noman, A.; Qiu, S.; Abed, S.M.; Qiu, S.-X. Response surface modeling and optimization of enzymolysis parameters for antidiabetic activities of peanut protein hydrolysates prepared using two proteases. Foods 2022, 11, 3303–3323. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ruan, G.; Qin, Z.; Li, H.; Zheng, Y. Antioxidant activity measurement and potential antioxidant peptides exploration from hydrolysates of novel continuous microwave-assisted enzymolysis of the Scomberomorus niphonius protein. Food Chem. 2017, 223, 89–95. [Google Scholar] [CrossRef]
- Tan, X.; Qi, L.; Fan, F.; Guo, Z.; Wang, Z.; Song, W.; Du, M. Analysis of volatile compounds and nutritional properties of enzymatic hydrolysate of protein from cod bone. Food Chem. 2018, 264, 350–357. [Google Scholar] [CrossRef]
- Yao, L.W.; Khan, F.S.A.; Mubarak, N.M.; Karri, R.R.; Khalid, M.; Walvekar, R.; Abdullah, E.C.; Mazari, S.A.; Ahmad, A.; Dehghani, M.H. Insight into immobilization efficiency of Lipase enzyme as a biocatalyst on the graphene oxide for adsorption of Azo dyes from industrial wastewater effluent. J. Mol. Liq. 2022, 354, 118849–118863. [Google Scholar] [CrossRef]
- Yang, G.; Wang, J.; Shao, W.; Wang, S.; Wu, B.; Gao, B.; He, B. Mussel-Inspired Electro-oxidation-Modified Three-Dimensional Printed Carriers for a Versatile Enzyme Immobilization Approach. ACS Sustain. Chem. Eng. 2023, 11, 1375–1385. [Google Scholar]
- Wang, H.; Wang, Y.; Sun, X.; Hu, H.; Peng, Q. Two functional post-cross-linked polystyrene resins: Effect of structure on the enhanced removal of benzene sulfonic acid. Colloid Surf. A. 2020, 588, 124398–124405. [Google Scholar] [CrossRef]
- Wu, J.; Han, J.; Mao, Y.; Wang, L.; Wang, Y.; Li, Y.; Wang, Y. Bionic mineralization growth of UIO-66 with bovine serum for facile synthesis of Zr-MOF with adjustable mesopores and its application in enzyme immobilization. Sep. Purif. Technol. 2022, 297, 121505–121515. [Google Scholar]
- Yu, X.; Li, J.; Sun, Y.; Xie, Y.; Su, Y.; Tang, S.; Bian, S.; Liu, L.; Huo, F.; Huang, Q.; et al. Co-immobilized multi-enzyme biocatalytic system on reversible and soluble carrier for saccharification of corn straw cellulose. Bioresour. Technol. 2024, 395, 130325. [Google Scholar]
- Bardestani, R.; Patience, G.S.; Kaliaguine, S. Experimental methods in chemical engineering: Specific surface area and pore size distribution measurements—BET, BJH, and DFT. Can. J. Chem. Eng. 2019, 97, 2781–2791. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, S.; Ma, Y.; Li, Y.; Fei, X.; Xu, L.; Wang, Y.; Tian, J. Synthesis of hydrogel microspheres with tunable pore size and their application in alkaline protease immobilization. New J. Chem. 2024, 48, 2950–2961. [Google Scholar]
- Hosseini, S.S.; Khodaiyan, F.; Mousavi, S.M.E.; Kennedy, J.F.; Azimi, S.Z. A health-friendly strategy for covalent-bonded immobilization of pectinase on the functionalized glass beads. Food Bioprocess Tech. 2020, 14, 177–186. [Google Scholar] [CrossRef]
- Li, J.; Feng, T.; Han, L.; Zhang, M.; Jiang, T. Fabricating biopolymer-inorganic hybrid microspheres for enzyme immobilization: Connect membrane emulsification with biomimetic mineralization. Particuology 2022, 64, 171–177. [Google Scholar] [CrossRef]
- Mazlan, S.Z.; Hanifah, S.A. Effects of temperature and pH on immobilized laccase activity in conjugated methacrylate-acrylate microspheres. Int. J. Polym. Sci. 2017, 2017, 5657271. [Google Scholar] [CrossRef]
- Baltieri, R.C.; Innocentini-Mei, L.H.; Tamashiro, W.M.S.C.; Peres, L.; Bittencourt, E. Synthesis of porous macrospheres from amino-polymers. Eur. Polym. J. 2002, 38, 57–62. [Google Scholar] [CrossRef]
- Guo, J.; Liu, X.; Zhang, X.; Wu, J.; Chai, C.; Ma, D.; Chen, Q.; Xiang, D.; Ge, W. Immobilized lignin peroxidase on Fe3O4@SiO2@polydopamine nanoparticles for degradation of organic pollutants. Int. J. Biol. Macromol. 2019, 138, 433–440. [Google Scholar] [CrossRef]
- Naghdi, M.; Taheran, M.; Brar, S.K.; Kermanshahi-Pour, A.; Verma, M.; Surampalli, R.Y. Immobilized laccase on oxygen functionalized nanobiochars through mineral acids treatment for removal of carbamazepine. Sci. Total Environ. 2017, 584, 393–401. [Google Scholar] [CrossRef]
- Lin, J.; Wen, Q.; Chen, S.; Le, X.; Zhou, X.; Huang, L. Synthesis of amine-functionalized Fe3O4@C nanoparticles for laccase immobilization. Int. J. Biol. Macromol. 2016, 96, 377–383. [Google Scholar] [CrossRef]
- Xia, G.; Huibin, P.; Siyu, T.; Le, S.; Zhanlong, H.; Chengfang, Q.; Qian, L.; Chunsheng, Z. Co-immobilization of bienzyme HRP/GOx on highly stable hierarchically porous MOF with enhanced catalytic activity and stability: Kinetic and thermodynamic studies. J. Environ. Chem. Eng. 2023, 11, 110684–110693. [Google Scholar]
- Karam, E.A.; Abdel Wahab, W.A.; Saleh, S.A.A.; Hassan, M.E.; Kansoh, A.L.; Esawy, M.A. Production, immobilization and thermodynamic studies of free and immobilized Aspergillus awamori amylase. Int. J. Biol. Macromol. 2017, 102, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. R. 1998, 62, 597–635. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, X.; Zhang, X.; Liu, H.; Ao, Q. Amino acid, structure and antioxidant properties of Haematococcus pluvialis protein hydrolysates produced by different proteases. Int. J. Food Sci. Tech. 2021, 56, 185–195. [Google Scholar] [CrossRef]
- Qian, C.; Jiang, Y.; Sun, Y.; Yin, X.; Zhang, M.; Kan, J.; Liu, J.; Xiao, L.; Jin, C.; Qi, X.; et al. Changes in the texture and flavor of lotus root after different cooking methods. Foods 2023, 12, 2012–2028. [Google Scholar] [CrossRef]
- Yang, A.; Long, C.; Xia, J.; Tong, P.; Cheng, Y.; Wang, Y.; Chen, H. Enzymatic characterisation of the immobilised Alcalase to hydrolyse egg white protein for potential allergenicity reduction. J. Sci. Food. Agr. 2017, 97, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Žuža, M.G.; Milašinović, N.Z.; Jonović, M.M.; Jovanović, J.R.; Kalagasidis Krušić, M.T.; Bugarski, B.M.; Knežević-Jugović, Z.D. Design and characterization of alcalase-chitosan conjugates as potential biocatalysts. Bioproc. Biosyst. Eng. 2017, 40, 1713–1723. [Google Scholar] [CrossRef]
- Jonović, M.; Žuža, M.; Đorđević, V.; Šekuljica, N.; Milivojević, M.; Jugović, B.; Bugarski, B.; Knežević-Jugović, Z. Immobilized Alcalase on micron- and submicron-sized alginate beads as a potential biocatalyst for hydrolysis of food proteins. Catalysts 2021, 11, 305–322. [Google Scholar] [CrossRef]
- Mao, Y.; Chen, L.; Zhang, L.; Bian, Y.; Meng, C. Synergistic hydrolysis of soy proteins using immobilized proteases: Assessing peptide profiles. Foods 2023, 12, 4115. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | SBET/m2·g−1 | DBJH/nm | Vtotal/cm3·g−1 |
|---|---|---|---|
| Resin | 113.678 | 13.981 | 0.442 |
| Activated resin | 50.261 | 10.970 | 0.154 |
| Co-immobilized enzyme11:1 | 75.923 | 13.072 | 0.260 |
| Co-immobilized enzyme9:1 | 81.112 | 8.701 | 0.184 |
| Co-immobilized enzyme7:1 | 77.231 | 14.217 | 0.301 |
| Co-immobilized enzyme5:1 | 81.201 | 7.065 | 0.151 |
| Co-immobilized enzyme3:1 | 89.214 | 12.757 | 0.296 |
| Co-immobilized enzyme1:1 | 93.074 | 13.152 | 0.328 |
| Amino Acid | 11:1 (mg/L) | 9:1 (mg/L) | 7:1 (mg/L) | 5:1 (mg/L) | 3:1 (mg/L) | 1:1 (mg/L) |
|---|---|---|---|---|---|---|
| Asp | 0.041 ± 0.005 | 0.040 ± 0.006 | 0.038 ± 0.007 | 1.126 ± 0.019 | 0.037 ± 0.006 | 0.125 ± 0.004 |
| Glu | 1.058 ± 0.007 | 1.11 ± 0.012 | 0.454 ± 0.014 | 0.058 ± 0.010 | 0.737 ± 0.012 | 1.288 ± 0.012 |
| Ser | 2.360 ± 0.006 | 2.574 ± 0.014 | 2.452 ± 0.016 | 2.182 ± 0.017 | 2.331 ± 0.013 | 0.160 ± 0.008 |
| Gly | 1.211 ± 0.008 | 1.277 ± 0.005 | 1.296 ± 0.017 | 1.131 ± 0.010 | 1.177 ± 0.017 | 0.987 ± 0.015 |
| Thr | 0.560 ± 0.014 | 0.602 ± 0.012 | 0.609 ± 0.015 | 0.485 ± 0.019 | 0.559 ± 0.012 | 0.445 ± 0.013 |
| Pro | 0.566 ± 0.016 | 0.604 ± 0.015 | 0.610 ± 0.011 | 0.507 ± 0.007 | 0.560 ± 0.015 | 0.504 ± 0.014 |
| Ala | 1.187 ± 0.012 | 1.286 ± 0.013 | 1.283 ± 0.012 | 1.085 ± 0.016 | 1.182 ± 0.012 | 0.792 ± 0.018 |
| Val | 1.01 ± 0.014 | 1.059 ± 0.017 | 1.054 ± 0.013 | 0.898 ± 0.018 | 0.959 ± 0.011 | 0.815 ± 0.013 |
| Met | 0.090 ± 0.005 | 0.108 ± 0.009 | 0.123 ± 0.007 | 0.117 ± 0.012 | 0.103 ± 0.007 | 0.065 ± 0.008 |
| Ile | 1.254 ± 0.015 | 1.346 ± 0.014 | 1.346 ± 0.011 | 1.133 ± 0.015 | 1.237 ± 0.012 | 1.01 ± 0.012 |
| Leu | 0.584 ± 0.014 | 0.608 ± 0.010 | 0.589 ± 0.016 | 0.552 ± 0.005 | 0.554 ± 0.015 | 0.438 ± 0.011 |
| Trp | 0.531 ± 0.017 | 0.537 ± 0.013 | 0.555 ± 0.007 | 0.529 ± 0.014 | 0.559 ± 0.012 | 0.191 ± 0.015 |
| Phe | 0.020 ± 0.007 | 0.022 ± 0.006 | 0.012 ± 0.003 | 0.032 ± 0.002 | 0.017 ± 0.006 | |
| His | 0.024 ± 0.006 | 0.021 ± 0.006 | 0.016 ± 0.003 | 0.034 ± 0.002 | 0.018 ± 0.006 | |
| Lys | 0.348 ± 0.005 | 0.315 ± 0.012 | 0.390 ± 0.013 | 0.436 ± 0.012 | 0.389 ± 0.014 | 0.129 ± 0.006 |
| Tyr | 0.005 ± 0.002 | 0.028 ± 0.005 | 0.013 ± 0.003 | |||
| Arg | 0.415 ± 0.005 | 0.452 ± 0.014 | 0.490 ± 0.013 | 0.399 ± 0.010 | 0.437 ± 0.010 | 0.449 ± 0.011 |
| Cys | 0.607 ± 0.015 | 0.637 ± 0.012 | 0.638 ± 0.012 | 0.552 ± 0.017 | 0.577 ± 0.016 | 0.509 ± 0.010 |
| Sample | Total Se | Inorganic Se | Organic Se |
|---|---|---|---|
| CV (mg/kg) | 1980.48 ± 50.11 | 162.77 ± 5.05 | 1817.70 ± 50.21 |
| CV protein (mg/kg) | 1034.45 ± 0.07 | 53.25 ± 3.20 | 981.19 ± 3.22 |
| 11:1 (mg/L) | 10.53 ± 0.21 | 0.78 ± 0.003 | 9.75 ± 0.21 |
| 9:1 (mg/L) | 11.21 ± 0.07 | 0.81 ± 0.03 | 10.39 ± 0.05 |
| 7:1 (mg/L) | 12.91 ± 0.41 | 0.97 ± 0.10 | 11.94 ± 0.49 |
| 5:1 (mg/L) | 12.80 ± 0.11 | 0.83 ± 0.04 | 11.98 ± 0.13 |
| 3:1 (mg/L) | 10.75 ± 0.11 | 0.93 ± 0.05 | 9.82 ± 0.13 |
| 1:1 (mg/L) | 11.17 ± 0.20 | 0.71 ± 0.06 | 10.45 ± 0.18 |
| Retention Time | MS (m/z) | Sequence | Molecular Weight (Da) |
|---|---|---|---|
| 1.8075 | 597.25 | SSSSTSKVFLC | 1192.4879 |
| 18.571 | 724.36 | ILELLLTTYAFC | 1446.6913 |
| 24.451 | 718.37 | EITVLCDAKVALI | 1434.7237 |
| 38.923 | 753.87 | CVIASTI | 753.3176 |
| 40.994 | 755.38 | LVNRKITALCNES | 1507.7261 |
| 43.816 | 528.95 | LTLEDPTATLEAFLCDKDA | 2112.937 |
| 44.59 | 696.87 | VLVNRKITALCN | 1390.7199 |
| 45.586 | 754.87 | VETQLQLFIGLPC | 1507.7189 |
| 47.201 | 604.31 | SLALCLLSLGGL | 1206.6127 |
| 48.606 | 905.92 | PKCVSDPL | 905.3761 |
| 54.373 | 548.27 | LGFVLVCIAL | 1094.5643 |
| 56.333 | 623.32 | PILVHCKTSAK | 1243.6192 |
| Carrier | Enzyme | Enzyme Loading (mg/g) | Enzyme Active (U/mg) | Loading Rate (%) | Reaction Time (min) | DH (%) | Reference |
|---|---|---|---|---|---|---|---|
| Resin | Alcalase | 405.6 | 6.7 | 45.1 | 120 | 25.8 | This study |
| Resin | Alcalase and Dispase | 497.8 | 41 | 77.7 | 120 | 27.2 | This study |
| Magnetic beads | Alcalase | 925 | 20.55 | 45 | 180 | 20 | [59] |
| Chitosan | Alcalase | 340.2 | 23.6 | 96.4 | 180 | 29.9 | [60] |
| Alginate beads | Alcalase | 671.6 | 2.7 | 98.7 | 225 | ˂25 | [61] |
| SiO2 | Alcalase and Flavorzyme | 25 | 180 | ˂ 80 | 1080 | 5.9 | [62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Li, Y.; Chen, X.; Zhu, Z.; Li, S.; Song, J.; Zheng, Z.; Cong, X.; Cheng, S. Co-Immobilization of Alcalase/Dispase for Production of Selenium-Enriched Peptide from Cardamine violifolia. Foods 2024, 13, 1753. https://doi.org/10.3390/foods13111753
Zhu S, Li Y, Chen X, Zhu Z, Li S, Song J, Zheng Z, Cong X, Cheng S. Co-Immobilization of Alcalase/Dispase for Production of Selenium-Enriched Peptide from Cardamine violifolia. Foods. 2024; 13(11):1753. https://doi.org/10.3390/foods13111753
Chicago/Turabian StyleZhu, Shiyu, Yuheng Li, Xu Chen, Zhenzhou Zhu, Shuyi Li, Jingxin Song, Zhiqiang Zheng, Xin Cong, and Shuiyuan Cheng. 2024. "Co-Immobilization of Alcalase/Dispase for Production of Selenium-Enriched Peptide from Cardamine violifolia" Foods 13, no. 11: 1753. https://doi.org/10.3390/foods13111753
APA StyleZhu, S., Li, Y., Chen, X., Zhu, Z., Li, S., Song, J., Zheng, Z., Cong, X., & Cheng, S. (2024). Co-Immobilization of Alcalase/Dispase for Production of Selenium-Enriched Peptide from Cardamine violifolia. Foods, 13(11), 1753. https://doi.org/10.3390/foods13111753