
Comprehensive Insights into Ochratoxin A: Occurrence, Analysis, and Control Strategies
 , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Ochratoxin A (OTA): Chemical Structure, Fungal Producers, and Conditions for Formation in Commodities
3. OTA Biosynthetic Pathway and Regulatory Mechanisms
4. Ochratoxin A Occurrence in Food
5. OTA Analytical Methods
6. Strategies for OTA Control
6.1. Chemical Methods
6.1.1. Synthetic Fungicides
6.1.2. Chemical Degradation of OTA
6.2. Physical Methods
6.3. Adsorption by Physical–Chemical Agents
6.4. Biocontrol Strategies for OTA Control
6.4.1. Microorganisms and Enzymes
6.4.2. Plant-Derived Extracts for OTA Biocontrol
7. Prospects and Application
- (a)
- Continuous improvements in analytical techniques, such as mass spectrometry and molecular approaches, will make it possible to identify low levels of OTA in food items more quickly.
- (b)
- The development of tests based on nanotechnology and biosensors for OTA detection can offer extremely sensitive and selective detection techniques, obviating the requirement for sophisticated laboratory apparatus and knowledgeable staff.
- (c)
- Predictive modelling, risk assessment, and early warning systems can be improved by incorporating machine learning and artificial intelligence algorithms into OTA analysis. These technologies can assist in locating high-risk areas and products, enabling focused monitoring and control efforts.
- (d)
- Researchers should explore diverse avenues to combat OTA contamination. Bioengineered microorganisms or molecular modified enzymes, with optimized OTA-degrading capabilities and stability, offer promise. In fact, it is desirable to obtain new, genetically engineered enzymes with higher pH stability, active over a wider temperature range, stable over time, and characterized by high expression/high yield in microbial culture; this could be a relevant step forward. Incorporating such microorganisms, especially enzymes, into food and beverage fermentation processes could reduce contamination, especially in items like wine, beer, and dairy products.
- (e)
- Crop susceptibility to OTA-producing molds can be decreased by developing best practices for crop management, including breeding programs for OTA-resistant crop varieties. Additionally, crop cultivation may be optimized and fungal contamination minimized with the aid of precision agriculture and weather forecasts.
- (f)
- OTA contamination during food production and distribution can be reduced due to advancements in food processing and storage technologies. Products shelf lives can be increased while OTA levels are decreased using better packing materials, altered atmospheres, and irradiation procedures.
- (g)
- Increased international collaboration and unified rules are also necessary. International standards, stricter laws, and the introduction of monitoring systems to verify compliance across borders are all possibilities for the future.
- (h)
- Demand for safer products may rise with increasing customer knowledge of the dangers of OTA-contaminated food. Future applications might make use of QR codes and mobile apps to inform customers about the OTA (low) content of the goods they buy.
- (i)
- Enhancing food safety measures in emerging markets might contribute significantly to both OTA contamination and control efforts as the world food trade expands.
- (j)
- Individuals may have the opportunity to assess their susceptibility to mycotoxin exposure through advancements in personalized nutrition and healthcare. This information could inform dietary choices and lead to customized recommendations for minimizing OTA intake.
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- González-Curbelo, M.Á.; Kabak, B. Occurrence of Mycotoxins in Dried Fruits Worldwide, with a Focus on Aflatoxins and Ochratoxin A: A Review. Toxins 2023, 15, 576. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, K.; Nabi, B.G.; Ansar, S.; Bhat, Z.F.; Aadil, R.M.; Mousavi Khaneghah, A. Mycotoxins and consumers’ awareness: Recent progress and future challenges. Toxicon 2023, 232, 107227. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.A.; Strub, C.; Fontana, A.; Schorr-Galindo, S. Crop molds and mycotoxins: Alternative management using biocontrol. Biol. Control 2017, 104, 10–27. [Google Scholar] [CrossRef]
- Ukwuru, M.U.; Ohaegbu, C.G.; Muritala, A. An overview of mycotoxin contamination of foods and feeds. J. Biochem. Microb. Toxicol. 2017, 1, 101. [Google Scholar] [CrossRef]
- Ben Miri, Y.; Ariño, A.; Djenane, D. Study of antifungal, anti-aflatoxigenic, antioxidant activity and phytotoxicity of Algerian Citrus limon var. Eureka and Citrus sinensis var. Valencia essential oils. J. Essent. Oil Bear. Plants 2018, 21, 345–361. [Google Scholar] [CrossRef]
- Li, X.; Ma, W.; Ma, Z.; Zhang, Q.; Li, H. The occurrence and contamination level of ochratoxin A in plant and animal-derived food commodities. Molecules 2021, 26, 6928. [Google Scholar] [CrossRef]
- Shukla, R.; Singh, P.; Prakash, B.; Dubey, N.K. Antifungal, aflatoxin inhibition and antioxidant activity of Callistemon lanceolatus (Sm.) Sweet essential oil and its major component 1, 8-cineole against fungal isolates from chickpea seeds. Food Control 2012, 25, 27–33. [Google Scholar] [CrossRef]
- Pallarés, N.; Berrada, H.; Tolosa, J.; Ferrer, E. Effect of high hydrostatic pressure (HPP) and pulsed electric field (PEF) technologies on reduction of aflatoxins in fruit juices. LWT Food Sci. Technol. 2021, 142, 3769. [Google Scholar] [CrossRef]
- Rhee, K.H.; Yang, S.A.; Pyo, M.C.; Lim, J.-M.; Lee, K.-W. MiR-155-5p Elevated by Ochratoxin A Induces Intestinal Fibrosis and Epithelial-to-Mesenchymal Transition through TGF-β Regulated Signaling Pathway In Vitro and In Vivo. Toxins 2023, 15, 473. [Google Scholar] [CrossRef]
- Su, B.; Zhang, Z.; Sun, Z.; Tang, Z.; Xie, X.; Chen, Q.; Hammock, B.D. Fluonanobody-based nanosensor via fluorescence resonance energy transfer for ultrasensitive detection of ochratoxin A. J. Hazard. Mater. 2022, 422, 126838. [Google Scholar] [CrossRef]
- Kaniou-Grigoriadou, I.; Eleftheriadou, A.; Mouratidou, T.; Katikou, P. Determination of aflatoxin M1 in ew’’s milk samples and the produced curd and Feta cheese. Food Control 2005, 16, 257–261. [Google Scholar] [CrossRef]
- Hymery, N.; Sibiril, Y.; Parent-Massin, D. In vitro effects of trichothecenes on human dendritic cells. Toxicol. Vitr. 2006, 20, 899–909. [Google Scholar] [CrossRef]
- Oguz, H. Mikotoksinler ve Önemi (Mycotoxins and their importance). Turk. Klin. J. Vet. Sci. Pharmacol. Toxicol. Spec. Top. 2017, 3, 113–119. [Google Scholar]
- El-Sayed, R.A.; Jebur, A.B.; Kang, W.; El-Demerdash, F.M. An overview on the major mycotoxins in food products: Characteristics, toxicity, and analysis. J. Fut. Foods 2022, 2, 91–102. [Google Scholar] [CrossRef]
- Nazhand, A.; Durazzo, A.; Lucarini, M.; Souto, E.B.; Santini, A. Characteristics, occurrence, detection and detoxification of aflatoxins in foods and feeds. Foods 2020, 9, 644. [Google Scholar] [CrossRef]
- Pickova, D.; Ostry, V.; Malir, F. A recent overview of producers and important dietary sources of aflatoxins. Toxins 2021, 13, 186. [Google Scholar] [CrossRef]
- Kamle, M.; Mahato, D.K.; Devi, S.; Lee, E.K.; Kang, S.G.; Pradeep, K. Fumonisins: Impact on agriculture, food, and human health and their management strategies. Toxins 2019, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Benthem de Grave, X.; Saltzmann, J.; Laurain, J.; Rodriguez, M.A.; Molist, F.; Danicke, S.; Santos, R.R. Transmission of zearalenone, deoxynivalenol, and their derivatives from sows to piglets during lactation. Toxins 2021, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, C.; Mao, Y.; Hong, X.; Du, D. Zearalenone contamination in corn, corn products, and swine feed in China in 2016–2018 as assessed by magnetic bead immunoassay. Toxins 2019, 11, 451. [Google Scholar] [CrossRef]
- Sajid, M.; Mehmood, S.; Yuan, Y.; Yue, T. Mycotoxin patulin in food matrices: Occurrence and its biological degradation strategies. Drug Metab. Rev. 2019, 51, 105–120. [Google Scholar] [CrossRef]
- Wei, C.; Yu, L.; Qiao, N.; Zhao, J.; Zhang, H.; Zhai, Q.; Tian, F.; Chen, W. Progress in the distribution, toxicity, control, and detoxification of patulin: A review. Toxicon 2020, 184, 83–93. [Google Scholar] [CrossRef]
- Altafini, A.; Roncada, P.; Guerrini, A.; Sonfack, G.M.; Fedrizzi, G.; Caprai, E. Occurrence of ochratoxin A in different types of cheese offered for sale in Italy. Toxins 2021, 13, 540. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Villeda, B.; Lobos, O.; Aguilar-Zuniga, K.; Carrasco-Sánchez, V. Ochratoxins in wines: A review of their occurrence in the last decade, toxicity, and exposure risk in humans. Toxins 2021, 13, 478. [Google Scholar] [CrossRef] [PubMed]
- Ben Miri, Y.; Benabdallah, A.; Taoudiat, A.; Mahdid, M.; Djenane, D.; Tacer-Caba, Z.; Topkaya, C.; Simal-Gandara, J. Potential of essential oils for protection of Couscous against Aspergillus flavus and aflatoxin B1 contamination. Food Control 2023, 145, 109474. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 13748. [Google Scholar] [CrossRef]
- Topi, D.; Babič, J.; Jakovac-Strajn, B.; Tavčar-Kalcher, G. Incidence of aflatoxins and ochratoxin A in wheat and corn from Albania. Toxins 2023, 15, 567. [Google Scholar] [CrossRef] [PubMed]
- Mavioğlu Kaya, M.; Deveci, H.A.; Kaya, İ.; Atar, N.; Yola, M.L. The Electrochemical Detection of Ochratoxin A in Apple Juice via MnCO3 Nanostructures Incorporated into Carbon Fibers Containing a Molecularly Imprinting Polymer. Biosensors 2023, 13, 760. [Google Scholar] [CrossRef]
- Hassan, H.F.; Tashani, H.; Ballouk, F.; Daou, R.; El Khoury, A.; Abiad, M.G.; AlKhatib, A.; Hassan, M.; El Khatib, S.; Dimassi, H. Aflatoxins and Ochratoxin A in Tea Sold in Lebanon: Effects of Type, Packaging, and Origin. Int. J. Environ. Res. Public Health 2023, 20, 6556. [Google Scholar] [CrossRef]
- Xu, A.; Zhou, H.; Yu, S.; Li, Y.; Wang, L.; Wu, A.; Liang, J.; Peng, S.; Liu, N. Fusarium Mycotoxins and OTA in Beer from Shanghai, the Largest Megacity in China: Occurrence and Dietary Risk Assessment. Foods 2023, 12, 3071. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-H.; Huang, C.H.; Liu, T.Y.; Liou, J.S.; Hou, C.Y.; Hsu, W.H. Microbial Diversity of Anaerobic-Fermented Coffee and Potential for Inhibiting Ochratoxin-Produced Aspergillus niger. Foods 2023, 12, 2967. [Google Scholar] [CrossRef]
- EFSA CONTAM Panel (EFSA Panel on Contaminants in the Food Chain); Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.-C.; Nebbia, C.S.; et al. Scientific Opinion on the risk assessment of ochratoxin A in food. EFSA J. 2020, 18, 6113. [Google Scholar] [CrossRef]
- Commission Regulation (EU) 2023/915 of 25 April 2023 on Maximum Levels for Certain Contaminants in Food and Repealing Regulation (EC) No 1881/2006, Consolidated Text. Available online: http://data.europa.eu/eli/reg/2023/915/2023-08-10 (accessed on 25 March 2024).
- Alshannaq, A.; Yu, J.H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed]
- Smaoui, S.; D’Amore, T.; Agriopoulou, S.; Mousavi Khaneghah, A. Mycotoxins in Seafood: Occurrence, Recent Development of Analytical Techniques and Future Challenges. Separations 2023, 10, 217. [Google Scholar] [CrossRef]
- Han, X.; Chakrabortti, A.; Zhu, J.; Liang, Z.X.; Li, J. Sequencing and functional annotation of the whole genome of the filamentous fungus Aspergillus westerdijkiae. BMC Genom. 2016, 17, 633. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Liu, F.; Wang, Q.; Selvaraj, J.N.; Xing, F.; Liu, Y. Ochratoxin A producing fungi, biosynthetic pathway and regulatory mechanisms. Toxins 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, Drugs and Other Extrolites Produced by Species in Penicillium Subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Abarca, M.L.; Accensi, F.; Cano, J.; Cabañes, F.J. Taxonomy and significance of black Aspergilli. Antonie Leeuwenhoek 2004, 86, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Noonim, P.; Meijer, M.; Houbraken, J.A.M.P.; Frisvad, J.C.; Varga, J. Diagnostic tools to identify black Aspergilli. Stud. Mycol. 2007, 59, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Bayman, P.; Baker, J.L.; Doster, M.A.; Michailides, T.J.; Mahoney, N.E. Ochratoxin production by the Aspergillus ochraceus group and Aspergillus alliaceus. Appl. Environ. Microbiol. 2002, 68, 2326–2329. [Google Scholar] [CrossRef] [PubMed]
- Baranyi, N.; Jaksić, D.D.; Palágyi, A.; Kiss, N.; Kocsubé, S.; Szekeres, A.; Kecskeméti, A.; Bencsik, O.; Vágvölgyi, C.; Segvić, K.M.; et al. Identification of Aspergillus Species in Central Europe Able to Produce G-Type Aflatoxins. Acta Biol. Hung. 2015, 66, 339–347. [Google Scholar] [CrossRef]
- Camiletti, B.X.; Torrico, A.K.; Maurino, M.F.; Cristos, D.; Magnoli, C.; Lucini, E.I.; Pecci, M.D.L.P.G. Fungal screening and aflatoxin production by Aspergillus section Flavi isolated from pre-harvest maize ears grown in two Argentine regions. Crop. Prot. 2017, 92, 41–48. [Google Scholar] [CrossRef]
- Saldan, N.C.; Almeida, R.T.; Avíncola, A.; Porto, C.; Galuch, M.B.; Magon, T.F.; Oliveira, C.C. Development of an analytical method for identification of Aspergillus flavus based on chemical markers using HPLC-MS. Food Chem. 2018, 241, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Houbraken, J. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Frenette, C.; Paugh, R.J.; Tozlovanu, M.; Juzio, M.; Pfohl-Leszkowicz, A.; Manderville, R.A. Structure-activity relationships for the fluorescence of ochratoxin A: Insight for detection of ochratoxin A metabolites. Anal. Chim. Acta 2008, 617, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Kiseleva, M.; Chalyy, Z.; Sedova, I.; Aksenov, I. Stability of Mycotoxins in Individual Stock and Multi-Analyte Standard Solutions. Toxins 2020, 12, 94. [Google Scholar] [CrossRef] [PubMed]
- Taniwaki, M.H.; Urbano, G.R.; Cabrera Palacios, H.A.; LeitaÄo, M.F.F.; Menezes, H.C.; Vicentini, M.C.; Iamanaka, B.T.; Taniwaki, N.N. Infuence of water activity on mould growth and ochratoxin A production in coffee. In Proceedings of the 19th ASIC Coffee Conference, Trieste, Italy, 14–18 May 2001. [Google Scholar]
- Ramos, A.; Labernia, N.; Marín, S.; Sanchis, V.; Magan, N. Effect of water activity and temperature on growth and ochratoxin production by three strains of Aspergillus ochraceus on a barley extract medium and on barley grains. Int. J. Food Microbiol. 1998, 44, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Bellí, N.; Mitchell, D.; Marín, S.; Alegre, I. Ochratoxin A-producing fungi in Spanish wine grapes and their relationship with meteorological conditions. J. Plant Pathol. 2005, 113, 233–239. [Google Scholar] [CrossRef]
- Esteban, A.; Abarca, M.; Bragulat, R.; Cabañes, F. Effect of water activity on ochratoxin A production by Aspergillus niger aggregate species. Int. J. Food Microbiol. 2006, 108, 188–195. [Google Scholar] [CrossRef]
- Lee, H.; Magan, N. Impact of environment and interspecific interactions between spoilage fungi and Aspergillus ochraceus on growth and Ochratoxin production in maize grain. Int. J. Food Microbiol. 2000, 61, 11–16. [Google Scholar] [CrossRef]
- Kuruc, J.; Hegstad, H.; Lee, K.; Wolf-Hall, C. Infestation and Quantification of Ochratoxigenic Fungi in Barley and Wheat Naturally Contaminated with Ochratoxin, A. J. Food Prot. 2015, 78, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; Aldred, D. Conditions of formation of Ochratoxin A in drying, transport and in different commodities. Food Addit. Contam. 2005, 1, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Czaban, J.; Wróblewska, B.; Stochmal, A.; Janda, B. Growth of Penicillium verrucosum and production of Ochratoxin A on nonsterilized wheat grain incubated at different temperatures and water content. Pol. J. Microbiol. 2006, 55, 321–331. [Google Scholar] [PubMed]
- Cairns, V.; Aldred, D.; Magan, N. Water temperature and gas composition interactions affect growth and Ochratoxin A production by isolates of Penicillium verrucosum on wheat grain. J. Appl. Microbiol. 2005, 99, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Kapetanakou, A.E.; Panagou, E.Z.; Gialitaki, M.; Drosinos, E.H.; Skandamis, P.N. Evaluating the combined effect of water activity, pH and temperature on ochratoxin A production by Aspergillus ochraceus and Aspergillus carbonarius on culture medium and Corinth raisins. Food Control 2009, 20, 725–732. [Google Scholar] [CrossRef]
- Koteswara Rao, V.; Ramana, M.V.; Girisham, S.; Reddy, S.M. Culture Media and Factors Influencing Ochratoxin A Production by Two Species of Penicillium Isolated from Poultry Feeds. Natl. Acad. Sci. Lett. 2013, 36, 101–110. [Google Scholar] [CrossRef]
- Passamani, F.R.; Hernandes, T.; Lopes, N.A.; Bastos, S.C.; Santiago, W.D.; Cardoso, M.D.; Batista, L.R. Effect of temperature, water activity, and pH on growth and production of ochratoxin A by Aspergillus niger and Aspergillus carbonarius from Brazilian grapes. J. Food Prot. 2014, 77, 1947–1952. [Google Scholar] [CrossRef]
- Huffman, J.; Gerber, R.; Du, L. Recent advancements in the biosynthetic mechanisms for polyketide-derived mycotoxins. Biopolymers 2010, 93, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.; Perrone, G.; Gambacorta, L.; Epifani, F.; Solfrizzo, M.; Gallo, A. Identification of a Halogenase Involved in the Biosynthesis of Ochratoxin A in Aspergillus carbonarius. Appl. Environ. Microbiol. 2016, 82, 5631–5641. [Google Scholar] [CrossRef]
- Gallo, A.; Ferrara, M.; Perrone, G. Phylogenetic study of polyketide synthases and nonribosomal peptide synthetases involved in the biosynthesis of mycotoxins. Toxins 2013, 5, 717–742. [Google Scholar] [CrossRef]
- Schwarzer, D.; Marahiel, M.A. Multimodular Biocatalysts for Natural Product Assembly. Naturwissenschaften 2001, 88, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Amoutzias, G.D.; Van de Peer, Y.; Mossialos, D. Evolution and taxonomic distribution of nonribosomal peptide and polyketide synthases. Future Microbiol. 2008, 3, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Färber, P.; Geisen, R. Analysis of differentially-expressed ochratoxin A biosynthesis genes of Penicillium nordicum. Eur. J. Plant Pathol. 2004, 110, 661–669. [Google Scholar] [CrossRef]
- Karolewiez, A.; Geisen, R. Cloning a part of the ochratoxin A biosynthetic gene cluster of Penicillium nordicum and characterization of the ochratoxin polyketide synthase gene. Syst. Appl. Microbiol. 2005, 28, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.B.; et al. A Consensus Ochratoxin A Biosynthetic Pathway: Insights from the Genome Sequence of Aspergillus ochraceus and a Comparative Genomic Analysis. Appl. Environ. Microbiol. 2018, 84, e01009-18. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya-Bayram, Ö.; Palmer, J.M.; Keller, N.; Braus, G.H.; Bayram, Ö. One Juliet and four Romeos: VeA and its methyltransferases. Front. Microbiol. 2015, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Feussner, K.; Dumkow, M.; Herrfurth, C.; Feussner, I.; Braus, G.H. Changes of global gene expression and secondary metabolite accumulation during light-dependent Aspergillus nidulans development. Fungal Genet. Biol. 2016, 87, 30–53. [Google Scholar] [CrossRef]
- Reverberi, M.; Gazzetti, K.; Punelli, F.; Scarpari, M.; Zjalic, S.; Ricelli, A.; Fanelli, C. Ao yap1 regulates OTA synthesis by controlling cell redox balance in Aspergillus ochraceus. Appl. Microbiol. Biotechnol. 2012, 95, 1293–1304. [Google Scholar] [CrossRef]
- Hashem, A.; Abd-Allah, E.F.; Al-Obeed, R.S.; Alqarawi, A.A.; Alwathnani, H.A. Effect of carbon, nitrogen sources and water activity on growth and ochratoxin production of Aspergillus carbonarius (Bainier) Thom. Jundishapur J. Microbiol. 2015, 8, e17569. [Google Scholar] [CrossRef]
- Rispail, N.; Soanes, D.M.; Ant, C.; Czajkowski, R.; Grünler, A.; Huguet, R.; Di Pietro, A. Comparative genomics of MAP kinase and calcium–calcineurin signalling components in plant and human pathogenic fungi. Fungal Genet. Biol. 2009, 46, 287–298. [Google Scholar] [CrossRef]
- Stoll, D.; Schmidt-Heydt, M.; Geisen, R. Differences in the regulation of ochratoxin A by the HOG pathway in Penicillium and Aspergillus in response to high osmolar environments. Toxins 2013, 5, 1282–1298. [Google Scholar] [CrossRef]
- Majeed, M.; Khaneghah, A.M.; Kadmi, Y.; Khan, M.U.; Shariati, M.A. Assessment of ochratoxin A in commercial corn and wheat products. Curr. Nutr. Food Sci. 2018, 14, 116–120. [Google Scholar] [CrossRef]
- Nazari, F.; Sulyok, M.; Yazdanpanah, H.; Kobarfard, F.; Krska, R. A survey of mycotoxins in domestic rice in Iran by liquid chromatography tandem mass spectrometry. Toxicol. Mech. Methods 2014, 24, 37–41. [Google Scholar] [CrossRef]
- Kuruc, J.A.; Schwarz, P.; Wolf-Hall, C. Ochratoxin A in stored US barley and wheat. J. Food Prot. 2015, 78, 597–601. [Google Scholar] [CrossRef]
- Lee, H.J.; Ryu, D. Significance of ochratoxin A in breakfast cereals from the United States. J. Agric. Food Chem. 2015, 63, 9404–9409. [Google Scholar] [CrossRef] [PubMed]
- Limay-Rios, V.; Miller, J.D.; Schaafsma, A.W. Occurrence of Penicillium verrucosum, ochratoxin A, ochratoxin B and citrinin in on-farm stored winter wheat from the Canadian Great Lakes Region. PLoS ONE 2017, 12, e0181239. [Google Scholar] [CrossRef] [PubMed]
- Zebiri, S.; Mokrane, S.; Verheecke-Vaessen, C.; Choque, E.; Reghioui, H.; Sabaou, N.; Riba, A. Occurrence of ochratoxin A in Algerian wheat and its milling derivatives. Toxin Rev. 2019, 38, 206–211. [Google Scholar] [CrossRef]
- Belasli, A.; Herrera, M.; Ariño, A.; Djenane, D. Occurrence and Exposure Assessment of Major Mycotoxins in Foodstuffs from Algeria. Toxins 2023, 15, 449. [Google Scholar] [CrossRef]
- Kosicki, R.; Twarużek, M.; Dopierała, P.; Rudzki, B.; Grajewski, J. Occurrence of mycotoxins in winter rye varieties cultivated in Poland (2017–2019). Toxins 2020, 12, 423. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.J.G.; Teixeira, A.C.; Pereira, A.M.P.T.; Pena, A.; Lino, C.M. Ochratoxin A in Beers Marketed in Portugal: Occurrence and Human Risk Assessment. Toxins 2020, 12, 249. [Google Scholar] [CrossRef] [PubMed]
- Jonatova, P.; Dzuman, Z.; Prusova, N.; Hajslova, J.; Stranska-Zachariasova, M. Occurrence of ochratoxin A and its stereoisomeric degradation product in various types of coffee available in the Czech market. World Mycotoxin J. 2020, 13, 97–107. [Google Scholar] [CrossRef]
- Vanesa, D.; Ana, P. Occurrence of Ochratoxin A in coffee beans, ground roasted coffee and soluble coffee and method validation. Food Control 2013, 30, 675–678. [Google Scholar] [CrossRef]
- Galarce-Bustos, O.; Alvarado, M.; Vega, M.; Aranda, M. Occurrence of ochratoxin A in roasted and instant coffees in Chilean market. Food Control 2014, 46, 102–107. [Google Scholar] [CrossRef]
- Coronel, M.B.; Marin, S.; Cano, G.; Ramos, A.J.; Sanchis, V. Ochratoxin A in Spanish retail ground roasted coffee: Occurrence and assessment of the exposure in Catalonia. Food Control 2011, 22, 414–419. [Google Scholar] [CrossRef]
- Pires, P.N.; Vargas, E.A.; Gomes, M.B.; Vieira, C.B.M.; Santos, E.A.D.; Bicalho, A.A.C.; Trovatti Uetanabaro, A.P. Aflatoxins and ochratoxin A: Occurrence and contamination levels in cocoa beans from Brazil. Food Addit. Contam. Part A 2019, 36, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, A.M.; Scott, P.M. Ochratoxin A in cocoa and chocolate sampled in Canada. Food Addit. Contam. Part A 2011, 28, 762–766. [Google Scholar] [CrossRef] [PubMed]
- Celik, D.; Kabak, B. Assessment to propose a maximum permitted level for ochratoxin A in dried figs. J. Food Compos. Anal. 2022, 112, 104705. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Marini, A.; Tosi, L. A review on the occurrence and control of ochratoxigenic fungal species and ochratoxin A in dehydrated grapes, non-fortified dessert wines and dried vine fruit in the Mediterranean area. Food Control 2012, 26, 347–356. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Krska, R.; Sulyok, M. Occurrence of ochratoxins, fumonisin B2, aflatoxins (B1 and B2), and other secondary fungal metabolites in dried date palm fruits from Egypt: A mini-survey. J. Food Sci 2018, 83, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Nikolchina, I.; Rodrigues, P. A preliminary study on microbiota and ochratoxin a contamination in commercial palm dates (Phoenix dactylifera). Mycotoxin Res. 2021, 37, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.D.; O’Keeffe, T.L.; Vasquez, S.J.; Mahoney, N.E. Isolation and identification of ochratoxin A producing Aspergillus section Nigri strains from California raisins. Lett. Appl. Microbiol. 2011, 52, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.Z.; Mumtaz, A.; Mahmood, Z.; Waqas, M.; Ghaffar, A.; Ismail, A.; Pervaiz, W. Assessment of aflatoxins and ochratoxin A in Chili sauce samples and estimation of dietary intake. Food Control 2021, 121, 107621. [Google Scholar] [CrossRef]
- Prelle, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Co-occurrence of aflatoxins and ochratoxin A in spices commercialized in Italy. Food Control 2014, 39, 192–197. [Google Scholar] [CrossRef]
- Gambacorta, L.; Magistà, D.; Perrone, G.; Murgolo, S.; Logrieco, A.F.; Solfrizzo, M. Co-occurrence of toxigenic moulds, aflatoxins, ochratoxin A, Fusarium and Alternaria mycotoxins in fresh sweet peppers (Capsicum annuum) and their processed products. World Mycotoxin J. 2018, 11, 159–174. [Google Scholar] [CrossRef]
- Armorini, S.; Altafini, A.; Zaghini, A.; Roncada, P. Ochratoxin A in artisan salami produced in Veneto (Italy). Food Addit. Contam. Part B 2016, 9, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Galaverna, G.; Bertuzzi, T.; Moseriti, A.; Pietri, A.; Dossena, A.; Marchelli, R. Occurrence of ochratoxin A in raw ham muscle, salami and dry-cured ham from pigs fed with contaminated diet. Food Chem. 2010, 120, 978–983. [Google Scholar] [CrossRef]
- Delfino, D.; Lucchetti, D.; Mauti, T.; Mancuso, M.; Di Giustino, P.; Triolone, D.; Vaccari, S.; Bonanni, R.C.; Neri, B.; Russo, K. Investigation of ochratoxin A in commercial cheeses and pork meat products by liquid chromatography-tandem mass spectrometry. J. Food Sci. 2022, 87, 4465–4475. [Google Scholar] [CrossRef]
- Abd-Elghany, S.M.; Sallam, K.I. Rapid determination of total aflatoxins and ochratoxins A in meat products by immuno-affinity fluorimetry. Food Chem. 2015, 179, 253–256. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Nisar, S.; Rafique Asi, M.; Jinap, S. Natural incidence of aflatoxins, ochratoxin A and zearalenone in chicken meat and eggs. Food Control 2014, 43, 98–103. [Google Scholar] [CrossRef]
- Zhang, Z.; Song, Y.; Ma, L.; Huang, K.; Liang, Z. Co-Occurrence of Staphylococcus aureus and Ochratoxin A in Pasteurized Milk. Toxins 2022, 14, 718. [Google Scholar] [CrossRef] [PubMed]
- Žurga, P.; Vahčić, N.; Pasković, I.; Banović, M.; Malenica Staver, M. Occurrence of ochratoxin A and biogenic amines in Croatian commercial red wines. Foods 2019, 8, 348. [Google Scholar] [CrossRef] [PubMed]
- Torović, L.; Lakatoš, I.; Majkić, T.; Beara, I. Risk to public health related to the presence of ochratoxin A in wines from Fruška Gora. LWT 2020, 129, 109537. [Google Scholar] [CrossRef]
- Carballo, D.; Fernández-Franzón, M.; Ferrer, E.; Pallarés, N.; Berrada, H. Dietary exposure to mycotoxins through alcoholic and non-alcoholic beverages in Valencia, Spain. Toxins 2021, 13, 438. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed]
- Corrêa Ribeiro, A.N.; Ferreira, C.D. Mycotoxins in Grains and Cereals Intended for Human Consumption: Brazilian Legislation, Occurrence Above Maximum Levels and Co-Occurrence. Food Rev. Int. 2023, 39, 5920–5933. [Google Scholar] [CrossRef]
- Dongo, L.; Bandyopadhyay, R.; Kumar, M.; Ojiambo, P.S. Occurrence of Ochratoxin A in Nigeria ready for sale cocoa beans. Agric. J. 2008, 3, 4–9. [Google Scholar]
- Prognosis Biotech. Mycotoxins Legislation Database. Available online: https://www.prognosis-biotech.com/mycotoxins-legislation/ (accessed on 28 March 2024).
- FAO WHO Codex Alimentarius—General Standard for Contaminants and Toxins in Food and Feed, CXS 193-1995. Available online: https://www.fao.org/fao-who-codexalimentarius/sh-proxy/en/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B193-1995%252FCXS_193e.pdf (accessed on 25 March 2024).
- Mitchell, N.J.; Chen, C.; Palumbo, J.D.; Bianchini, A.; Cappozzo, J.; Stratton, J.; Wu, F. Risk Assessment of Dietary Ochratoxin A in the United States. Food Chem. Toxicol. 2017, 100, 265–273. [Google Scholar] [CrossRef]
- Ridgway, K.; Scientific, R. Sample preparation for food contaminant analysis. LC GC Eur. 2012, 25, 60–71. [Google Scholar]
- Shephard, G.S. Current status of mycotoxin analysis: A critical review. J. AOAC Int. 2016, 99, 842–848. [Google Scholar] [CrossRef]
- Krska, R.; Schubert-Ullrich, P.; Molinelli, A.; Sulyok, M.; Mac Donald, S.; Crews, C. Mycotoxin analysis: An update. Food Addit. Contam. Part A 2008, 25, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Shephard, G.S. Determination of mycotoxins in human foods. Chem. Soc. Rev. 2008, 37, 2468–2477. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.; Jinap, S.; Soleimany, F. Qualitative and quantitative analysis of mycotoxins. Compr. Rev. Food Sci. Food Saf. 2009, 8, 202–251. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.W.; Subrahmanyam, S.; Piletsky, S. Analytical methods for determination of mycotoxins: A review. Anal. Chim. Acta 2009, 632, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Prakasham, K.; Gurrani, S.; Shiea, J.; Wu, M.T.; Wu, C.F.; Lin, Y.C.; Ponnusamy, V.K. Ultra-sensitive determination of Ochratoxin A in coffee and tea samples using a novel semi-automated in-syringe based coagulant-assisted fast mycotoxin extraction (FaMEx) technique coupled with UHPLC-MS/MS. Food Chem. 2023, 417, 135951. [Google Scholar] [CrossRef] [PubMed]
- Karami-Osboo, R. Nanofluid extraction of Ochratoxin A in food. J. Food Compos. Anal. 2020, 87, 103425. [Google Scholar] [CrossRef]
- Jalili, M.; Jinap, S. Optimization and validation of a method for extraction and quantification of ochratoxin A in black pepper. Int. Food Res. J. 2015, 22, 502–509. Available online: http://www.ifrj.upm.edu.my/22%20(02)%202015/(9).pdf (accessed on 25 March 2024).
- Juan, C.; González, L.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Accelerated Solvent Extraction of Ochratoxin A from Rice Samples. J. Agric. Food Chem. 2005, 53, 9348–9351. [Google Scholar] [CrossRef]
- Hu, X.; Hu, R.; Zhang, Z.; Li, P.; Zhang, Q.; Wang, M. Development of a multiple immunoaffinity column for simultaneous determination of multiple mycotoxins in feeds using UPLC-MS/MS. Anal. Bioanal. Chem. 2016, 408, 6027–6036. [Google Scholar] [CrossRef]
- Koesukwiwat, U.; Sanguankaew, K.; Leepipatpiboon, N. Evaluation of a modified QuEChERS method for analysis of mycotoxins in rice. Food Chem. 2014, 153, 44–51. [Google Scholar] [CrossRef] [PubMed]
- National Toxicology Program. Toxicology and Carcinogenesis Studies of Ochratoxin A (CAS No. 303-47-9) in F344/N Rats (Gavage Studies). Natl. Toxicol Program Tech. Rep. Ser. 1989, 358, 1–142. Available online: https://ntp.niehs.nih.gov/sites/default/files/ntp/htdocs/lt_rpts/tr358.pdf (accessed on 25 March 2024).
- Delaunay, N.; Combès, A.; Pichon, V. Immunoaffinity Extraction and Alternative Approaches for the Analysis of Toxins in Environmental, Food or Biological Matrices. Toxins 2020, 12, 795. [Google Scholar] [CrossRef] [PubMed]
- Cavalera, S.; Anfossi, L.; Di Nardo, F.; Baggiani, C. Mycotoxins Imprinted Polymers: A State-of-the-Art Review. Toxins 2024, 16, 47. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, T.R.; Hoeltz, M.; Einloft, T.C.; Dottori, H.A.; Manfroi, V.; Noll, I.B. Determination of ochratoxin A in wine from the southern region of Brazil by thin layer chromatography with a charge-coupled detector. Food Addit. Contam. Part B 2011, 4, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Antep, H.M.; Merdivan, M. Determination of ochratoxin A in grape wines after dispersive liquid–liquid microextraction using high-performance thin layer and liquid chromatography–fluorescence detection. Hacet. J. Biol. Chem. 2012, 40, 155–163. [Google Scholar]
- Kupski, L.; de Carvalho Silvello, M.A.; Fontes, M.R.V.; Lima, T.S.; Treichel, H.; Furlong, E.B. R. oryzae Cellulases: A New Approach to Degrading Lignocellulosic Material. J. Food Biochem. 2015, 39, 129–138. [Google Scholar] [CrossRef]
- Mohebbi, A.; Nemati, M.; Farajzadeh, M.A.; Mogaddam, M.R.A.; Lotfipour, F. High-performance liquid chromatography-tandem mass spectrometry determination of patulin and ochratoxin A in commercial fruit juices after their extraction with a green synthesized metal organic framework-based dispersive micro solid-phase extraction procedure. Microchem. J. 2022, 179, 107558. [Google Scholar] [CrossRef]
- Badji, T.; Durand, N.; Bendali, F.; Piro-Metayer, I.; Zinedine, A.; Salah-Abbès, J.B.; Brabet, C. In vitro detoxification of aflatoxin B1 and ochratoxin A by lactic acid bacteria isolated from Algerian fermented foods. Biol. Control 2023, 179, 105181. [Google Scholar] [CrossRef]
- Heperkan, Z.D.; Gunalan-Inci, E.; Ceyhan, T. Unexpectedly high patulin contamination and co-occurrence of ochratoxin A in homemade vinegar. Food Control 2023, 148, 109685. [Google Scholar] [CrossRef]
- Chen, D.; Cao, X.; Tao, Y.; Wu, Q.; Pan, Y.; Huang, L.; Yuan, Z. Development of a sensitive and robust liquid chromatography coupled with tandem mass spectrometry and a pressurized liquid extraction for the determination of aflatoxins and ochratoxin A in animal-derived foods. J. Chromatogr. A 2012, 1253, 110–119. [Google Scholar] [CrossRef]
- Meerpoel, C.; Vidal, A.; di Mavungu, J.D.; Huybrechts, B.; Tangni, E.K.; Devreese, M.; De Saeger, S. Development and validation of an LC–MS/MS method for the simultaneous determination of citrinin and ochratoxin A in a variety of feed and foodstuffs. J. Chromatogr. A 2018, 1580, 100–109. [Google Scholar] [CrossRef]
- Modh, H.; Scheper, T.; Walter, J.G. Detection of ochratoxin A by aptamer-assisted real-time PCR-based assay (Apta-qPCR). Eng. Life Sci. 2017, 17, 923–930. [Google Scholar] [CrossRef]
- Rahman, H.U.; Yue, X.; Ren, X.; Zhang, W.; Zhang, Q.; Li, P. Multiplex PCR assay to detect Aspergillus, Penicillium and Fusarium species simultaneously. Food Addit. Contam. Part A 2020, 37, 1939–1950. [Google Scholar] [CrossRef] [PubMed]
- Susca, A.; Anelli, P.; Haidukowski, M.; Probyn, C.E.; Epifani, F.; Logrieco, A.F.; Proctor, R.H. A PCR method to identify ochratoxin A-producing Aspergillus westerdijkiae strains on dried and aged foods. Int. J. Food Microbiol. 2021, 344, 109113. [Google Scholar] [CrossRef]
- Ekhtelat, M. Enzyme-linked Immunosorbent Assay and High-performance Liquid Chromatography analysis of Ochratoxin A in Zataria multiflora and Foeniculum vulgare in Ahvaz (Iran). Asian J. Pharm. 2018, 12, S523 . [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Q.; Tang, X.; Zhang, W.; Li, P. Development of an anti-idiotypic VHH antibody and toxin-free enzyme immunoassay for ochratoxin A in cereals. Toxins 2019, 11, 280. [Google Scholar] [CrossRef] [PubMed]
- Pietri, A.; Leni, G.; Mulazzi, A.; Bertuzzi, T. Ochratoxin A and sterigmatocystin in long-ripened grana cheese: Occurrence, wheel rind contamination and effectiveness of cleaning techniques on grated products. Toxins 2022, 14, 306. [Google Scholar] [CrossRef] [PubMed]
- Hajrulai-Musliu, Z.; Uzunov, R.; Jovanov, S.; Jankuloski, D.; Stojkovski, V.; Pendovski, L.; Sasanya, J.J. A new LC–MS/MS method for multiple residues/contaminants in bovine meat. BMC Chem. 2021, 15, 62. [Google Scholar] [CrossRef]
- Armutcu, C.; Uzun, L.; Denizli, A. Determination of Ochratoxin A traces in foodstuffs: Comparison of an automated on-line two-dimensional high-performance liquid chromatography and off-line immunoaffinity-high-performance liquid chromatography system. J. Chromatogr. A 2018, 1569, 139–148. [Google Scholar] [CrossRef]
- Wei, D.; Pan, A.; Zhang, C.; Guo, M.; Lou, C.; Zhang, J.; Wang, X. Fast extraction of aflatoxins, ochratoxins and enniatins from maize with magnetic covalent organic framework prior to HPLC-MS/MS detection. Food Chem. 2023, 404, 134464. [Google Scholar] [CrossRef]
- Palma, P.; Calderón, R.; Godoy, M.; Vidal, M.; Rivera, A. Human exposure to ochratoxin A and its natural occurrence in spices marketed in Chile (2016–2020): A case study of merkén. J. Food Compos. Anal. 2023, 115, 104885. [Google Scholar] [CrossRef]
- Kresse, M.; Drinda, H.; Romanotto, A.; Speer, K. Simultaneous determination of pesticides, mycotoxins, and metabolites as well as other contaminants in cereals by LC-LC-MS/MS. J. Chromatogr. B 2019, 1117, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Kholová, A.; Lhotská, I.; Uhrová, A.; Špánik, I.; Machyňáková, A.; Solich, P.; Švec, F.; Šatínský, D. Determination of Ochratoxin A and Ochratoxin B in Archived Tokaj Wines (Vintage 1959–2017) Using On-Line Solid Phase Extraction Coupled to Liquid Chromatography. Toxins 2020, 12, 739. [Google Scholar] [CrossRef] [PubMed]
- Potipun, J.; Ruaylarb, W.; Nilthong, R.; Owatworakit, A. Quantification of Ochratoxin A–Producing Fungi in Coffee Products Using Quantitative PCR. Chiang Mai J. Sci. 2018, 17, 39–46. [Google Scholar] [CrossRef]
- Zuo, H.; Wang, X.; Liu, W.; Chen, Z.; Liu, R.; Yang, H.; Ning, B. Nanobody-based magnetic chemiluminescence immunoassay for one-pot detection of ochratoxin A. Talanta 2023, 258, 124388. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.S.; Zhao, J.; Ma, T.T.; Li, Z.Y.; Wang, L.L.; Ji, S.L.; Gong, H.S. Magnetic covalent organic framework for effective solid-phase extraction and HPLC determination of ochratoxin A in food. LWT 2023, 179, 114639. [Google Scholar] [CrossRef]
- Somerson, J.; Plaxco, K.W. Electrochemical aptamer-based sensors for rapid point-of-use monitoring of the mycotoxin ochratoxin a directly in a food stream. Molecules 2018, 23, 912. [Google Scholar] [CrossRef]
- Viter, R.; Savchuk, M.; Iatsunskyi, I.; Pietralik, Z.; Starodub, N.; Shpyrka, N.; Ramanavicius, A. Analytical, thermodynamical and kinetic characteristics of photoluminescence immunosensor for the determination of Ochratoxin A. Biosens. Bioelectron. 2018, 99, 237–243. [Google Scholar] [CrossRef]
- Tang, Z.; Liu, X.; Su, B.; Chen, Q.; Cao, H.; Yun, Y.; Hammock, B.D. Ultrasensitive and rapid detection of ochratoxin A in agro-products by a nanobody-mediated FRET-based immunosensor. J. Hazard. Mater. 2020, 387, 121678. [Google Scholar] [CrossRef]
- Chen, X.; Wu, H.; Tang, X.; Zhang, Z.; Li, P. Recent advances in electrochemical sensors for mycotoxin detection in food. Electroanalysis 2023, 35, e202100223. [Google Scholar] [CrossRef]
- Zhang, S.; Luan, Y.; Xiong, M.; Zhang, J.; Lake, R.; Lu, Y. DNAzyme amplified aptasensing platform for ochratoxin A detection using a personal glucose meter. ACS Appl. Mater. Interfaces 2021, 13, 9472–9481. [Google Scholar] [CrossRef] [PubMed]
- Karczmarczyk, A.; Haupt, K.; Feller, K.H. Development of a QCM-D biosensor for Ochratoxin A detection in red wine. Talanta 2017, 166, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.K.; Joung, H.A.; Jung, M.; Lee, H.; Kim, M.G. Rapid and simple detection of ochratoxin a using fluorescence resonance energy transfer on lateral flow immunoassay (FRET-LFI). Toxins 2019, 11, 292. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Gaba, J.; Singh, K.; Bhatia, Y.; Singh, A.; Singh, N. Recent Advances in Recognition Receptors for Electrochemical Biosensing of Mycotoxins- A Review. Biosensors 2023, 13, 391. [Google Scholar] [CrossRef] [PubMed]
- Clemons, T.D.; Kerr, R.H.; Joos, A. Multifunctional magnetic nanoparticles: Design, synthesis, and biomedical applications. In Comprehensive Nanoscience and Nanotechnology, 2nd ed.; Andrews, D.L., Lipson, R.H., Nann, T., Goreham, R.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 193–210. [Google Scholar] [CrossRef]
- Guan, Y.; Si, P.B.; Yang, T.; Wu, Y.; Yang, Y.H.; Hu, R. A novel method for detection of ochratoxin A in foods—Co-MOFs based dual signal radiometric electrochemical aptamer sensor coupled with DNA walker. Food Chem. 2023, 403, 134316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, D.; Zhang, Y.; Luo, Z.; Zhao, Y.; Zheng, X.; Yang, H.; Yu Zhou, Y. Gold nanoparticle-mediated fluorescence immunoassay for rapid and sensitive detection of Ochratoxin A. Spectrochim. Acta Part A 2024, 304, 1386–1425. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, H.; Aberg, L. Near-infrared spectroscopy for determination of mycotoxins in cereals. Food Control 2003, 14, 229–232. [Google Scholar] [CrossRef]
- Wendy, A.; Anton, S. Fluorescence Polarization Assays in Small Molecule Screening. Expert Opin. Drug Discov. 2011, 6, 17–32. [Google Scholar] [CrossRef]
- Keshri, G.; Magan, N. Detection and differentiation between mycotoxigenic and non-mycotoxigenic strains of two Fusarium spp. using volatile production profiles and hydrolytic enzymes. J. Appl. Microbiol. 2000, 89, 825–833. [Google Scholar] [CrossRef]
- Kabak, B. Aflatoxins and ochratoxin A in chocolate products in Turkey. Food Addit. Contam. Part B 2019, 12, 225–230. [Google Scholar] [CrossRef]
- Adebiyi, J.A.; Kayitesi, E.; Adebo, O.A.; Changwa, R.; Njobeh, P.B. Food fermentation and mycotoxin detoxification: An African perspective. Food Control 2019, 106, 106731. [Google Scholar] [CrossRef]
- Wafula, E.N.; Muhonja, C.N.; Kuja, J.O.; Owaga, E.E.; Makonde, H.M.; Mathara, J.M.; Kimani, V.W. Lactic Acid Bacteria from African Fermented Cereal-Based Products: Potential Biological Control Agents for Mycotoxins in Kenya. J. Toxicol. 2022, 2022, 2397767. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mulè, G. Mycotoxin biotransformation by native and commercial enzymes: Present and future perspectives. Toxins 2017, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Mwabulili, F.; Xie, Y.; Li, Q.; Sun, S.; Yang, Y.; Ma, W. Research progress of ochratoxin a bio-detoxification. Toxicon 2022, 222, 107005. [Google Scholar] [CrossRef] [PubMed]
- Amézqueta, S.; González-Peñas, E.; Murillo-Arbizu, M.; de Cerain, A.L. Ochratoxin A decontamination: A review. Food Control 2009, 20, 326–333. [Google Scholar] [CrossRef]
- Dammak, I.; Hamdi, Z.; Kammoun El Euch, S.; Zemni, H.; Mliki, A.; Hassouna, M.; Lasram, S. Evaluation of antifungal and anti-ochratoxigenic activities of Salvia officinalis, Lavandula dentata, and Laurus nobilis essential oils and a major monoterpene constituent 1,8-cineole against Aspergillus carbonarius. Ind. Crop. Prod. 2019, 128, 85–93. [Google Scholar] [CrossRef]
- El Khoury, R.; Atoui, A.; Mathieu, F.; Kawtharani, H.; El Khoury, A.; Maroun, R.G.; El Khoury, A. Antifungal and Antiochratoxigenic Activities of Essential Oils and Total Phenolic Extracts: A Comparative Study. Antioxidants 2017, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Wu, Z.; Wu, S.; Dai, Y.; Sun, C. Degradation of ochratoxin A by Bacillus amyloliquefaciens ASAG1. Food Addit. Contam. Part A 2015, 32, 564–571. [Google Scholar] [CrossRef]
- Belkacem-Hanfi, N.; Fhoula, I.; Semmar, N.; Guesmi, A.; Perraud-Gaime, I.; Ouzari, H.-I.; Boudabous, A.; Roussos, S. Lactic acid bacteria against post-harvest moulds and ochratoxin A isolated from stored wheat. Biol. Control 2014, 76, 52–59. [Google Scholar] [CrossRef]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int. J. Food Microbiol. 2017, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.S.; Yu, D.; Wu, A. Enzymes for Degradation of Fusarium Mycotoxins. In Food Safety & Mycotoxins; Wu, A., Ed.; Springer: Singapore, 2019; pp. 113–135. [Google Scholar] [CrossRef]
- Amiri, A.; Dugas, R.; Pichot, A.L.; Bompeix, G. In vitro and in vitro activity of eugenol oil (Eugenia caryophylata) against four important postharvest apple pathogens. Int. J. Food Microbiol. 2008, 126, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Belli, N.; Marin, S.; Argiles, E.; Ramos, A.J.; Sanchis, V. Effect of chemical treatments on ochratoxigenic fungi and common mycobiota of grapes (Vitis vinifera). J. Food Prot. 2007, 70, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Lagogianni, C.S.; Tsitsigiannis, D.I. Effective chemical management for prevention of aflatoxins in maize. Phytopathol. Mediterr. 2018, 57, 186–197. [Google Scholar] [CrossRef]
- Oiki, S.; Yaguchi, T.; Urayama, S.I.; Hagiwara, D. Wide distribution of resistance to the fungicides fludioxonil and iprodione in Penicillium species. PLoS ONE 2022, 17, e0262521. [Google Scholar] [CrossRef]
- Özcan, S.; Gökmen, V.U.R.A.L. Alkali-based pre-treatment may prevent ochratoxin A in grapes. World Mycotoxin J. 2016, 9, 517–523. [Google Scholar] [CrossRef]
- Samuel, M.S.; Jeyaram, K.; Datta, S.; Chandrasekar, N.; Balaji, R.; Selvarajan, E. Detection, contamination, toxicity, and prevention methods of ochratoxins: An update review. J. Agric. Food Chem. 2021, 69, 13974–13989. [Google Scholar] [CrossRef]
- Amézqueta, S.; González-Peñas, E.; Lizarraga, T.; Murillo-Arbizu, M.; López Demmunen, A. A simple method reduces Ochratoxin A in contaminated cocoa shells. J. Food Prot. 2008, 71, 1422–1426. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Smith, I.N.; Mikiashvili, N. Reducing Ochratoxin A Content in Grape Pomace by Different Methods. Toxins 2020, 12, 424. [Google Scholar] [CrossRef]
- Qi, L.; Li, Y.; Luo, X.; Wang, R.; Zheng, R.; Wang, L.; Chen, Z. Detoxification of zearalenone and ochratoxin A by ozone and quality evaluation ofozonised corn. Food Addit. Contam. A 2016, 33, 1700–1710. [Google Scholar] [CrossRef]
- Torlak, E. Use of gaseous ozone for reduction of ochratoxin A and fungal populations on sultanas. Aust. J. Grape Wine Res. 2019, 25, 25–29. [Google Scholar] [CrossRef]
- Bhatnagar, D.; Ehrlich, K.; Cleveland, T. Molecular genetic analysis and regulation of aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2003, 61, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Afsah-Hejri, L.; Hajeb, P.; Ehsani, R.J. Application of ozone for degradation of mycotoxins in food: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1777–1808. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Carmody, M.; Sierla, M.; Kangasjärvi, J. Ozone and reactive oxygen species. eLS 2017, 1–9. [Google Scholar] [CrossRef]
- Trombete, F.M.; Freitas-Silva, O.; Saldanha, T.; Venâncio, A.A.; Fraga, M.E. Ozone against mycotoxins and pesticide residues in food: Current applications and perspectives. Int. Food Res. J. 2016, 23, 2545–2556. [Google Scholar]
- Çelebi, Y.; Koç, G.Ç.; Süfer, Ö.; Tekgül Barut, Y.; Şahin Ercan, S.; Yüksel, A.N.; Sezer, S.; Ramniwas, S.; Rastogi, S.; Chandra Khanashyam, A.; et al. Impact of Ozone Treatment on Lipid Oxidation in Foods: A Critical Review. Ozone Sci. Eng. 2024, 1–25. [Google Scholar] [CrossRef]
- Kaur, K.; Pandiselvam, R.; Kothakota, A.; Ishwarya, S.P.; Zalpouri, R.; Mahanti, N.K. Impact of ozone treatment on food polyphenols—A comprehensive review. Food Control 2022, 142, 109207. [Google Scholar] [CrossRef]
- Akbar, A.; Medina, A.; Magan, N. Potential Control of Mycotoxigenic Fungi and Ochratoxin A in Stored Coffee Using Gaseous Ozone Treatment. Microorganisms 2020, 8, 1462. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Nwozo, O.S.; Aja, P.M.; Odongo, G.A. High-pressure acidified steaming with varied citric acid dosing can successfully detoxify mycotoxins. Food Sci. Nutr. 2023, 11, 2677–2685. [Google Scholar] [CrossRef]
- Lee, H.J. Stability of ochratoxin A in oats during roasting with reducing sugars. Food Control 2020, 118, 107382. [Google Scholar] [CrossRef]
- Jalili, M.; Selamat, J.; Rashidi, L. Effect of thermal processing and traditional flavouring mixture on mycotoxin reduction in pistachio. World Mycotoxin J. 2020, 13, 381–389. [Google Scholar] [CrossRef]
- Sumbal, G.A.; Hussain Shar, Z.; Hussain Sherazi, S.T.; Sirajuddin Nizamani, S.M.; Mahesar, S.A. Decontamination of poultry feed from ochratoxin A by UV and sunlight radiations. J. Sci. Food Agric. 2016, 96, 2668–2673. [Google Scholar] [CrossRef] [PubMed]
- Khalil, O.A.; Hammad, A.A.; Sebaei, A.S. Aspergillus flavus and Aspergillus ochraceus inhibition and reduction of aflatoxins and ochratoxin A in maize by irradiation. Toxicon 2021, 198, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Calado, T.; Fernández-Cruz, M.L.; Verde, S.C.; Venâncio, A.; Abrunhosa, L. Gamma irradiation effects on ochratoxin A: Degradation, cytotoxicity and application in food. Food Chem. 2018, 240, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Krogh, P.; Hald, B.; Gjertsen, P.; Myken, F. Fate of ochratoxin A and citrinin during malting and brewing experiments. Appl. Microbiol. 1974, 28, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Scott, P. Effects of processing and detoxification treatments on ochratoxin A: Introduction. Food Addit. Contam. 1996, 13, 19–21. [Google Scholar] [PubMed]
- Aguilar-Alvarez, M.E.; Saucedo-Castañeda, G.; Durand, N.; Perraud-Gaime, I.; González-Robles, R.O.; Rodríguez-Serrano, G.M. The variety, roasting, processing, and type of cultivation determine the low OTA levels of commercialized coffee in Chiapas State, Mexico. Food Control 2021, 126, 108088. [Google Scholar] [CrossRef]
- Cramer, B.; Konigs, M.; Humpf, H.U. Identification and in vitro cytotoxicity of ochratoxin A degradation products formed during coffee roasting. J. Agric. Food Chem. 2008, 56, 5673–5681. [Google Scholar] [CrossRef] [PubMed]
- Sueck, F.; Hemp, V.; Specht, J.; Torres, O.; Cramer, B.; Humpf, H.U. Occurrence of the ochratoxin A degradation product 2’R-ochratoxin A in coffee and other food: An update. Toxins 2019, 11, 329. [Google Scholar] [CrossRef]
- Bittner, A.; Cramer, B.; Harrer, H.; Humpf, H.U. Structure elucidation and in vitro cytotoxicity of ochratoxin α amide, a new degradation product of ochratoxin A. Mycotoxin Res. 2015, 31, 83. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Stępniewska, S.; Modrzewska, M.; Waśkiewicz, A.; Szymczyk, K.; Szafrańska, A. Transformation of ochratoxin A during bread-making processes. Food Control 2021, 125, 107950. [Google Scholar] [CrossRef]
- Zapaśnik, A.; Bryła, M.; Waśkiewicz, A.; Ksieniewicz-Woźniak, E.; Podolska, G. Ochratoxin A and 2′R-Ochratoxin A in selected foodstuffs and dietary risk assessment. Molecules 2022, 27, 188. [Google Scholar] [CrossRef] [PubMed]
- Ouf, S.A.; Basher, A.H.; Mohamed, A.A.H. Inhibitory effect of double atmospheric pressure argon cold plasma on spores and mycotoxin production of Aspergillus niger contaminating date palm fruits. J. Sci. Food Agric. 2015, 95, 3204–3210. [Google Scholar] [CrossRef] [PubMed]
- Durek, J.; Schlüter, O.; Roscher, A.; Durek, P.; Fröhling, A. Inhibition or stimulation of ochratoxin A synthesis on inoculated barley triggered by diffuse coplanar surface barrier discharge plasma. Front. Microbiol. 2018, 9, 2782. [Google Scholar] [CrossRef]
- Hoppanova, L.; Dylikova, J.; Kovacik, D.; Medvecka, V.; Durina, P.; Krystofova, S.; Kalinakova, B. The effect of cold atmospheric pressure plasma on Aspergillus ochraceus and ochratoxin A production. Antonie Leeuwenhoek 2020, 113, 1479–1488. [Google Scholar] [CrossRef] [PubMed]
- Casas-Junco, P.P.; Solís-Pacheco, J.R.; Ragazzo-Sánchez, J.A.; Aguilar-Uscanga, B.R.; Bautista-Rosales, P.U.; Calderón-Santoyo, M. Cold plasma treatment as an alternative for ochratoxin A detoxification and inhibition of mycotoxigenic fungi in roasted coffee. Toxins 2019, 11, 337. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; He, Z.; Ma, C.; Li, W.; Wang, J.; Lin, F.; Liu, X.; Li, L. Evaluation of cold plasma for decontamination of molds and mycotoxins in rice grain. Food Chem. 2023, 402, 134159. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Ding, Y.; An, F.; Wang, L.; Li, S.; Nie, Y.; Zhou, L.; Li, Y.; Wang, C.; Li, S. Degradation of ochratoxin A in aqueous solutions by electron beam irradiation. J. Radioanal. Nucl. Chem. 2015, 306, 39–46. [Google Scholar] [CrossRef]
- Luo, X.; Qi, L.; Liu, Y.; Wang, R.; Yang, D.; Li, K.; Wang, L.; Li, Y.; Zhang, Y.; Chen, Z. Effects of electron beam irradiation on zearalenone and ochratoxin A in naturally contaminated corn and corn quality parameters. Toxins 2017, 9, 84. [Google Scholar] [CrossRef]
- Yang, K.; Li, K.; Pan, L.; Luo, X.; Xing, J.; Wang, J.; Wang, L.; Wang, R.; Zhai, Y.; Chen, Z. Effect of ozone and electron beam irradiation on degradation of zearalenone and ochratoxin A. Toxins 2020, 12, 138. [Google Scholar] [CrossRef]
- Feliciano, C.P. High-dose irradiated food: Current progress, applications, and prospects. Radiat. Phys. Chem. 2018, 144, 34–36. [Google Scholar] [CrossRef]
- Söbeli, C.; Uyarcan, M.; Kayaardı, S. Pulsed UV-C radiation of beef loin steaks: Effects on microbial inactivation, quality attributes, and volatile compounds. Innov. Food Sci. Emerg. Technol. 2021, 67, 102558. [Google Scholar] [CrossRef]
- Chen, J.; Wei, Z.; Wang, Y.; Long, M.; Wu, W.; Kuca, K. Fumonisin B1: Mechanisms of toxicity and biological detoxification progress in animals. Food Chem. Toxicol. 2021, 149, 111977. [Google Scholar] [CrossRef]
- Kaya, Z.; Unluturk, S.; Martin-Belloso, O.; Soliva-Fortuny, R. Effectiveness of pulsed light treatments assisted by mild heat on Saccharomyces cerevisiae inactivation in verjuice and evaluation of its quality during storage. Innov. Food Sci. Emerg. Technol. 2020, 66, 102517. [Google Scholar] [CrossRef]
- Wang, L.; Liu, X.; Cai, R.; Ge, Q.; Zhao, Z.; Yue, T.; Yuan, Y.; Gao, Z.; Wang, Z. Detoxification of ochratoxin A by pulsed light in grape juice and evaluation of its degradation products and safety. Innov. Food Sci. Emerg. Technol. 2022, 78, 103024. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, G.; Yang, Q.; Yang, X.; Zheng, Y.; Liu, Y.; Xing, F. Effects of Light on the Ochratoxigenic Fungi Aspergillus ochraceus and A. carbonarius. Toxins 2021, 13, 251. [Google Scholar] [CrossRef] [PubMed]
- Abdelnaby, A.; Abdelaleem, N.M.; Elshewy, E.; Mansour, A.H.; Ibrahim, S. The efficacy of clay bentonite, date pit, and chitosan nanoparticles in the detoxification of aflatoxin M1 and ochratoxin A from milk. Environ. Sci. Pollut. Res. Int. 2022, 29, 20305–20317. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.M.; Calado, T.; Guimaraes, A.; Rodrigues, M.A.M.; Abrunhosa, L. In vitro adsorption of aflatoxin B1, ochratoxin A, and zearalenone by micronized grape stems and olive pomace in buffer solutions. Mycotoxin Res. 2019, 35, 243–252. [Google Scholar] [CrossRef]
- Nobre, C.; Gonzalez, A.; Losoya, C.; Teixeira, J.A.; Belmares, R.; Abrunhosa, L. Detoxification of ochratoxin A and zearalenone by Pleurotus ostreatus during in vitro gastrointestinal digestion. Food Chem. 2022, 384, 132525. [Google Scholar] [CrossRef]
- Var, I.; Kabak, B.; Erginkaya, Z. Reduction in ochratoxin A levels in white wine, following treatment with activated carbon and sodium bentonite. Food Control 2008, 19, 592–598. [Google Scholar] [CrossRef]
- Carraro Di Gregorio, M.; de Valganon Neeff, D.; Vincenzi Jager, A.; Corassin, C.H.; de Pinho Carão, Á.; de Albuquerque, R.; de Azevedo, A.C.; Fernandes Oliveira, C.A. Mineral adsorbents for prevention of mycotoxins in animal feeds. Toxin Rev. 2014, 33, 125–135. [Google Scholar] [CrossRef]
- Cosme, F.; Inês, A.; Silva, D.; Filipe-Ribeiro, L.; Abrunhosa, L.; Nunes, F.M. Elimination of ochratoxin A from white and red wines: Critical characteristics of activated carbons and impact on wine quality. LWT 2021, 140, 110838. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, N.; Yang, L.; Wang, J.; Song, S.; Nie, D.; Yang, X.; Hou, J.; Wu, A. Cross-linked chitosan polymers as generic adsorbents for simultaneous adsorption of multiple mycotoxins. Food Control 2015, 57, 362–369. [Google Scholar] [CrossRef]
- Solis-Cruz, B.; Hernandez-Patlan, D.; Beyssac, E.; Latorre, J.D.; Hernandez-Velasco, X.; Merino-Guzman, R.; Tellez, G.; Lopez-Arellano, R. Evaluation of Chitosan and Cellulosic Polymers as Binding Adsorbent Materials to Prevent Aflatoxin B1, Fumonisin B1, Ochratoxin, Trichothecene, Deoxynivalenol, and Zearalenone Mycotoxicoses Through an In Vitro Gastrointestinal Model for Poultry. Polymers 2017, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Loffredo, E.; Scarcia, Y.; Parlavecchia, M. Removal of ochratoxin A from liquid media using novel low-cost biosorbents. Environ. Sci. Pollut. Res. Int. 2020, 27, 34484–34494. [Google Scholar] [CrossRef] [PubMed]
- Quintela, S.; Villarán, M.C.; De López Armentia, I.; Elejalde, E. Ochratoxin A removal from red wine by several oenological fining agents: Bentonite, egg albumin, allergen-free adsorbents, chitin and chitosan. Food Addit. Contam. Part A 2012, 29, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, U.; Ain, Q.U.; Yaseen, M.; Fan, X.; Yao, X.; Tong, Z.; Liu, B. Modification of bentonite with orange peels extract and its application as mycotoxins’ binder in buffered solutions and simulated gastrointestinal fluids. J. Clean. Prod. 2020, 267, 122105. [Google Scholar] [CrossRef]
- Appell, M.; Jackson, M.A. Sorption of ochratoxin A from aqueous solutions using beta-cyclodextrin-polyurethane polymer. Toxins 2012, 4, 98–109. [Google Scholar] [CrossRef]
- Alford, R.; Mishael, Y.G. Bifunctional clay based sorbent for ‘Ochratoxin’ removal and wine fining. Food Chem. 2023, 416, 135827. [Google Scholar] [CrossRef]
- Dumeau, F.; Trione, D. Influence de Différents Traitements sur la Concentration en Ochratoxine A des Vins Rouges. Rev. Des Anol. 2000, 95, 37–38. [Google Scholar]
- Savino, F.; Pelle, E.; Palumeri, E.; Oggero, R.; Miniero, R. Lactobacillus reuteri (American Type Culture Collection Strain 55730) versus Simethicone in the Treatment of Infantile Colic: A Prospective Randomized Study. Pediatrics 2007, 119, e124–e130. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Marín, M.; Del Prete, V.; Garcia-Moruno, E.; Fernández-González, C.; Macías-García, A.; Gómez-Serrano, V. The Development of an Activated Carbon from Cherry Stones and Its Use in the Removal of Ochratoxin A from Red Wine. Food Control 2009, 20, 298–303. [Google Scholar] [CrossRef]
- Gambuti, A.; Strollo, D.; Genovese, A.; Ugliano, M.; Ritieni, A.; Moio, L. Influence of Enological Practices on Ochratoxin A Concentration in Wine. Am. J. Enol. Vitic. 2005, 56, 155–162. [Google Scholar] [CrossRef]
- Visconti, A.; Perrone, G.; Cozzi, G.; Solfrizzo, M. Managing Ochratoxin A Risk in the Grape-Wine Food Chain. Food Addit. Contam. 2008, 25, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Quintela, S.; Villarán, M.C.; de López Armentia, I.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Castro, R.I.; Laurie, V.F.; Padilla, C.; Carrasco-Sánchez, V. Removal of Ochratoxin A from Red Wine Using Alginate-PVA-L. plantarum (APLP) Complexes: A Preliminary Study. Toxins 2022, 14, 230. [Google Scholar] [CrossRef] [PubMed]
- Bleve, G.; Grieco, F.; Cozzi, G.; Logrieco, A.; Visconti, A. Isolation of Epiphytic Yeasts with Potential for Biocontrol of Aspergillus carbonarius and A. niger on Grape. Int. J. Food Microbiol. 2006, 108, 204–209. [Google Scholar] [CrossRef]
- Cserha´ti, M.; Kriszt, B.; Krifaton, C.; Szoboszlay, S.; Kukolya, J. Mycotoxin-degradation profile of Rhodococcus strains. Int. J. Food Microbiol. 2013, 166, 176–185. [Google Scholar] [CrossRef]
- Ferenczi, S.; Cserhati, M.; Krifaton, C.; Szoboszlay, S.; Kukolya, J.; Szoke, Z.; Kriszt, B. A new ochratoxin A biodegradation strategy using Cupriavidus basilensis Or16 strain. PLoS ONE 2014, 9, e109817. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhang, H.Y.; Yang, Q.Y. Screening and identification of microorganisms for ochratoxin A degradation. Food Sci. 2014, 35, 154–158. [Google Scholar] [CrossRef]
- De Bellis, P.; Tristezza, M.; Haidukowski, M.; Fanelli, F.; Sisto, A.; Mulè, G.; Grieco, F. Biodegradation of Ochratoxin A by Bacterial Strains Isolated from Vineyard Soils. Toxins 2015, 7, 5079–5093. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, F.; Chiara, M.; Liuzzi, V.C.; Haidukowski, M.; Tristezza, M.; Caterina, M.; D’Erchia, A.M.; Pesole, G.; Horner, D.S.; Mule’, G. Draft genome sequence of Acinetobacter sp. neg1 capable of degrading ochratoxin A. FEMS Microbiol. Lett. 2015, 362, fnv004. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Wang, X.; Zhou, Y.; Wei, C.; Wu, Y. New Luteimonas Species Useful in Preparing Biodegradable Agents for Detoxification Treatment of Moldy Food Materials and Animal Feed; State Intellectual Property Office: Beijing, China, 2016. [Google Scholar]
- Zhang, H.H.; Wang, Y.; Zhao, C.; Wang, J.; Zhang, X.L. Biodegradation of ochratoxin A by Alcaligenes faecalis isolated from soil. J. Appl. Microbiol. 2017, 123, 661–668. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, R.; Mathieu, F.; Atoui, A.; Kawtharani, H.; El Khoury, A.; Afif, C.; El Khoury, A. Ability of soil isolated actinobacterial strains to prevent, bind, and biodegrade ochratoxin A. Toxins 2017, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Luz, C.; Ferrer, J.; Mañes, J.; Meca, G. Toxicity reduction of ochratoxin A by lactic acid bacteria. Food Chem. Toxicol. 2018, 112, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Minguez, C.L.; Garrigues, M.A.R.; Ocana, L.L.; Novella, R.A.; Vinuesa, J.M.; Meca, G. Transformation of ochratoxin A by microorganisms isolated from Tempranillo grapes in wine systems. Am. J. Enol. Viticult. 2020, 71, 167–174. [Google Scholar] [CrossRef]
- Xu, X.; Pang, M.; Liu, J.; Wang, Y.; Wu, X.; Huang, K.; Liang, Z. Genome mining reveals the genes of carboxypeptidase for OTA detoxification in Bacillus subtilis CW14. Int. J. Biol. Macromol. 2021, 186, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Mateo, E.M.; Tarazona, A.; Jiménez, M.; Mateo, F. Lactic Acid Bacteria as Potential Agents for Biocontrol of Aflatoxigenic and Ochratoxigenic Fungi. Toxins 2022, 14, 807. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, C.; Liu, H.; Du, W.; Sun, J.; Zhou, W.; Sun, C. Biodegradation of ochratoxin A by two novel strains of Brevibacillus sp. isolated from wheat (Triticum aestivum L.). Food Biosci. 2023, 54, 102847. [Google Scholar] [CrossRef]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Biodegradation of Ochratoxin A by Aspergillus section Nigri species isolated from French grapes: A potential means of Ochratoxin A decontamination in grape juices and musts. FEMS Microbiol. Lett. 2006, 255, 203–208. [Google Scholar] [CrossRef]
- Valero, A.; Marín, S.; Ramos, A.J.; Sanchis, V. Survey: Ochratoxin A in European Special Wines. Food Chem. 2008, 108, 593–599. [Google Scholar] [CrossRef]
- Xiong, K.; Wang, X.L.; Zhi, H.W.; Sun, B.G.; Li, X.T. Identification and safety evaluation of a product from the biodegradation of ochratoxin A by an Aspergillus strain. J. Sci. Food Agric. 2017, 97, 434–443. [Google Scholar] [CrossRef] [PubMed]
- de Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans can lower ochratoxin A contamination in wine grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Freire, L.; Braga, P.A.C.; Furtado, M.M.; Delafiori, J.; Dias-Audibert, F.L.; Pereira, G.E.; Reyes, F.G.; Catharino, R.R.; Sant’Ana, A.S. From grape to wine: Fate of ochratoxin A during red, rose, and white winemaking process and the presence of ochratoxin derivatives in the final products. Food Control 2020, 113, 107167. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, J.; Zhang, H.; Li, C.; Zhang, X. Ochratoxin A is degraded by Yarrowia lipolytica and generates non-toxic degradation products. World Mycotoxin J. 2016, 9, 269–278. [Google Scholar] [CrossRef]
- Patharajan, S.; Reddy, K.; Karthikeyan, V.; Spadaro, D.; Lore, A.; Gullino, M.L.; Garibaldi, A. Potential of yeast antagonists on in vitro biodegradation of ochratoxin A. Food Control 2011, 22, 290–296. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Venâncio, A.; Teixeira, J.A. Optimization of process parameters for the production of an OTA-hydrolyzing enzyme from Aspergillus niger under solid-state fermentation. J. Biosci. Bioeng. 2011, 112, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Poulsen, C.H.; Dalsgaard, S.; Wang, H.; Nikolaev, I. Food Additive Comprising an Amidase for Detoxifying Ochratoxin; State Intellectual Property Office: Beijing, China, 2015. [Google Scholar]
- Liuzzi, V.C.; Fanelli, F.; Tristezza, M.; Haidukowski, M.; Picardi, E.; Manzari, C. Transcriptional analysis of Acinetobacter sp. neg1 capable of degrading Ochratoxin A. Front. Microbiol. 2016, 7, 2162. [Google Scholar] [CrossRef]
- Wei, W.; Qian, Y.; Wu, Y.; Chen, Y.; Zhou, Y. Detoxification of ochratoxin A by Lysobacter sp. CW239 and characteristics of a novel degrading gene carboxypeptidase cp4. Environ. Pollut. 2020, 258, 113677. [Google Scholar] [CrossRef]
- de Oliveira Garcia, S.; Sibaja, K.V.M.; Nogueira, W.V.; Feltrin, A.C.P.; Pinheiro, D.F.A.; Cerqueira, M.B.R.; Furlong, E.B.; Garda-Buffon, J. Peroxidase as a simultaneous degradation agent of ochratoxin A and zearalenone applied to model solution and beer. Food Res. Int. 2020, 131, 109039. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Cimmarusti, M.T.; Mirabelli, V.; Haidukowski, M.; Logrieco, A.F.; Caliandro, R.; Mule, G. In vitro single and combined mycotoxins degradation by Ery4 laccase from Pleurotus eryngii and redox mediators. Food Control 2018, 90, 401–406. [Google Scholar] [CrossRef]
- Orozco-Cortes, P.C.; Flores-Ortiz, C.M.; Hernandez-Portilla, L.B.; Vazquez Medrano, J.; Rodriguez-Pena, O.N. Molecular docking and in vitro studies of Ochratoxin A (OTA) Biodetoxification Testing Three Endopeptidases. Molecules 2023, 28, 2019. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, G.; Chen, N.; Fang, Z.; Xiao, Y.; Zhang, M.; Gerelt, K.; Qian, Y.; Lai, R.; Zhou, Y. A Superefficient Ochratoxin A Hydrolase with Promising Potential for Industrial Applications. Appl. Environ. Microbiol. 2022, 88, e01964-21. [Google Scholar] [CrossRef] [PubMed]
- Campos-Avelar, I.; Colas de la Noue, A.; Durand, N.; Fay, B.; Schorr-Galindo, S. Minimizing ochratoxin A contamination through the use of actinobacteria and their active molecules. Toxins 2020, 12, 296. [Google Scholar] [CrossRef] [PubMed]
- Muhialdin, B.J.; Saari, N.; Meor Hussin, A.S. Review on the biological detoxification of mycotoxins using lactic acid bacteria to enhance the sustainability of food supply. Molecules 2020, 25, 2655. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, S.; Sontag, G.; Stidl, R.; Ehrlich, V.; Kundi, M.; Knasmueller, S. Detoxification of patulin and ochratoxin A, two abundant mycotoxins, by lactic acid bacteria. Food Chem. Toxicol. 2008, 46, 1398–1407. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Gutiérrez, G.A.; Perraud-Gaime, I.; Escalona-Buendía, H.; Durand, N.; Champion-Martínez, E.I.; Fernández-Soto, R.R.; Saucedo-Castañeda, G.; Rodríguez-Serrano, G. Inhibition of Aspergillus carbonarius growth and Ochratoxin A production using lactic acid bacteria cultivated in an optimized medium. Int. J. Food Microbiol. 2023, 404, 110320. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Liang, Z.; Li, J.; Hao, J.; Xu, Y.; Huang, K.; Tian, J.; He, X.; Xu, W. Ochratoxin A biocontrol and biodegradation by Bacillus subtilis CW14. J. Sci. Food Agric. 2014, 94, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Rodríguez, M.; Luque, M.I.; Justesen, A.F.; Córdoba, J.J. Quantification of Ochratoxin A-Producing Molds in Food Products by SYBR Green and TaqMan Real-Time PCR Methods. Int. J. Food Microbiol. 2011, 149, 226–235. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Song, J.Y.; Park, M.A.; Seo, J.K.; Yang, L.; Lee, C.H.; Cho, K.J.; Ha, J.K. Isolation, screening, and identification of swine gut microbiota with Ochratoxin A biodegradation ability. Asian-Australas. J. Anim. Sci. 2012, 25, 114–121. [Google Scholar] [CrossRef]
- Wei, M.; Dhanasekaran, S.; Ji, Q.; Yang, Q.; Zhang, H. Sustainable and efficient method utilizing N-acetyl-L-cysteine for complete and enhanced ochratoxin A clearance by antagonistic yeast. J. Hazard. Mater. 2023, 448, 130975. [Google Scholar] [CrossRef] [PubMed]
- Zeidan, R.; Ul-Hassan, Z.; Al-Thani, R.; Migheli, Q.; Jaoua, S. In vitro application of a Qatari Burkholderiacepacia strain (QBC03) in the biocontrol of mycotoxigenic fungi and in the reduction of ochratoxin A biosynthesis by Aspergillus carbonarius. Toxins 2019, 11, 700. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Ji, J.; Ye, Y.; Yang, Y.; Yu, J.; Wang, M.; Zheng, Y.; Sun, X. Degradation of Ochratoxin A by a UV-Mutated Aspergillus niger Strain. Toxins 2022, 14, 343. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Bai, X.; Chen, C.; Liu, L.; Li, M.; Xia, X.; Wang, Y. Isolation and identification of a Bacillus megaterium strain with ochratoxin A removal ability and antifungal activity. Food Control 2019, 106, 106743. [Google Scholar] [CrossRef]
- Varga, J.; Rigó, K.; Téren, J. Degradation of Ochratoxin A by Aspergillus species. Int. J. Food Microbiol. 2000, 59, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Péteri, Z.; Tábori, K.; Téren, J.; Vágvölgyi, C. Degradation of Ochratoxin A and other mycotoxins by Rhizopus isolates. Int. J. Food Microbiol. 2005, 99, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Serra, R.; Venâncio, A. Biodegradation of Ochratoxin A by fungi isolated from grapes. J. Agric. Food Chem. 2002, 50, 7493–7496. [Google Scholar] [CrossRef]
- Cho, S.M.; Jeong, S.E.; Lee, K.R.; Sudhani, H.P.; Kim, M.; Hong, S.Y.; Chung, S.H. Biodegradation of Ochratoxin A by Aspergillus tubingensis Isolated from Meju. J. Microbiol. Biotechnol. 2016, 26, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Ponsone, M.L.; Chiotta, M.L.; Combina, M.; Dalcero, A.; Chulze, S. Biocontrol as a strategy to reduce the impact of ochratoxin A and Aspergillus section Nigri in grapes. Int. J. Food Microbiol. 2011, 151, 70–77. [Google Scholar] [CrossRef]
- Ponsone, M.L.; Nally, M.C.; Chiotta, M.L.; Combina, M.; Köhl, J.; Chulze, S.N. Evaluation of the effectiveness of potential biocontrol yeasts against black sur rot and ochratoxin A occurring under greenhouse and field grape production conditions. Biol. Control 2016, 103, 78–85. [Google Scholar] [CrossRef]
- Petruzzi, L.; Corbo, M.R.; Baiano, A.; Beneduce, L.; Sinigaglia, M.; Bevilacqua, A. In vivo stability of the complex ochratoxin A-Saccharomyces cerevisiae starter strains. Food Control 2015, 50, 516–520. [Google Scholar] [CrossRef]
- Angioni, A.; Caboni, P.; Garau, A.; Farris, A.; Orro, D.; Budroni, M.; Cabras, P. In Vitro Interaction Between Ochratoxin A and Different Strains of Saccharomyces cerevisiae and Kloeckeraapiculata. J. Agric. Food Chem. 2007, 55, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migheli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int. J. Food Microbiol. 2014, 189, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Dhanasekaran, S.; Legrand Ngolong Ngea, G.; Abiso Godana, E.; Zhang, X.; Yang, Q.; Zheng, X.; Zhang, H. Cryptococcus podzolicus Y3 degrades ochratoxin A by intracellular enzymes and simultaneously eliminates citrinin. Biol. Control 2022, 168, 104857. [Google Scholar] [CrossRef]
- Taheur, F.B.; Kouidhi, B.; Al Qurashi, Y.M.A.; Salah-Abbès, J.B.; Chaieb, K. Biotechnology of mycotoxins detoxification using microorganisms and enzymes. Toxicon 2019, 160, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Han, M.; Wang, X.; Guo, Y. Ochratoxin A: Overview of Prevention, Removal, and Detoxification Methods. Toxins 2023, 15, 565. [Google Scholar] [CrossRef] [PubMed]
- Dobritzsch, D.; Wang, H.; Schneider, G.; Yu, S. Structural and functional characterization of ochratoxinase, a novel mycotoxin-degrading enzyme. Biochem. J. 2014, 462, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Santos, L.; Venâncio, A. Degradation of Ochratoxin A by Proteases and by a Crude Enzyme of Aspergillus niger. Food Biotechnol. 2006, 20, 231–242. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Venâncio, A. Isolation and purification of an enzyme hydrolyzing ochratoxin A from Aspergillus niger. Biotechnol. Lett. 2007, 29, 1909–1914. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Paterson, R.R.; Venâncio, A. Biodegradation of Ochratoxin A for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef]
- Stander, M.A.; Bornscheuer, U.T.; Henke, E.; Steyn, P.S. Screening of commercial hydrolases for the degradation of ochratoxin A. J. Agric. Food Chem. 2000, 48, 5736–5739. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, P.W.; Petersen, B.O.; Duus, J.Ø.; Zidorn, C.; Frisvad, J.C.; Christophersen, C.; Larsen, T.O. Atlantinone A, a Meroterpenoid Produced by Penicillium ribeum and Several Cheese Associated Penicillium Species. Metabolites 2012, 2, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Zhang, Z.; Xu, X.; Zhang, H.; Zhao, Z.; Liang, Z. Purification and characterization of the enzymes from Brevundimonas naejangsanensis that degrade ochratoxin A and B. Food Chem. 2023, 419, 135926. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Qiu, X.; Li, Y.; Tang, S.; Shen, W.; Xing, C.; Kong, D.; Sheng, J. Carboxypeptidase A immobilization with zeolitic imidazolate framework for enhancement of ochratoxin A degradation ability. Food Agric. Immunol. 2020, 31, 587–599. [Google Scholar] [CrossRef]
- Nahle, S.; Atoui, A.; Assaf, J.C.; El Khoury, A.; Louka, N.; Chokr, A. Time-Dependent Effect of Surface Material on Lactobacillus rhamnosus GG Biofilm Formation and Gene Expression. Microbiology 2023, 92, 55–65. [Google Scholar] [CrossRef]
- Nahle, S.; El Khoury, A.; Savvaidis, I.; Chokr, A.; Louka, N.; Atoui, A. Detoxification approaches of mycotoxins: By microorganisms, biofilms and enzymes. Int. J. Food Contam. 2022, 9, 3. [Google Scholar] [CrossRef]
- Da Cruz Cabral, L.; Pinto, F.V.; Patriarca, A. Application of plant-derived compounds to control fungal spoilage and mycotoxin production in foods. Int. J. Food Microbiol. 2013, 166, 1–14. [Google Scholar] [CrossRef] [PubMed]
- de Coutinho Oliveira, T.L.; de Araújo Soares, R.; Mendes Ramos, E.; das Graças Cardoso, M.; Alves, E.; Piccoli, R.H. Antimicrobial activity of Satureja montana L. essential oil against Clostridium perfringens type A inoculated in mortadella-type sausages formulated with different levels of sodium nitrite. Int. J. Food Microbiol. 2011, 144, 546–555. [Google Scholar] [CrossRef] [PubMed]
- El Mogy, M.M.; Alsanius, B.W. Cassia oil for controlling plant and human pathogens on fresh strawberries. Food Control 2012, 28, 157–162. [Google Scholar] [CrossRef]
- Passone, M.A.; Girardi, N.S.; Etcheverry, M. Evaluation of the control ability of five essential oils against Aspergillus section Nigri growth and ochratoxin A accumulation in peanut meal extract agar conditioned at different water activities levels. Int. J. Food Microbiol. 2012, 159, 198–206. [Google Scholar] [CrossRef]
- Murthy, P.S.; Borse, B.B.; Khanum, H.; Srinivas, P. Inhibitory effects of Ajowan (Trachyspermum ammi) ethanolic extract on A. ochraceus growth and ochratoxin production. Turkish J. Biol. 2009, 33, 211–217. [Google Scholar] [CrossRef]
- Hua, H.; Xing, F.; Selvaraj, J.N.; Wang, Y.; Zhao, Y.; Zhou, L.; Liu, X.; Liu, Y. Inhibitory effect of essential oils on Aspergillus ochraceus growth and ochratoxin A production. PLoS ONE 2014, 9, e108285. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.; Gallo, A.; Scalzo, R.L.; Haidukowski, M.; Picchi, V.; Perrone, G. Inhibition of ochratoxin A production in Aspergillus carbonarius by hydroxycinnamic acids from grapes. World Mycotoxin J. 2015, 8, 283–289. [Google Scholar] [CrossRef]
- El Khoury, R.; Atoui, A.; Verheecke, C.; Maroun, R.; El Khoury, A.; Mathieu, F. Essential Oils Modulate Gene Expression and Ochratoxin A Production in Aspergillus carbonarius. Toxins 2016, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- El-Desouky, T.A. Evaluation of effectiveness aqueous extract for some leaves of wild edible plants in Egypt as anti-fungal and anti-toxigenic. Heliyon 2021, 7, e06209. [Google Scholar] [CrossRef] [PubMed]
- Naz, G.; Anjum, A.A.; Nawaz, M.; Iqbal, S.; Azeem, S.; Ali, T.; Manzoor, R. Evaluation of Cinnamomum verum Essential Oils against Ochratoxin A-producing Aspergillus parasiticus in Stored Wheat, Maize, and Rice. Polish J. Environ. Stud. 2023, 32, 667–676. [Google Scholar] [CrossRef]
- El-Desouky, T.; Ammar, H. Honey-mediated silver nanoparticles and their inhibitory effect on aflatoxins and ochratoxin A. J. Appl. Pharm. Sci. 2016, 6, 83–90. [Google Scholar] [CrossRef]
- Pushpalatha, R.; Selvamuthukumar, S.; Kilimozhi, D. Cross-linked, cyclodextrin-based nanosponges for curcumin delivery: Physicochemical characterization, drug release, stability, and cytotoxicity. J. Drug Deliv. Sci. Technol. 2018, 45, 45–53. [Google Scholar] [CrossRef]
- Dhakar, N.K.; Matencio, A.; Caldera, F.; Argenziano, M.; Cavalli, R.; Dianzani, C.; Zanetti, M.; Lopez-Nicolas, J.M.; Trotta, F. Comparative evaluation of solubility, cytotoxicity, and photostability studies of resveratrol and oxyresveratrol loaded nanosponges. Pharmaceutics 2019, 11, 545. [Google Scholar] [CrossRef]
- Redondo-Blanco, S.; Fernandez, J.; Lopez-Ibanez, S.; Miguelez, E.M.; Villar, C.J.; Lombo, F. Plant phytochemicals in food preservation: Antifungal bioactivity: A review. J. Food Prot. 2019, 83, 163–171. [Google Scholar] [CrossRef]
- Sajid, M.; Cameotra, S.S.; Sajjad, M.; Khan, A.; Ahmad, I. Nanoparticle-based delivery of phytomedicines: Challenges and opportunities. In New Look to Phytomedicine; Ahmad Khan, M.S., Ahmad, I., Chattopadhyay, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 597–623. [Google Scholar] [CrossRef]
- Thipe, V.C.; Panjtan Amiri, K.; Bloebaum, P.; Raphael Karikachery, A.; Khoobchandani, M.; Katti, K.K.; Jurisson, S.S.; Katti, K.V. Development of resveratrol-conjugated gold resveratrol corona on anti-tumor efficacy against nanoparticles: Interrelationship of increased breast, pancreatic, and prostate cancers. Int. J. Nanomed. 2019, 14, 4413–4428. [Google Scholar] [CrossRef] [PubMed]
- Pawar, A.Y.; Naik, A.K.; Jadhav, K.R. Nanosponges: A novel drug delivery system. Asian J. Pharm. Pharmacol. 2016, 10, S456–S463. [Google Scholar] [CrossRef]
- Sherje, A.P.; Dravyakar, B.R.; Kadam, D.; Jadhav, M. Cyclodextrin-based nanosponges: A critical review. Carbohydr. Polym. 2017, 173, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, H.; Venkatesh, D.N.; Kuila, A.; Kumar, K.H. Nanosponges: A review. Int. J. Appl. Pharm. 2018, 10, 1. [Google Scholar] [CrossRef]
- Haimhoffer, Á.; Rusznyák, Á.; Réti-Nagy, K.; Vasvári, G.; Váradi, J.; Vecsernyés, M.; Bácskay, I.; Fehér, P.; Ujhelyi, Z.; Fenyvesi, F. Cyclodextrins in drug delivery systems and their effects on biological barriers. Sci. Pharm. 2019, 87, 33. [Google Scholar] [CrossRef]
- Ananya, K.V.; Preethi, S.; Amit, B.P.; Gowda, D.V. Recent review on Nano sponge. Int. J. Res. Pharm. Sci. 2020, 11, 1085–1096. [Google Scholar] [CrossRef]
- Asghari, F.; Jahanshiri, Z.; Imani, M.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Antifungal nanomaterials: Synthesis, properties, and applications. In Nanobiomaterials in Antimicrobial Therapy; Mihai Grumezescu, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 343–383. [Google Scholar] [CrossRef]
- Appell, M.; Evans, K.O.; Jackson, M.A.; Compton, D.L. Determination of ochratoxin A in grape juice and wine using nanosponge solid phase extraction clean-up and liquid chromatography with fluorescence detection. J. Liq. Chromatogr. Relat. Technol. 2018, 41, 949–954. [Google Scholar] [CrossRef]




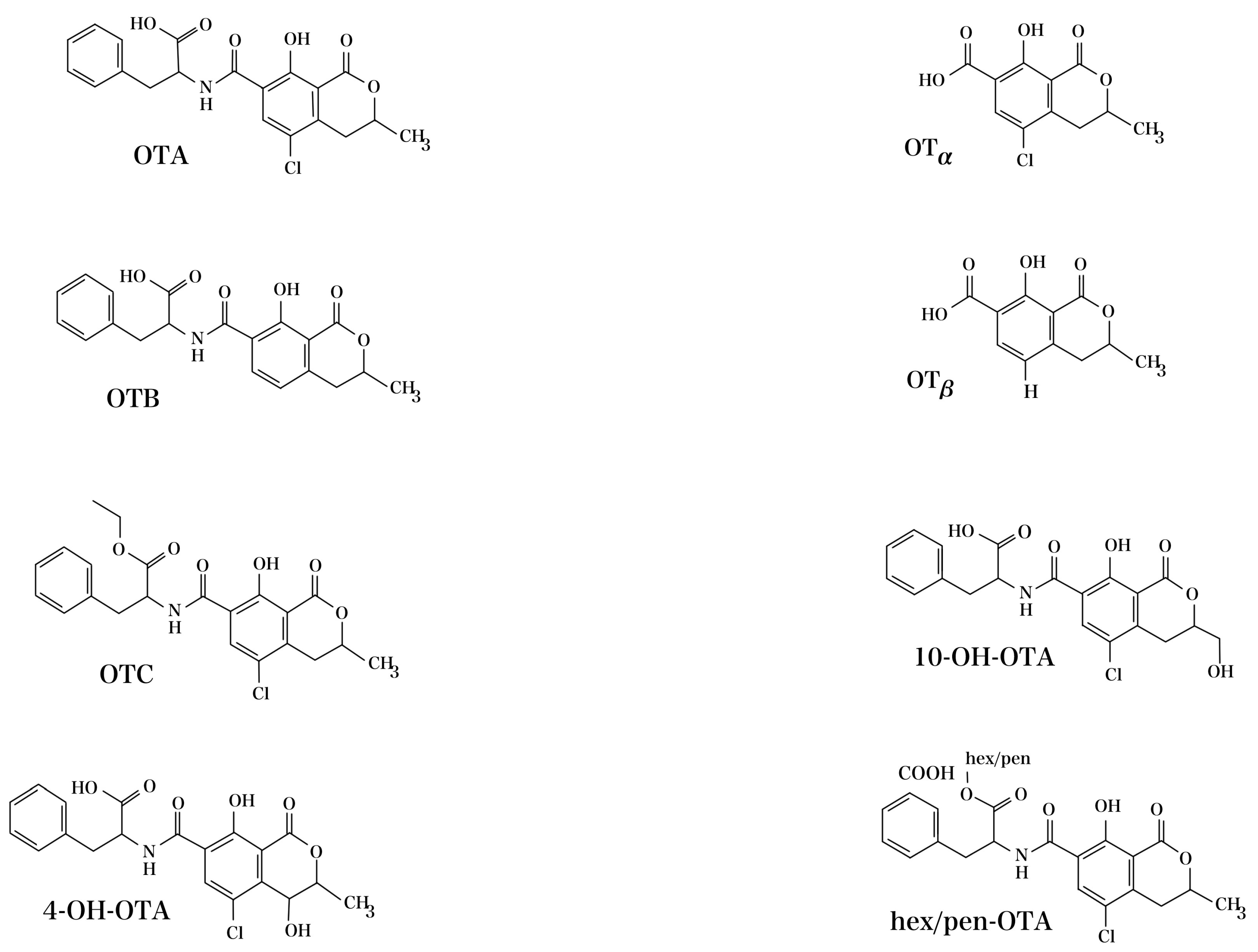{kind=link}
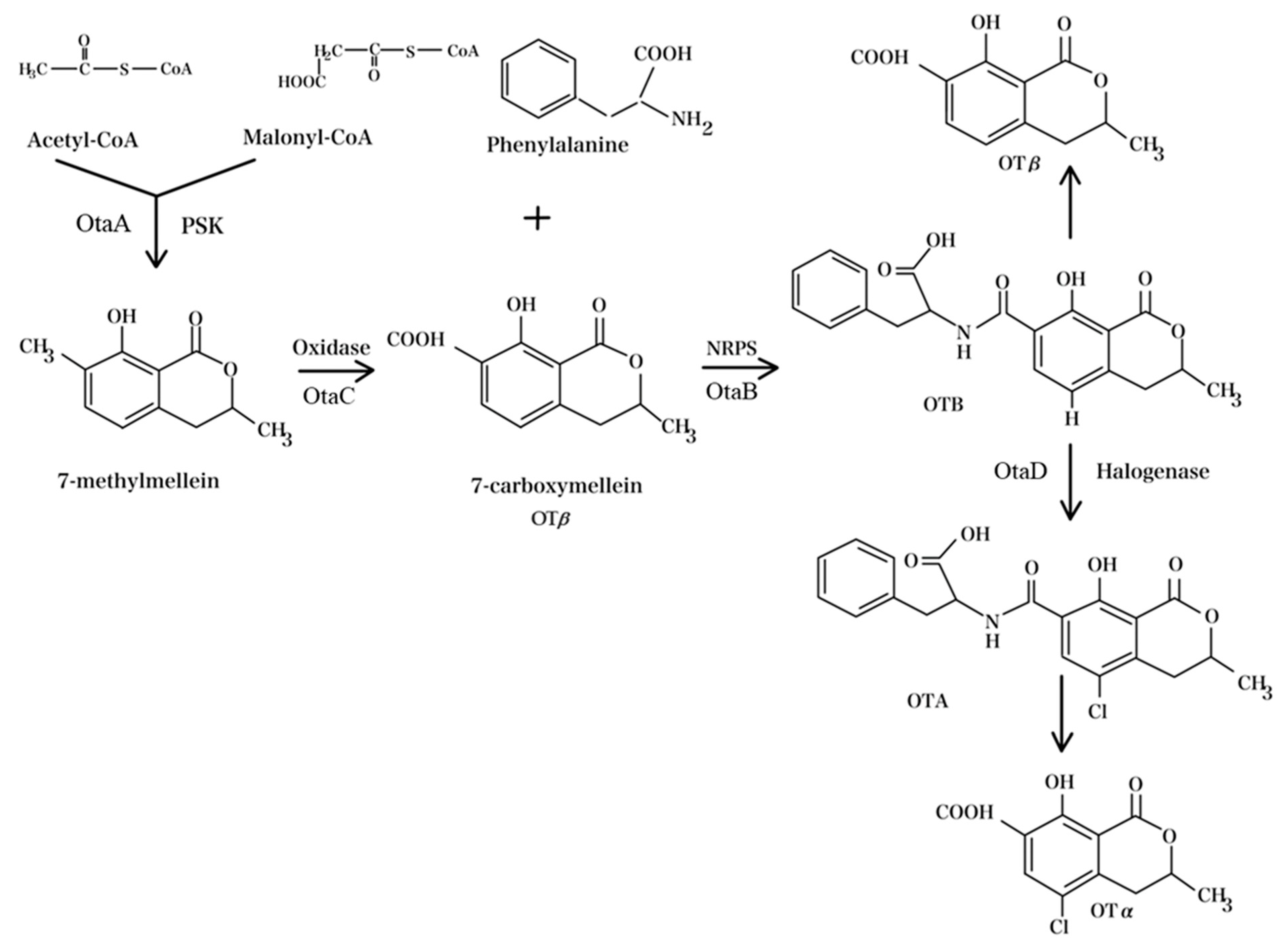{kind=link}
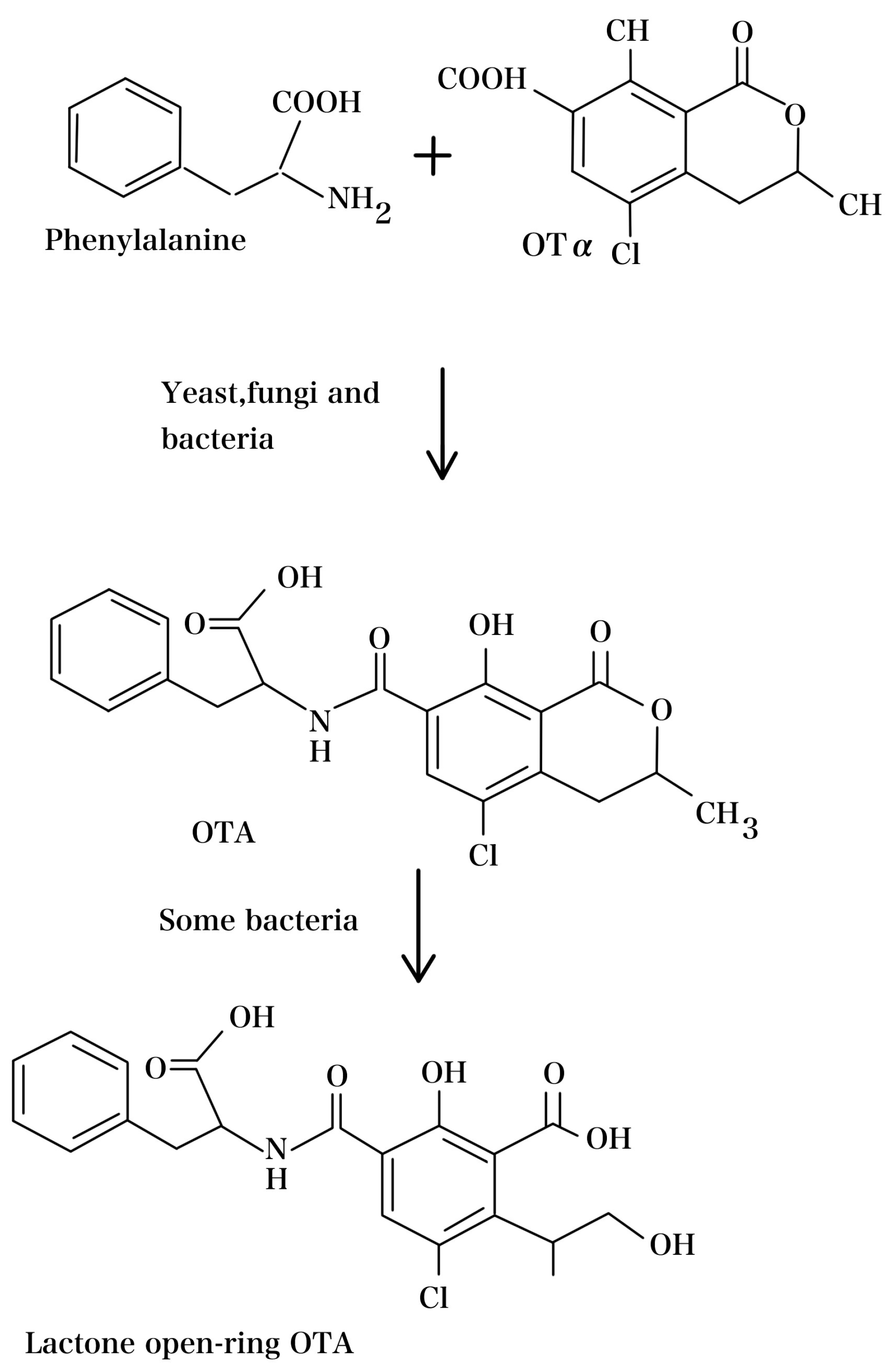{kind=link}
| Organism | Section | References |
|---|---|---|
| Aspergillus westerdijkiae | Circumdati | [35] |
| A. melleus | Circumdati | |
| A. ochraceus | Circumdati | |
| A. steynii | Circumdati | |
| A. subramanianii | Circumdati | |
| A. sesamicola | Circumdati | |
| A. affinis | Circumdati | [36] |
| A. muricatus | Circumdati | |
| A. occultus | Circumdati | |
| A. ochraceopetaliformis | Circumdati | |
| A. flocculosus | Circumdati | |
| A. pseudoelegans | Circumdati | [37] |
| A. roseoglobulosus | Circumdati | |
| A. sclerotiorum | Circumdati | [35] |
| A. persii | Circumdati | |
| A. salwaensis | Circumdati | |
| A. carbonarius | Nigri | [38,39] |
| A. niger | Nigri | |
| A. lacticoffeatus | Nigri | |
| A. sclerotioniger | Nigri | |
| A. alliaceus | Flavi | [40] |
| A. avenaceus | Flavi | |
| A. bertholletius | Flavi | |
| A. coremiiformis | Flavi | |
| A. leporis | Flavi | [41,42,43,44] |
| A. nomius | Flavi | |
| A. tamarii | Flavi | |
| A. pseudotamarii | Flavi | |
| A. vandermerwei | Flavi | |
| A. neoalliaceus | Flavi |
| Matrix | Country | Number of Sample | Occurrence (%) | Maximum µg/kg | Mean µg/kg | Ref. |
|---|---|---|---|---|---|---|
| Corn, wheat | Pakistan | 40 | 27.5 | 360 | / | [77] |
| Rice | Iran | 65 | 4.6 | 11.5 | 5.02 | [78] |
| Barley and wheat | USA | 262 | 12.2 | 185.2 | / | [79] |
| Corn, rice, wheat, and oat-based foods | USA | 489 | 41 | 9.3 | / | [80] |
| Wheat | Canada | 232 | 2.2 | / | 14.7 | [81] |
| Wheat grain clumps around or under manhole openings of storage bins | Canada | 5 | 100 | 370 | [81] | |
| Wheat and derived samples | Algeria | 81 | 76.54 | 34.75 | / | [82] |
| Cereal-based foods | Algeria | 16.9 | 50 | / | 0.15 | [83] |
| Rye | Poland | 60 | 3 | 2.75 | / | [84] |
| Fermented coffees | Brazil | 14 | 21.4 | 0.87 | 0.18 | [85] |
| Roasted and instant coffee | Czech | 103 | 80.6 | 12.8 | / | [86] |
| Coffee bean roasted coffee and soluble coffee | Argentina | 51 | 69 | 20.3 | / | [87] |
| Roasted and instant coffee | Chile | 63 | 33 | 7.25 | 1.3 | [88] |
| Roasted coffee | Spain | 72 | 48.6 | 4.21 | 2.17 | [89] |
| Cocoa bean | Brazil | 123 | 22.8 | 7.2 | 1.2 | [90] |
| Cocoa and chocolate | Canada | 60 | 100 | 7.8 | 0.95 | [91] |
| Dried figs | Turkey | 100 | 8 | 1.72 | 0.64 | [92] |
| Dried grape | Iran | 66 | 40.9 | 8.4 | 2.98 | [93] |
| Palm dates | Tunisia and Algeria | 27 | 11 | 6.07 | 58.7 | [94] |
| Raisin | USA | 40 | 93 | 11.4 | 0.7 | [95] |
| Dried fruit and nuts | China | 253 | 1.6 | 9.39 | 6.23 | [96] |
| Chili sauce | Pakistan | 252 | 71 | 114 | / | [97] |
| Allspice, pepper, chili, cinnamon, ginger, and mixture | Italy | 94 | 30 | 34 | 7.1 | [98] |
| Grounded sweet pepper | Italy | 8 | 100 | / | 23.6 | [99] |
| Dried sweet pepper | Italy | 23 | 39 | / | 53.9 | [99] |
| Salami | Italy | 50 | 10 | 103.69 | / | [100] |
| Dry-cured ham | Italy | 110 | 76.4 | 5.64 | / | [101] |
| Dry-cured ham | Italy | 18 | 22.2 | 69.3 | / | [102] |
| Cheese | Italy | 30 | 13.3 | 4.7 | / | [102] |
| Cheese | Italy | 84 | 8.3 | 22.4 | / | [20] |
| Beef burger | Egypt | 25 | 100 | / | 4.55 | [103] |
| Chicken meat | India | 115 | 41 | / | 1.41 | [104] |
| Eggs | India | 80 | 35 | / | 1.17 | [104] |
| Milk | China | 120 | 25.8 | 18.8 | / | [105] |
| Red wine | Croatia | 110 | 98.2 | 0.163 | 0.040 | [106] |
| Red, rose, and white wine | Serbia | 113 | 52.2 | 0.134 | 0.026 | [107] |
| Wine | Spain | 40 | 47 | 2.28 | 1.13 | [108] |
| Beer | Mainly Portugal | 85 | 10.6 | 11.25 | / | [85] |
| OTA Maximum Level in Different Countries (μg/kg) | |||||||
|---|---|---|---|---|---|---|---|
| Matrix | European Union | Brazil | China | Turkey | Egypt | Vietnam | Codex Alimentarius |
| Dried vine fruits (currants, raisins and sultanas) and dried figs | 8.0 | 10.0 | 10.0 | 10.0 | 3.0 | ||
| Other dried fruits | 2.0 | 10.0 | |||||
| Dried herbs | 10.0 | ||||||
| Sunflower seeds, pumpkin seeds, (water) melon seeds, hempseeds, soybeans | 5.0 | ||||||
| Unprocessed cereal grains | 5.0 | 10.0 | 5.0 | 5.0 | 5.0 | 5.0 | 5 |
| Bakery wares, cereal snacks and breakfast cereals | 2.0 | 3.0 | 3.0 | 3.0 | |||
| Roasted coffee beans and ground roasted coffee | 3.0 | 10.0 | 5.0 | 5.0 | 5.0 | 5.0 | |
| Soluble coffee (instant coffee) | 5.0 | 10.0 | 10.0 | 10.0 | 10.0 | ||
| Cocoa powder | 3.0 | ||||||
| Cocoa beans | 10.0 | ||||||
| Dried spices | 15.0 | 15.0 | |||||
| Capsicum spp. (dried fruits including chillies, chilli powder, cayenne or paprika) | 20.0 | 30.0 | 30.0 | 20.0 | |||
| Wine, fruit wine, aromatized wine, grape juice, grape must | 2.0 | 2.0 | 2.0 | 2.0 | 2.0 | ||
| Baby food and processed cereal-based food, for infants and young children | 0.50 | 2.0 | 0.50 | 0.5 | |||
| Methods | Matrix | LOD * | LOQ * | Ref. |
|---|---|---|---|---|
| LC-MS | Cheese | 0.05 µg/kg | 0.15 µg/kg | [143] |
| LC-MS/MS | Bovine meat | 0.059–291.36 μg/kg | 0.081–328.13 μg/kg | [144] |
| 2D-HPLC | Beer, wine, corn, coffee | 21.2 pg/mL | 64.3 pg/mL | [145] |
| HPLC-MS/MS | Maize | 0.26 μg/kg | 0.87 μg/kg | [146] |
| UHPLC-MS/MS | Coffee | 3 pg/g | 1 pg/g | [121] |
| HPLC-fluorescence detection (FLD) | Spices | 0.03 ng/g | 1.0 ng/g | [147] |
| LC- LC-MS/MS | Cereals | 1 pg/kg | _ | [148] |
| Nanofluid extraction coupled with HPLC with fluorescence detector HPLC–FLD | Food | 0.2 μg/kg | 0.5 μg/kg | [122] |
| SPE–HPLC | Wine | 0.03 µg/L | 0.10 µg/L | [149] |
| qPCR | Coffee | 3.85 × 103 copies of the PKS gene | _ | [150] |
| magnetic beads (MBS) ELISA | Cereal | 0.07 ng/mL | 0.249–5.28 ng/mL | [151] |
| magnetic solid-phase extraction (MSPE) GC-MS/MS | Beer, Chinese Baijiu and vinegar | 0.03 µg/L | 0.1–800 μg/L | [152] |
| ELISA | Cereals | 0.001 ng/mL | 0.003–0.673 ng/mL | [142] |
| Electrochemical aptamer sensors | Food | 6.5 µM | _ | [153] |
| Photoluminescence immunosensor | Food commodities | 0.01 ng/mL | _ | [154] |
| Nanobody (Nb) Förster Resonance Energy Transfer (FRET) immunosensor | Cereal | 5 pg/mL | _ | [155] |
| Electrochemical (EC) sensors | Food | 0.38 ng/mL | 1–20 ng/mL | [156] |
| Aptamer based sensors | Food | 0.88 pg/mL | 1 pg to 300 ng/mL | [157] |
| Fluonanobodies (FN) nanosensor | Cereals | 5 pg/mL | 5–5000 pg/mL | [10] |
| Quartz crystal microbalance with dissipation monitoring (QCM-D) | Red wine | 0.16 ng/mL | 0.55 ng/mL | [158] |
| FRET-LFI | Coffee | 0.88 ng/mL | _ | [159] |
| Method | Agents | Condition | Food Matrix | Effect on OTA | Ref. |
|---|---|---|---|---|---|
| Alkalinization | Potassium carbonate | 40 °C for pH 10 | Grape | Reduced by up to 50% | [184] |
| Ammonia | High pressure (60 psi) and normal temperature | Wheat | Reduced by 79% | [185] | |
| Potassium carbonate | 2% solution at 90 °C and 6894.76 kPa for 10 min. | Cocoa shells | Reduced by 93% | [186] | |
| Acidification | Acetic acid, citric acid, lactic acid, or hydrochloric acid | 50 °C for 24 h at pH 2 | Grape pomaces | Reduced by up to 67.23% (lactic acid) | [187] |
| Ozonation | ozone | 100 mg/L for 180 min | Corn | Reduced by 70.7% | [188] |
| ozone | gaseous ozone at 12.8 mg/L for 240 min | Sultanas | Reduced by 82.5% | [189] |
| Method | Condition | Food Matrix | Effect | Ref. |
|---|---|---|---|---|
| Heat treatment | 120 °C and 180 °C for 30 min and 60 min | Oats | Reduction of 2–18% | [198] |
| 150 °C for 50 min | Pistachios | Reduction of over 60% | [199] | |
| Ultraviolet radiation | 180 min of UV radiation | Poultry feed | Completely decontaminated | [200] |
| Gamma radiation | 20 kGy, | Corn | Reduction of 61.1% | [201] |
| 30.5 kGy | Wheat flour, Grape juice, and wine | Reduction of 24%, 12%, and 23%, respectively | [202] |
| Material | Adsorbents | Medium/Matrix | Removal | Ref. |
|---|---|---|---|---|
| Ground nuts, coconut fiber, waste coffee grounds, and citrus peel | Liquid (ethanol/water mixture = 14/86, v/v) | Up to 100% | [233] | |
| Natural | Powdered Pleurotus ostreatus | in vitro gastrointestinal digestion | 85% at the end of the intestinal phase | [227] |
| Egg albumin, gelatin, chitin, and chitosan | Red Wine | Between 13% and 34% | [234] | |
| Inorganic Mineral | Activated carbon 5 mg/mL | In vitro | More than 89% | [226] |
| Activated carbon | PBS and wine | 87–100% | [228] | |
| Activated carbon | Red and white wine | Up to 100% | [230] | |
| Bentonite-orange peel extract | Gastrointestinal fluids | Up to 2.13 mg/g | [235] | |
| Organic Synthetic | Cyclodextrin-polyurethane polymer | Wine Samples | Up to 10 μg/L | [236] |
| Clay polymer nanocomposite | grape juice and wine | Up to 92% (15 μg/L) | [237] |
| Microorganism/Enzymes | Source | Reaction Condition | OTA (μg/mL) | Degradation Rate (%) | Ref. |
|---|---|---|---|---|---|
| Bacteria/Actinobacter | |||||
| Rhodococcus erythropolis GD2A, BRB 1AB | Natural soil | 72 h/liquid medium | 2 | 27–34 | [246] |
| Cupriavidus basilensis ŐR16 | Soil | 5 d/liquid medium | 100 | >90 | [247] |
| Brevundimonas vesicularis | Vineyard soil | 28 °C/water | 1 | 100 | [248] |
| Acinetobacter calcoaceticus strain 396.1 | Vineyard soil | 6 d/liquid medium | 1 | 82 | [249] |
| Acinetobacter sp. Neg1 | Vineyard soil | 6 d/liquid medium | 1 | >70 | [250] |
| Bacillus amyloliquefaciens ASAG1 a | Grain, maize | 31 °C, 10 h/nutrient culture media | 1 | 98.5 | [176] |
| Luteimonas sp. CW574 | Soil and moldy food | 48 h/feed | 0.02 | 48.3 | [251] |
| Alcaligenes faecalis ASAGF 0D-1 | Soil | 48 h/liquid medium | 1 | 92 | [252] |
| Streptomyces AT10, AT8, SN7, MS1, ML5, G10,PT1 | Soil | 5 d/liquid medium | 0.095 | 23–53 | [253] |
| Bifidobacterium bifidum CECT 870T, B. breve CECT 4839T; Lactobacillus casei CECT 475T, Lactobacillus casei CECT 4040, L. casei CECT 4045, L. delbrueckii bulgaricus CECT 4005, L. johnsonii CECT 289, L. paracasei CECT 4022, L. plantarum CECT 220, L. plantarum CECT 221, L. plantarum CECT 222, L. plantarum CECT 223, L. plantarum CECT 748, L. plantarum CECT 749, L. rhamnosus CECT 278T, L. rhamnosus CECT 288, L. salivarius CECT 4062 | Spanish Type Culture Collection | 24 h/liquid medium | 0.6 | 30–97 | [254] |
| Lactobacillus rhamnosu | Winegrapes | TSB medium | 1 | 55 | [255] |
| Bacillus subtilis CW14 | fresh elk | 30 °C, 24 h/liquid medium | 1 | 71.3 | [256] |
| Leuconostoc paracasei ssp. Paracasei (3T3R1), L. mesenteroides ssp. Dextranicum (T2MM3) | Food | 20, 25, 30 °C, 10 days/MRS-CYA20S medium | NR | 7.3–100 | [257] |
| Brevibacillus sp.(ALJ01), Brevibacillus schisleri (ALJ02) | 24 h/MSM medium | 5 | 89.8, 96.5 | [258] | |
| Fungi | |||||
| A. carbonarius SA332 | French grapes | 5 d/liquid medium | 2 | 83 | [259] |
| Botrytis cinerea UdLTA 3·95, UdLTA 3·102, UdLTA 3·115 | Grapes | 7 d/solid grape synthetic medium | 1 | 24.2–26.7 | [260] |
| 124.niger M00120 | Soil | 2 d/liquid medium | 0.2 | 99 | [261] |
| Aureobasidium pullulans AU14-3-1, AU18-3B, AU34-2, LS30 | Apple leaves, Plum fruits, Grapevine leaves, Apple | 6 d/liquid medium | 0.8 | 75–90.5 | [262] |
| A. carbonarius 10614, A. niger 10,443 | Wine grapes | 25 °C, 7 d/Wine | 6 | 83.44 | [263] |
| Yeast | |||||
| Yarrowia lipolytica Y-2 | Vineyard soil | 2 d/liquid medium | 1 | 84 | [249] |
| Yarrowia lipolytica | vineyard | 2 d/liquid medium | 1 | 88 | [264] |
| Yarrowia lipolytica Y-2 | The surface of grapes | 28 °C/liquid medium | 1 | 97.2 | [264] |
| Metschnikowia pulcherrima MACH1, M320; Kloeckera lindneri GAL5; Pichia guilliermondii M8, M29; Rhodococcus erythropolis AR14 | Agroinnova culture collection centre (Italy) | 15 d/liquid medium | 7.5 | 25.8–84 | [265] |
| Enzyme | |||||
| Carboxypeptidase | Bacillus amyloliquefaciens | Overnight/Tris-HCL | 10 | 72 | [177] |
| Hydrolase | Aspergillus niger MUM 03 | 4 h/phosphate buffer | 1000 | Up to 154 ng/min g | [266] |
| Amidase 2 | Aspergillus niger | 30 min/Mops-NaOH | 0.85 | 83 | [267] |
| Carboxypeptidase PJ_1540 | Acinetobacter sp. Neg1 | Overnight/Tris buffer | 1 | 33 | [268] |
| Carboxypeptidase cp4 | Lysobacter sp. CW239 | 24 h | 250 | 86.2 | [269] |
| Peroxidase | Armoracia rusticana | 72 h/PBS | 0.01 | 27 | [270] |
| Laccase | Pleurotus eryngii | 72 h/sodium acetate buffer | 0.05 | 27 | [271] |
| Carboxypeptidase | Bacillus subtilis | 48 h/PBS | 1 | 71.3 | [256] |
| Metalloendopeptidase | Bacillus subtilis | 1 h, 41 °C/Phosphate buffer pH 4.6, 5 and 7 | 0.1 | between 8.2 and 45.3% | [272] |
| Amidohydrolase ADH3 | Stenotrophomonas acidaminiphila CW117 | 24 h, 60 °C/PBS, pH 7.2 | 0.05 | 100 | [273] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Miri, Y.; Benabdallah, A.; Chentir, I.; Djenane, D.; Luvisi, A.; De Bellis, L. Comprehensive Insights into Ochratoxin A: Occurrence, Analysis, and Control Strategies. Foods 2024, 13, 1184. https://doi.org/10.3390/foods13081184
Ben Miri Y, Benabdallah A, Chentir I, Djenane D, Luvisi A, De Bellis L. Comprehensive Insights into Ochratoxin A: Occurrence, Analysis, and Control Strategies. Foods. 2024; 13(8):1184. https://doi.org/10.3390/foods13081184
Chicago/Turabian StyleBen Miri, Yamina, Amina Benabdallah, Imene Chentir, Djamel Djenane, Andrea Luvisi, and Luigi De Bellis. 2024. "Comprehensive Insights into Ochratoxin A: Occurrence, Analysis, and Control Strategies" Foods 13, no. 8: 1184. https://doi.org/10.3390/foods13081184
APA StyleBen Miri, Y., Benabdallah, A., Chentir, I., Djenane, D., Luvisi, A., & De Bellis, L. (2024). Comprehensive Insights into Ochratoxin A: Occurrence, Analysis, and Control Strategies. Foods, 13(8), 1184. https://doi.org/10.3390/foods13081184