1. Introduction
Sanghuangporus sanghuang is a medicinal fungus that has been used in traditional medicine for centuries in Southeast Asia [
1].
S. sanghuang parasitizes on mulberry trees and is found in the north-eastern regions of Asia [
2].
S. sanghuang is associated with anticancer activities [
3] as well as antioxidant and anti-inflammatory properties [
4]. The mycelium of
S. sanghuang is rich in histones and improves sleep [
5].
Sanghuangporus spp. are rich in polysaccharides, flavonoids, terpenes and other active substances.
Sanghuangporus spp. polysaccharide is an important active substance of
Sanghuangporus spp. Intracellular polysaccharides are mainly derived from fermentation broth [
6,
7,
8,
9]. The extracellular polysaccharides of
S. sanghuang are widely-studied as bioactive compounds and show a broad range of pharmacological activities, including anticancer [
10], hypoglycemic [
11], antioxidant [
12], and immunity-enhancing [
13] properties. Liu et al. [
14] found that
Phellinus baumii polysaccharide can promote the proliferation of RAW264.7 cells, enhance their phagocytic ability, promote their secretion of IL-6 and TNF-α cytokines, and have immunomodulatory activity. Dong et al. [
15] have shown that the crude extracellular polysaccharide (EPS) in
Phellinus igniarius liquid fermentation broth can significantly inhibit the growth of mouse sarcoma S180 and mouse liver cancer H22.
As
S. sanghuang has immense medicinal value, there is an increased demand for various natural products and health-improving nutritional food supplements containing
S. sanghuang. Therefore, there is an increasing demand for culturing
S. sanghuang on a large-scale. However, the yield of wild
S. sanghuang is poor and insufficient to meet the medical and market demands. Although the cultivation of fungi such as
Sanghuangporus vaninii,
Inonotus hispidus, and
Sanghuangporus baumii has been achieved successfully, the cultivation of
Sanghuangporus sanghuang has remained difficult. Because the types of mulberry trees and habitats (temperature, humidity, light, oxygen concentration, etc.) that parasitize
Sanghuangporus sanghuang are varied, their quality evaluation lacks scientificity and rigor. The current artificial cultivation technology is not yet mature, and the growth cycle is long, the yield is low, and it is susceptible to diseases and insect pests, so the cultivation of
Sanghuangporus sanghuang is still difficult. Liquid medium is the preferred method for the large-scale cultivation of fungal mycelium because it enables full contact of the fungal clumps with the medium after inoculation and promotes the immediate absorption of nutrients. The liquid medium also enables rapid release of the fungal metabolites. Liquid culturing is simple and the formulation is flexible and amenable to adjustments. Therefore, liquid culturing is routinely used for large-scale fungal culturing and the extraction of fungal metabolites [
16]. In this study, we used the one-way testing and response surface methodology to optimize the optimal carbon source, nitrogen source, and exogenous growth factors in the liquid-fermentation-media formulation for improving the yield of
S. sanghuang. Furthermore, we performed chemical antioxidant and MTT assays to determine the antioxidant and anticancer activities of the extracellular polysaccharides of
S. sanghuang to provide a theoretical basis for future industrial applications and drug development.
4. Discussion
Sanghuangporus spp. is a rare medicinal fungus. Polysaccharide is an important active substance of its fruiting body and mycelium; it is a type of compound with hypoglycemic, hypolipidemic, antioxidant, anti-tumor, and immunomodulatory activities [
22].
Sanghuangporus spp. is a perennial fungus. Since the growth of wild
S. sanghuang fruiting bodies is limited by natural conditions and has a long growth cycle, its output cannot meet market demand. At present, the cultivation scale of
S. sanghuang is small and unevenly distributed, and the planting technology is immature, making it difficult to guarantee the yield and quality of
S. sanghuang, and it is difficult to increase the added value of
S. sanghuang products [
23]. There is a large talent gap in the resource evaluation, standard formulation, improved variety selection, and technology promotion of
S. sanghuang, which restricts the high-quality development of a
S. sanghuang industry [
23]. Therefore, the artificial cultivation technology of
S. sanghuang is still in the bottleneck period, and liquid fermentation culture of
S. sanghuang has become another research hotspot. Liquid fermentation culture can quickly obtain a large amount of metabolites and can directly extract physiologically active substances for medicinal research. Compared with solid fermentation, liquid fermentation has the advantages of continuous production, low cost, simple operation, short fermentation time, high yield, and high efficiency.
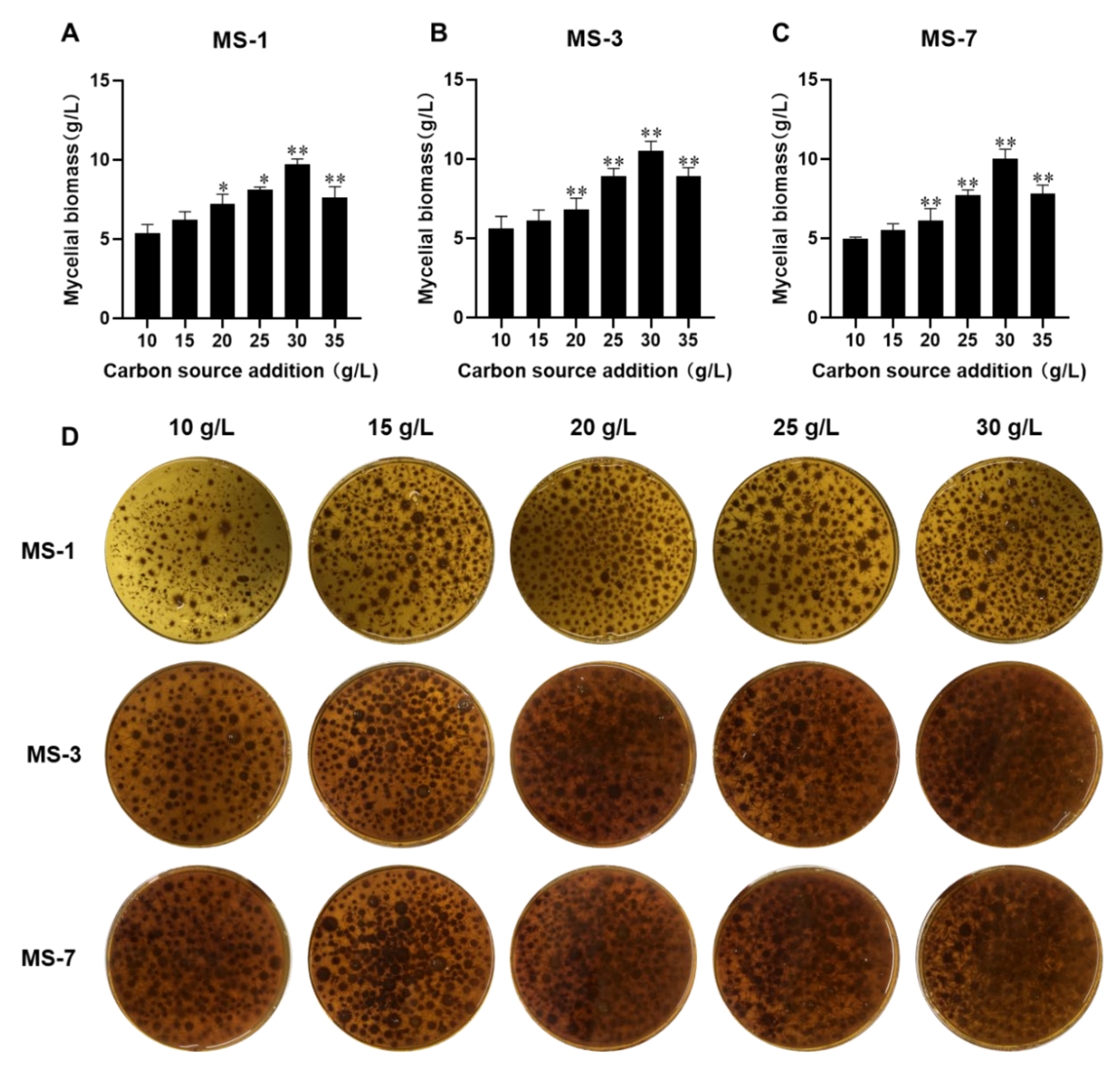
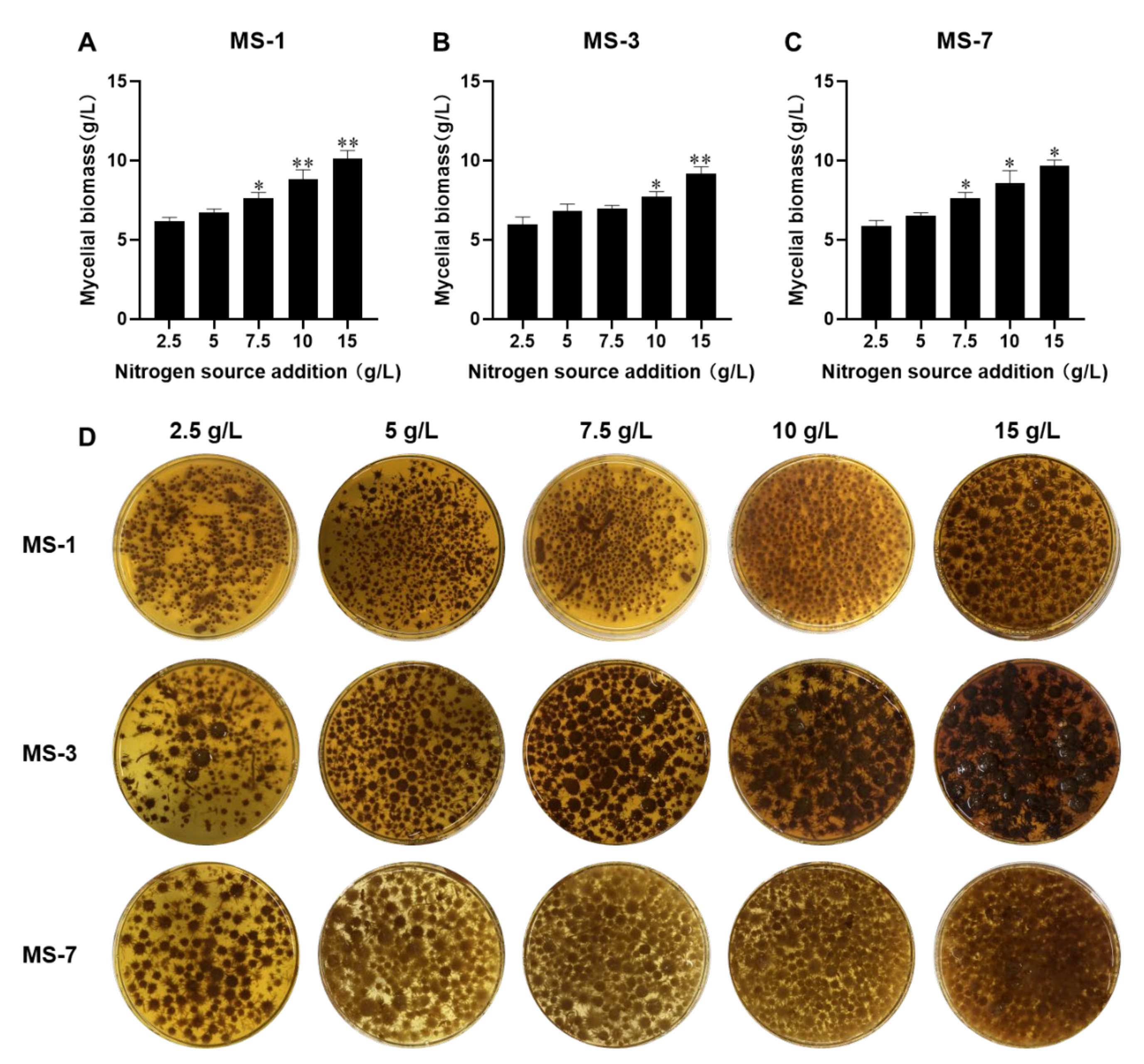
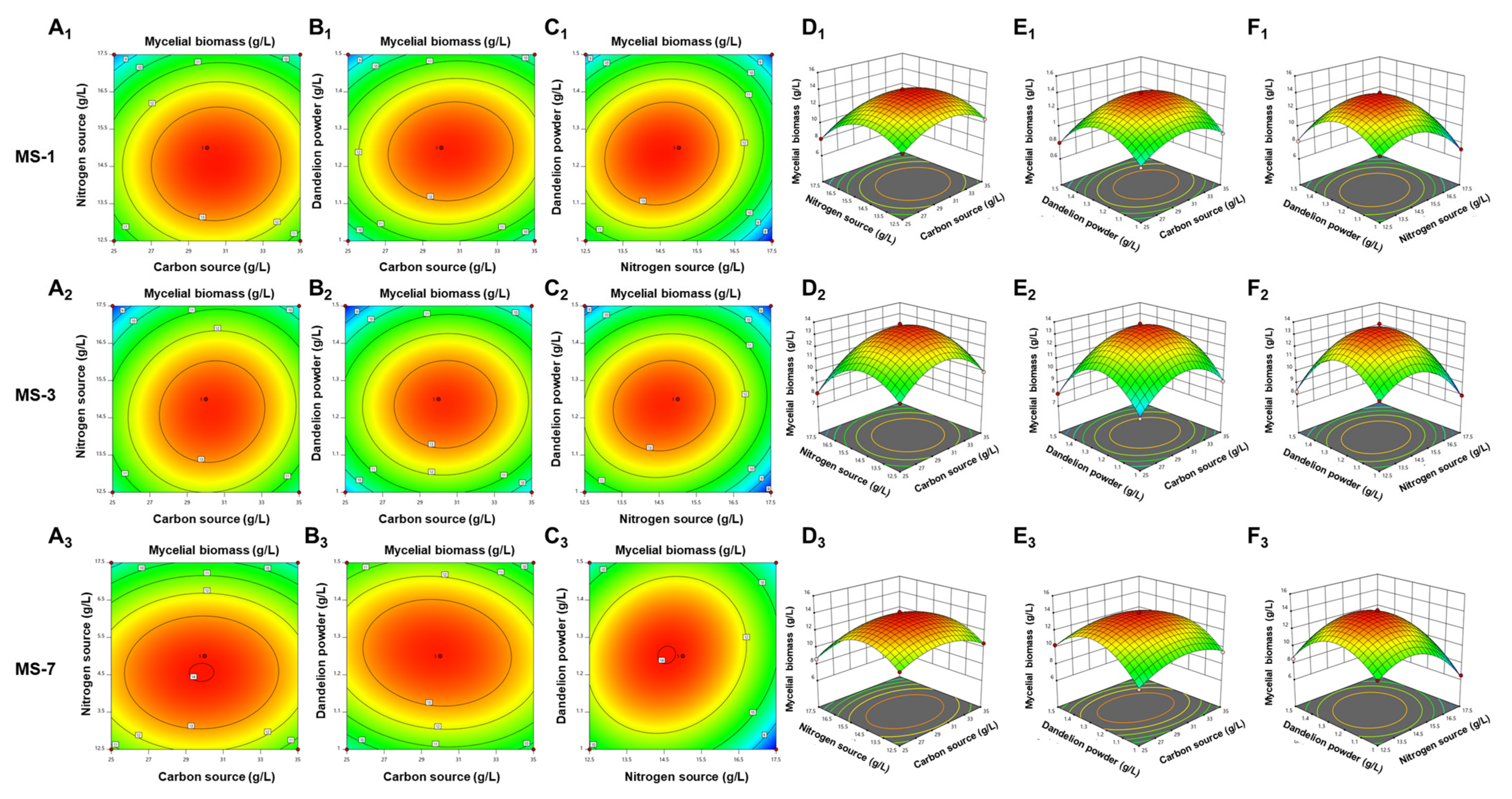
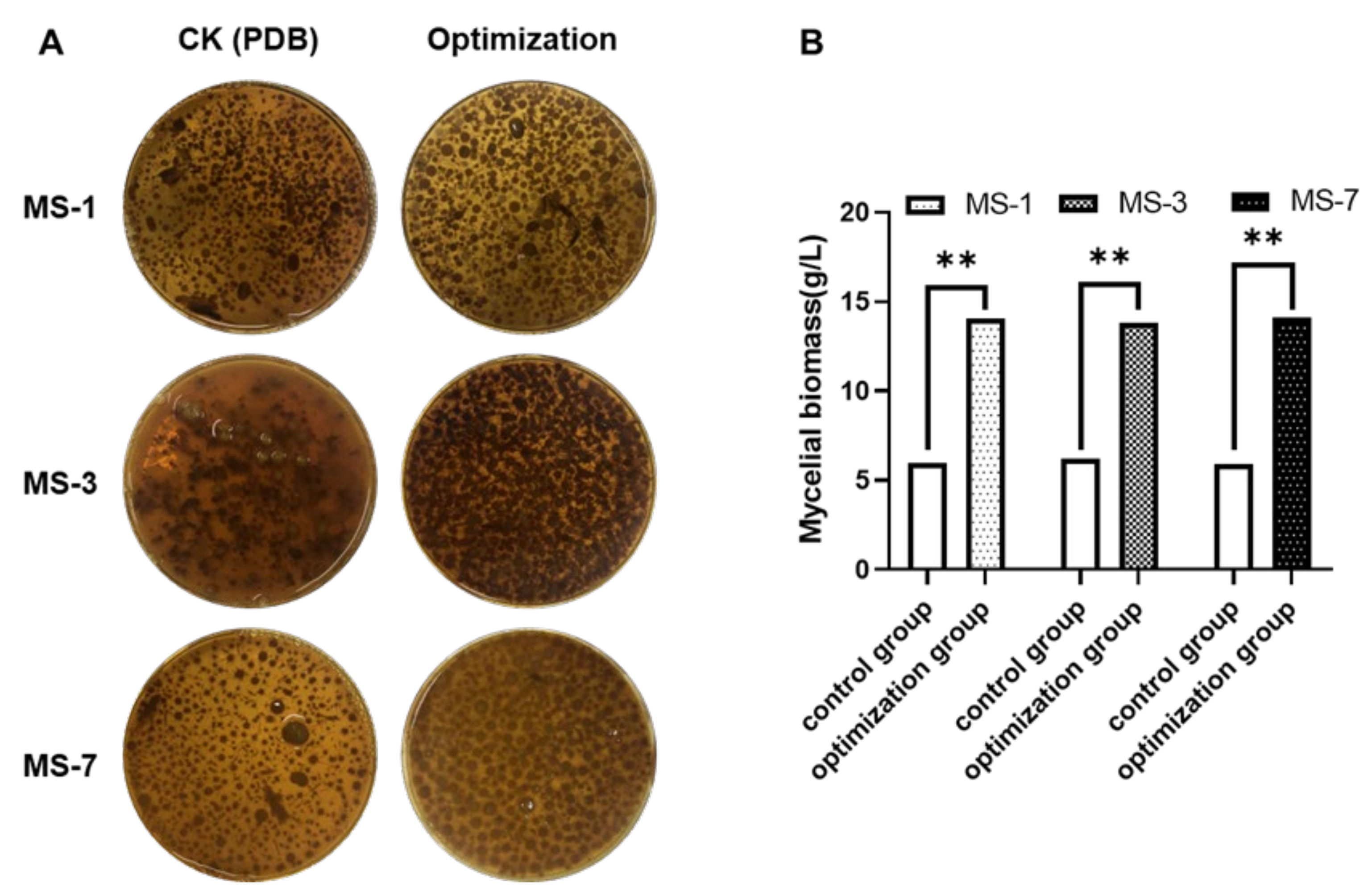
Therefore, development of the optimal liquid-fermentation-media formulation and conditions were necessary for the large-scale production of
S. sanghuang. In this study, we comprehensively analyzed the effects of different carbon and nitrogen sources, as well as exogenous growth factors, to develop the optimal liquid-fermentation-media formulation for maximal mycelial yield. We also performed the response surface method optimization test using a Box–Behnken design to analyze interactions between various factors. Furthermore, we analyzed the antioxidant and anticancer activities of the extracellular polysaccharides from three strains of
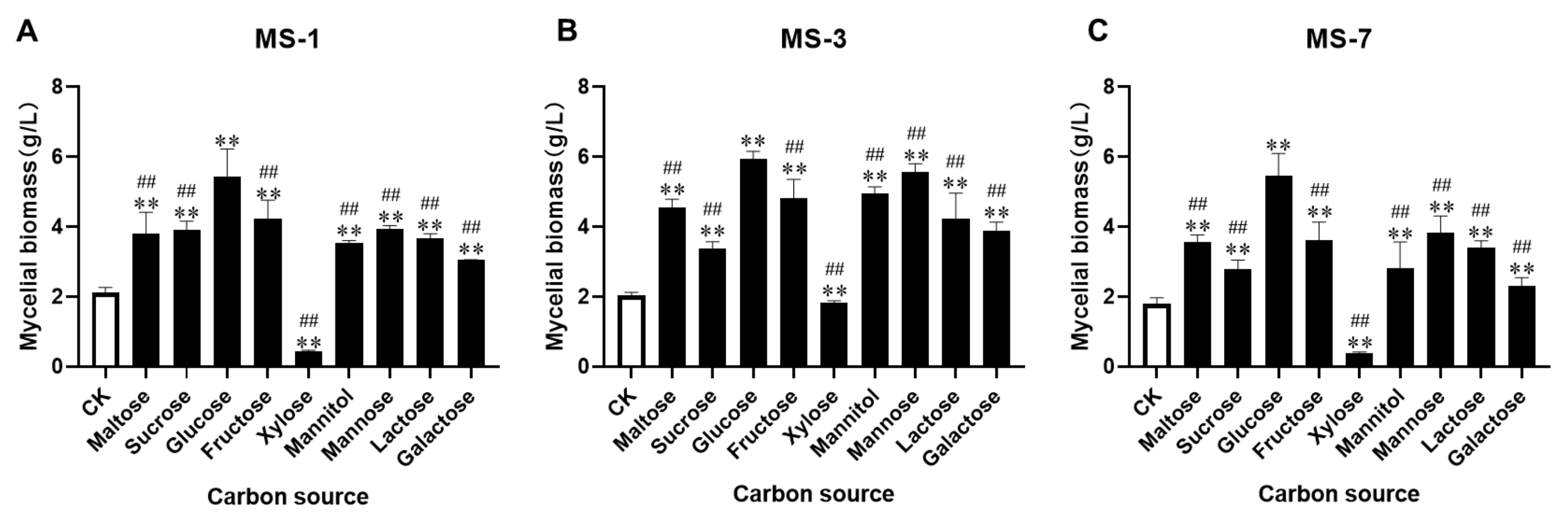
S. sanghuang. Previous studies have shown that, although different strains of
S. sanghuang show similar morphology, their requirements for carbon, nitrogen, and growth factors vary considerably. Our data showed that glucose was the optimal carbon source for the liquid fermentation medium used to grow
S. sanghuang. This was comparable with the composition of the PDA medium that was used to grow
S. Sanghuang in a previous study [
24]. Jiang et al. also reported that glucose was the most significant requirement for the growth of
S. sanghuang [
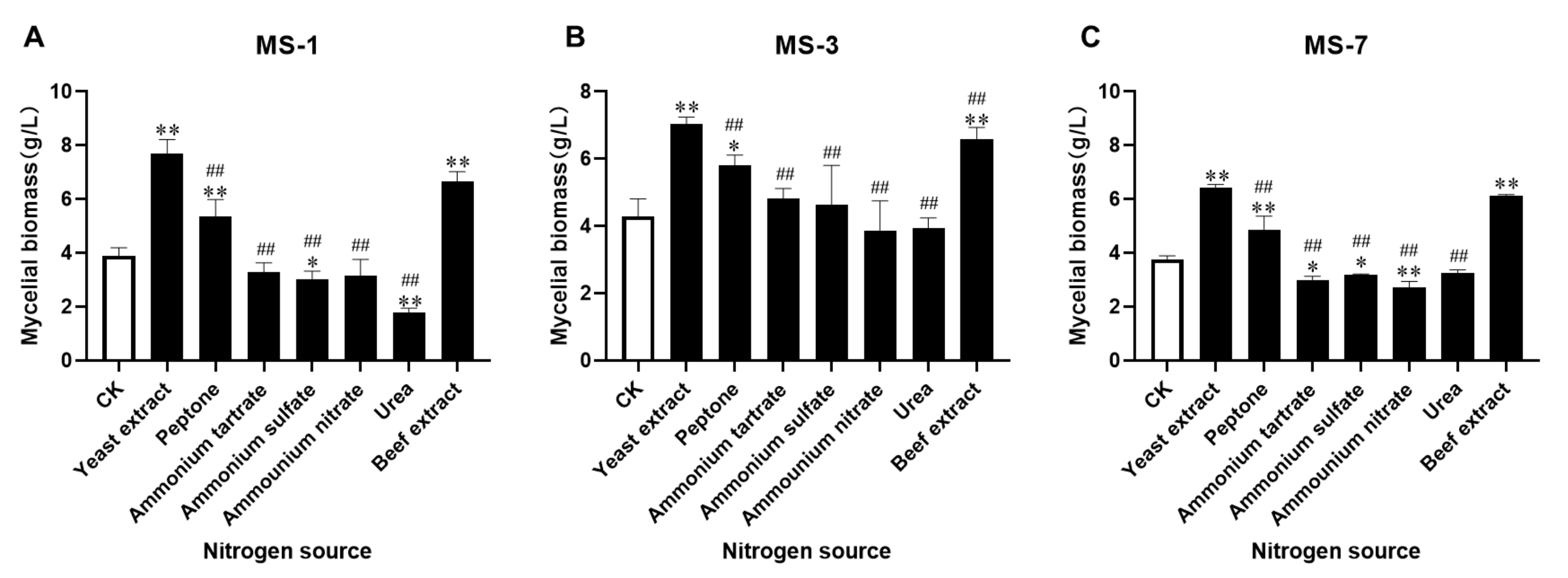
25]. Furthermore, our data showed that yeast extract was the most suitable nitrogen source for growing
S. Sanghuang. Organic nitrogen sources such as yeast extract, beef extract, and peptone were favorable for the optimal growth of
S. sanghuang, but inorganic nitrogen sources such as ammonium tartrate, ammonium sulfate, ammonium nitrate, and urea were unfavorable for the growth of
S. sanghuang. These results were comparable with the findings reported by Cheng et al. [
11]. Since the nutritional conditions significantly alter the growth of
S. sanghuang, a comprehensive study of nutrients and growth factors was necessary to develop an efficient, affordable, and optimal culture medium for obtaining maximal mycelial yield. Many researchers have chosen to add growth factors of natural components to promote the growth or the production of bioactive compounds of
S. sanghuang. For example, based on the study of
Sanghuangporus baumii medium of
Morus alba L., Wang et al. reported that a mulberry bark extract promoted the growth of
S. sanghuang [
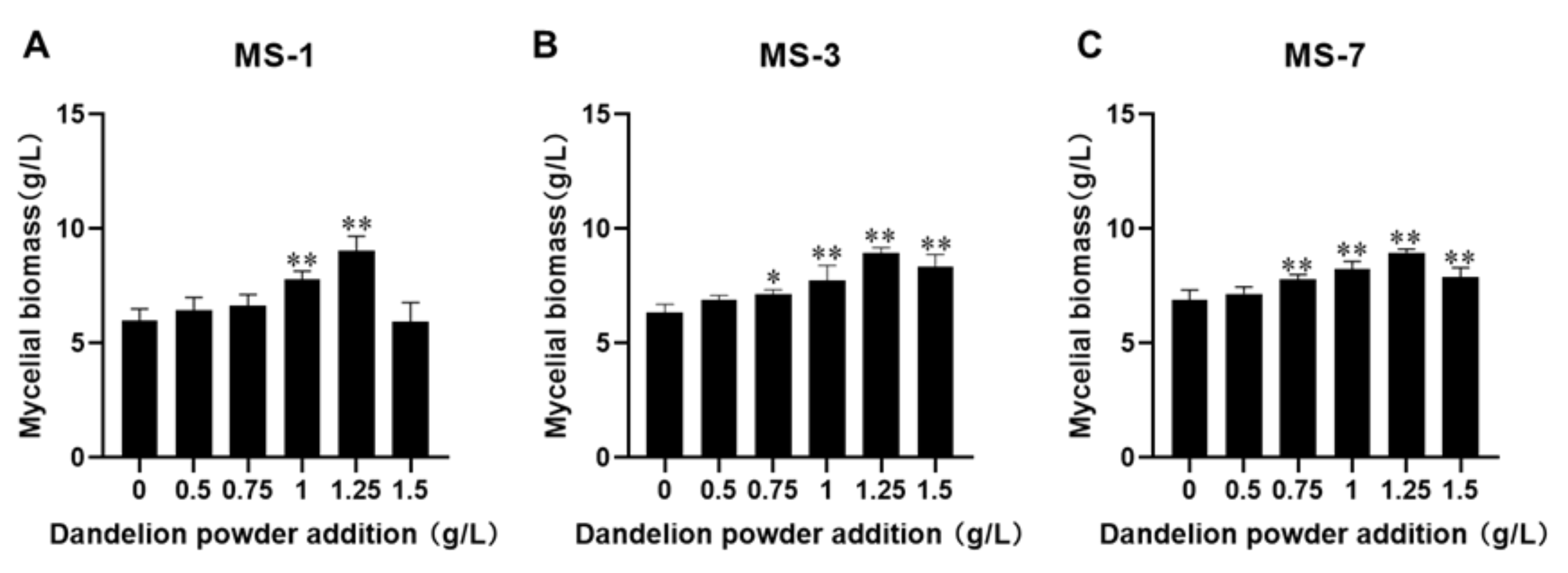
26]. In this study, 1.25 g/L dandelion powder significantly promoted the growth of
S. sanghuang.
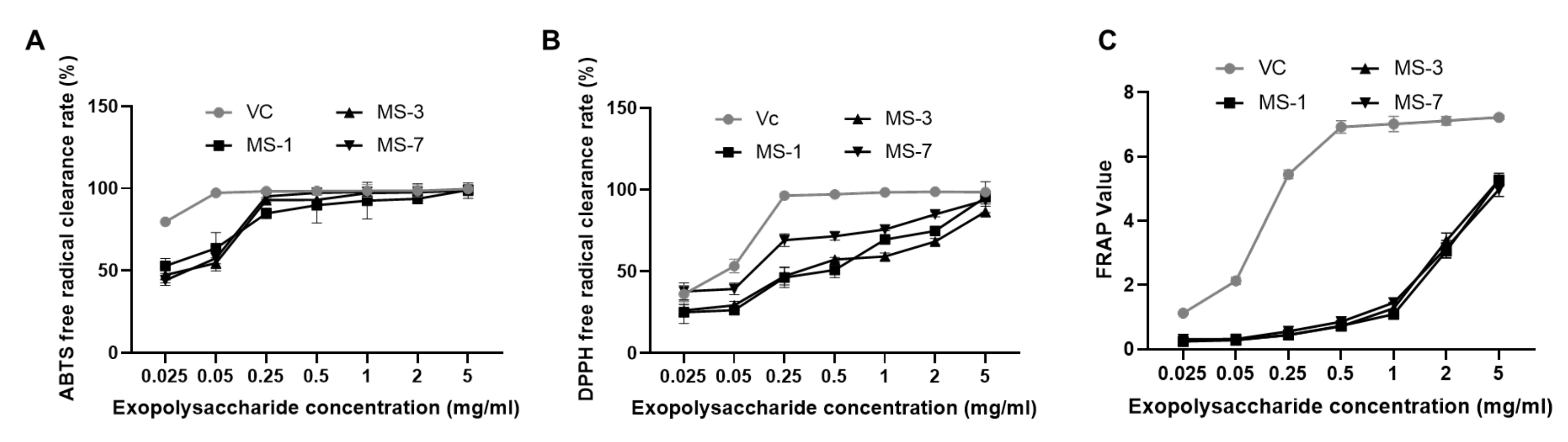
Furthermore, we analyzed the antioxidant and anticancer activities of the extracellular polysaccharides from
S. sanghuang by in vitro experiments. The polysaccharides from
S. sanghuang showed stronger ABTS-radical scavenging ability than DPPH radical scavenging ability. The total antioxidant capacity was also stronger at higher concentrations of the polysaccharides. When the concentration of extracellular polysaccharide is 0.5 mg/mL, the scavenging rate of ABTS free radicals by extracellular polysaccharide can reach 90% and above, which shows that the antioxidant capacity of
S. sanghuang extracellular polysaccharides is substantial. Within a certain concentration range, increasing the polysaccharide concentration can improve the scavenging ability of ABTS and DPPH free radicals. The antioxidant effects of the exopolysaccharide of the
S. sanghuang can provide a theoretical basis for its application in medical dressings, facial cleansers, facial masks, antioxidants, etc.
S. sanghuang also has triterpenoids, flavonoids, and other compounds, but its biological activity needs to be further explored. Lang et al. [
27] found that
Phellinus baumii flavonoids have strong reducing power and strong scavenging ability for DPPH free radicals and hydroxyl free radicals, and the antioxidant capacity is dose-dependent on the flavonoid concentration. Li [
28] reported that the flavonoid compounds of sanghuang had strong DPPH free radical scavenging ability and iron reduction ability.
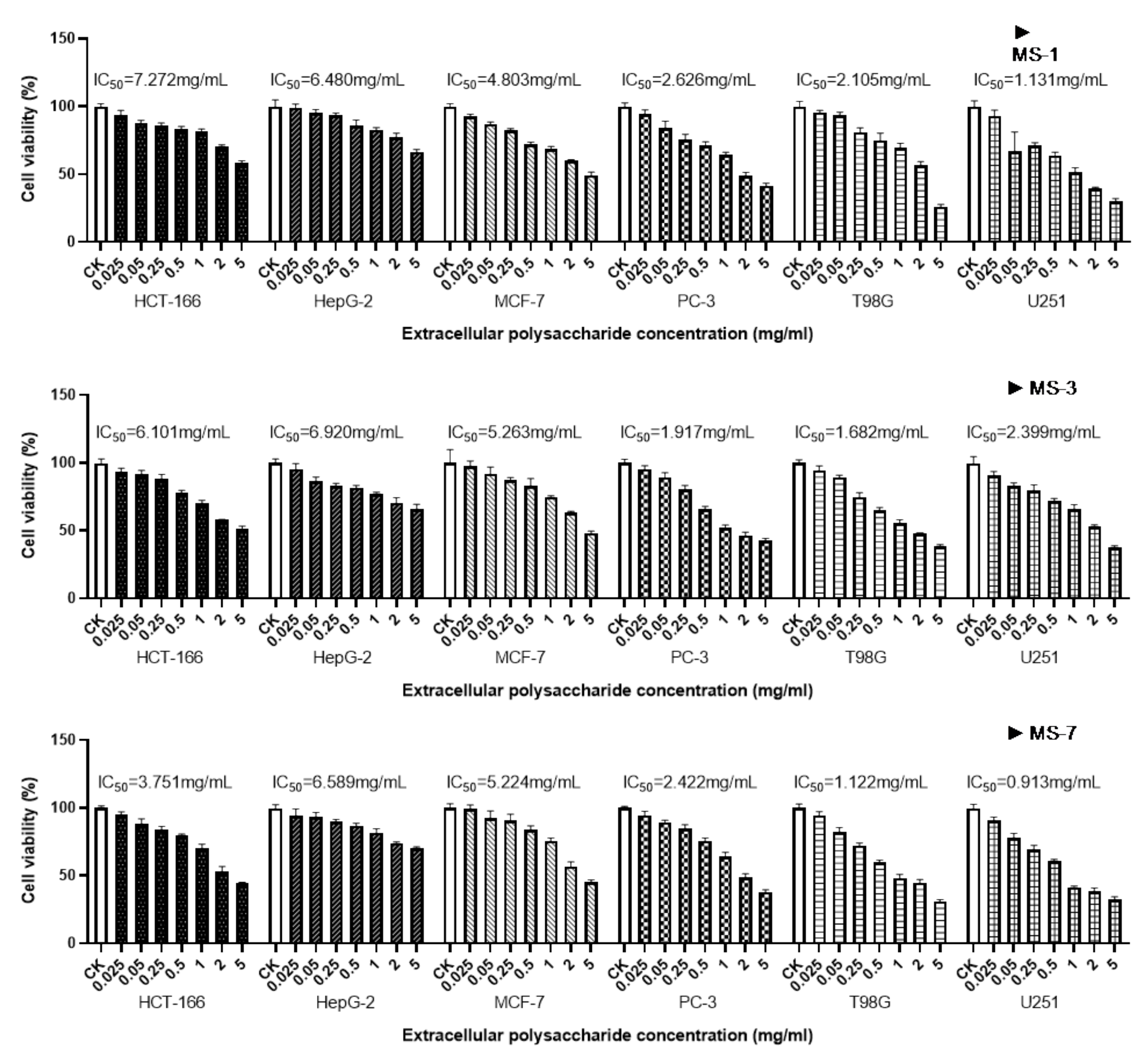
MTT assay results showed that the polysaccharides from
S. sanghuang significantly decreased the survival rate of U251 and T98G cells (human glioma cells), and PC-3 cells (human prostate cancer cells). The anticancer effects of the polysaccharides from
S. sanghuang were poorer on the remaining three types of cancer cells. Therefore, in future studies, further investigations are necessary to determine the mechanisms by which the extracellular polysaccharides of
S. sanghuang inhibit the growth and survival of cancer cells. The unique anticancer effects of sanghuang polysaccharides has made this a hot topic in functional food research at home and abroad. Liu et al. [
14] have shown that
Phellinus baumii polysaccharides can inhibit the proliferation of Hela and SGC-7901 cells by hindering cell division. How
S. sanghuang extracellular polysaccharides exert anticancer effects remains to be further studied in later trials. Guo’s [
29] research found that
S. sanghuang triterpenoids have inhibitory effects on PC-3, HepG-2, MDA-MB-231, HCT-116, and other cancer cells, and that they have a certain anticancer potential. In the future, we may measure the metabolic profile of
S. sanghuang and analysis its correlation with biological activity.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}