3.2. Green Leaf Volatile Evolution
GLVs are generated from unsaturated fatty acids through enzymatic processes involving lipoxygenases, hydroperoxide lyases, as well as alcohol dehydrogenases [
24]. GLVs in grapes are responsible for citrus, leafy, and green sensory attributes, and they play a significant role in the fragrance profile of grapes [
25]. We studied the development of total GLVs in six grape types, as shown in
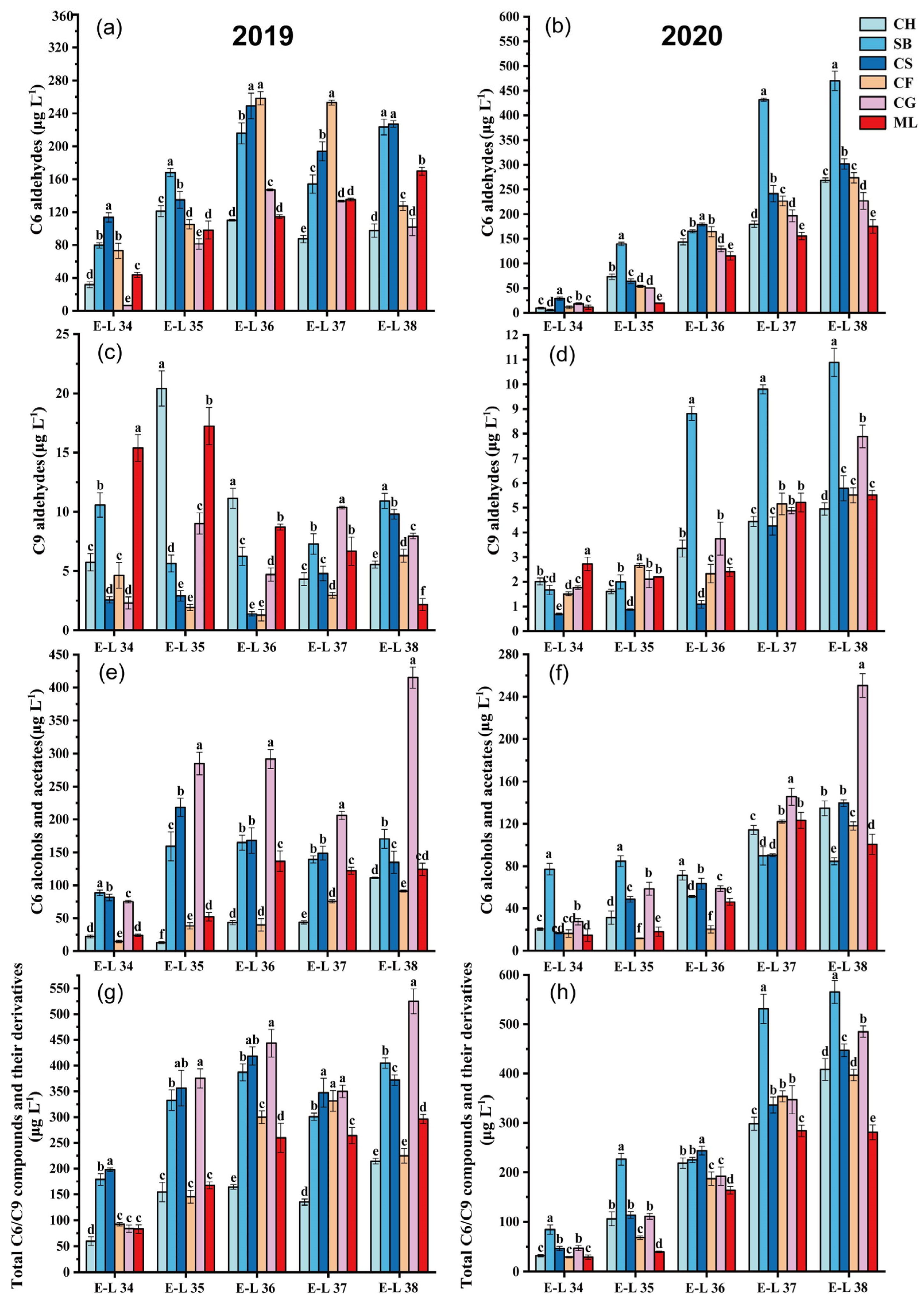
Figure 1a–h.
In 2019, during the grape maturation process, all the varieties exhibited lower C6 aldehyde concentration at E-L 34 (
Figure 1a). The C6 aldehyde concentration in the six grape types at harvest varied from 97.59 to 227.11 μg L
−1. Cabernet Sauvignon and Sauvignon Blanc had significantly greater total C6 aldehyde levels at harvest E-L38 than the other varieties. The C6 aldehyde growth pattern in the 2020 grapes resembled that of 2019, showing lower concentrations in the early stages of development and steady accumulation with maturation (
Figure 1b). Compared with those of the other varieties, the overall C6 aldehyde levels in Sauvignon Blanc and Cabernet Sauvignon significantly increased throughout development. The C6 aldehyde levels in the six grape types at harvest in 2020 varied between 174.93 and 469.86 μg L
−1. In line with our results, Lu et al. [
26] found that C6 aldehyde concentrations were different in summer and winter, suggesting that the yearly variation in C6 aldehyde levels in grapes might be due to seasonal weather differences, necessitating additional research.
In 2019, the trends in total C6 alcohols and acetates varied significantly among different types (
Figure 1c). The concentrations of C6 alcohol and acetate in grapes rose by factors of 4.04—(Chardonnay), 0.93—(Sauvignon Blanc), 0.65—(Cabernet Sauvignon), 5.25—(Cabernet Franc), 4.52—(Cabernet Gernischt), and 4.18—fold (Marselan). In 2020, Sauvignon Blanc grapes exhibited the greatest levels of total C6 alcohol and acetate concentrations at E-L 34 and E-L 35. The highest levels of total C6 alcohols and ethyl esters at harvest were discovered in Cabernet Gernischt grapes, with Cabernet Sauvignon and Merlot varietals displaying lower levels (
Figure 1d). The growth trends of total C6 alcohol and acetate varied within the six grape types in 2020 compared to 2019. Except for those of Chardonnay and Cabernet Franc, the total C6 alcohol and acetate contents of the 2020 grapes were significantly lower than those of the 2019 harvest, which was possibly related to climate differences. A warmer climate facilitates the transformation of free C6 aldehydes into C6 alcohols, which generally have higher herbaceous odor thresholds (negative aroma) than the corresponding aldehydes [
26]. Additionally, alcohols are more likely to form esters, thereby minimizing the herbaceous characteristics of aldehydes and maximizing the fruity characteristics of esters [
25].
C9 compounds in grapes primarily exist in aldehyde form, such as (E)-2-nonenal, (E, E)-2,6-nonadienal and nonanal. C9 aldehydes, even in low quantities, have low thresholds that enable them to impart citrus and cucumber scents to grapes [
13]. In 2019, Marselan at E-L 34 exhibited the highest C9 aldehyde level compared to all other grape types (
Figure 1e). During the mature stage (E-L 38), Sauvignon Blanc exhibited the highest concentration of C9 aldehyde, with Cabernet Sauvignon following closely behind. The overall accumulation trend of C9 aldehydes in 2020 was similar to that in the 2019 vintage, with six varieties gradually accumulating C9 aldehydes or exhibiting stabilization of C9 aldehyde levels with growth and development (
Figure 1f). A comparison with existing literature reveals that the presence and concentration of C9 aldehydes in grapes are influenced by a multitude of factors, including genetic varietal characteristics, viticultural practices, and, notably, climatic conditions [
27,
28]. Studies have highlighted the sensitivity of C9 aldehyde synthesis to environmental factors, suggesting that variations in temperature, sunlight exposure, and rainfall can significantly affect their accumulation in grapes [
29]. The meteorological data presented for the vintages 2019 and 2020 (
Table S1) provide critical insights into the climatic backdrop against which these aromatic compounds developed. The 2019 vintage, characterized by higher mean maximum temperatures and lower relative humidity during critical growth phases, might have facilitated the enhanced synthesis or preservation of C9 aldehydes in Marselan and, subsequently, Sauvignon Blanc. Conversely, the 2020 vintage, with its increased rainfall and slightly lower temperatures, suggests a climatic influence on the slightly different accumulation pattern observed.
With respect to the total GLV contents in 2019, except for Cabernet Franc, in which the GLV contents slightly declined at maturity, all the other varieties exhibited maximum GLV levels at maturity (
Figure 1g). The GLV contents of all the varieties at maturity ranged from 214.86 to 524.81 μg L
−1. Cabernet Gernischt and Cabernet Sauvignon had higher GLV contents during growth and development. At harvest, Cabernet Gernischt exhibited the greatest GLV contents. The GLV concentration among all berries at harvest in the 2020 vintage varied between 281.14 and 565.31 μg L
−1 (
Figure 1h). Sauvignon Blanc and Cabernet Gernischt had higher total GLV contents at harvest, while Marselan and Chardonnay had lower GLV contents at harvest.
As all grape samples originated from the same vineyard, differences in external influences were reduced when comparing different grape kinds and vintages. These results indicate that variations in GLV contents among different varieties might be due to genetic variations affecting LOX-HPL gene expression and the influence of different environmental conditions [
13] (Wang et al., 2019). Consistent with our findings, Yue et al. [
12] found that the vintage had an impact on the variation patterns of GLVs. Seasonal variations in the microclimate, such as sunlight and rainfall, could impact gene expression in the GLVs synthesis pathway, thereby affecting the function of enzymes associated with GLVs biosynthesis [
8,
18]. The cultivar significantly impacts the aroma of grapes. Li et al. [
1] noted substantial variations in GLV levels between Marselan and Merlot scion–rootstock grapes. There is insufficient research on the alterations in GLV compounds associated with green aromas as different wine grape varieties ripen. Our study fills this gap by demonstrating that GLV volatiles not only affect the aromatic characteristics of grapes but are also a component of varietal typicity.
3.3. Analysis of Green Leaf Volatile Profiles in Grapes as They Mature
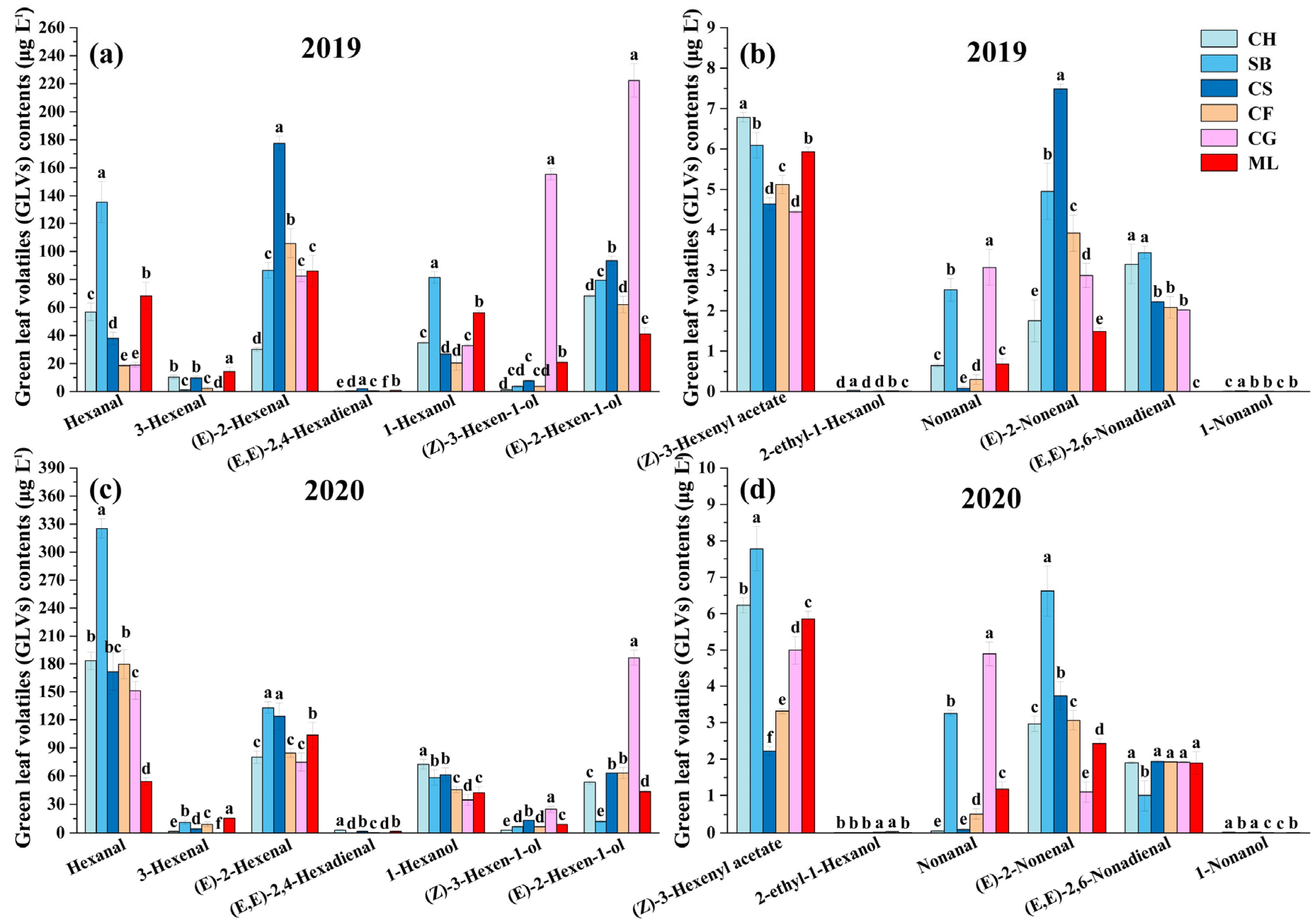
Brix is widely regarded as a reliable maturity indicator. Higher Brix levels typically signify greater ripeness, which is associated with not only increased sugar concentration but also alterations in the grape’s aromatic compound profile. Among the six grape varieties harvested, 13 GLVs were detected (
Figure 2). The main GLVs in all mature 2019 berries were (E)-2-hexenal and (E)-2-hexen-1-ol. (E)-2-hexenal makes up 14.01–47.71% of the total GLVs composition, whereas (E)-2-hexen-1-ol accounts for 13.93–42.34% (
Figure 2a,b). Hexanal and (E)-2-hexenal constituted the predominant components in mature grapes in 2020, with hexanal making up 31.26–57.51% of the total GLV contents and (E)-2-hexenal accounting for 15.42–27.73% (
Figure 2c,d). This suggests that, despite the varietal differences in sugar accumulation, as reflected by Brix levels, certain characteristic compounds were universally present in grape varieties, contributing to the baseline aromatic complexity of wines.
Regarding C6 alcohols, at harvest, all grapes in both vintages contained 1-hexanol, (E)-2-hexen-1-ol, 2-ethyl-1-hexanol, and (Z)-3-hexen-1-ol. Cabernet Gernischt had the greatest C6 alcohol concentration in 2019 and 2020 among the six mature varieties, while Cabernet Franc had the lowest level in 2019 and Sauvignon Blanc in 2020. 1-hexanol enhances the perception of fruity, floral, and green notes in grapes [
30]. In both vintages, Sauvignon Blanc and Cabernet Sauvignon exhibited higher 1-hexanol concentrations. (E)-2-hexen-1-ol imparts fruity aromas to grapes, produced through the catalytic action of ADH genes from (E)-2-hexenal [
31]. According to the C6 alcohol profile, (E)-2-hexen-1-ol made up 14.16–74.43% among all grapes in both years. Cabernet Gernischt had the greatest concentrations of (E)-2-hexen-1-ol among all the types, with 222.21 μg L
−1 in 2019 and 186.46 μg L
−1 in 2020, similar to its overall C6 alcohol values.
Four distinct C6 aldehydes were identified in ripe berries. (E)-2-hexenal and hexanal, the primary C6 aldehydes, were found in all grape types at harvest, in line with prior research on fresh grape varietals [
5,
32]. (E)-2-hexenal concentrations in mature berries varied between 30.05 and 177.74 μg L
−1 over the two years. All the mature grapes had higher hexanal contents in 2020 than in 2019. In particular, mature Sauvignon Blanc had the greatest hexanal levels in 2019 and 2020. Consistent with our findings, (E)-2-hexenal and hexanal were the predominant C6 volatiles in the six wild grape species [
9,
25]. These compounds might impart a rich grassy and tomato flavor to grapes [
33]. Wang et al. [
11] also suggested that hexanal production is influenced by the growing environment and variety of grape plants.
The GLV profiles showed (Z)-3-hexenyl acetate as the only C6 ester. In 2019, the (Z)-3-hexenyl acetate content was highest in Chardonnay, and, in 2020, it was highest in Sauvignon Blanc. Some studies showed that (Z)-3-hexenyl had a minor impact on grape fragrance [
23].
Four distinct C9 compounds were identified in ripe berries. Nonanal, (E)-2-nonenal, 1-nonanol, and (E, E)-2,6-nonadienal were detected in all grapes. Minimal quantities of 1-nonanol found in wine grapes (0.00–0.02 μg L
−1) were consistent with a previous study [
11]. In future studies, implementing comprehensive two-dimensional gas chromatography (2D GC) analysis could enhance detection accuracy and reduce detection thresholds for this set of chemicals [
18,
34]. The levels of (E)-2-nonenal and (E, E)-2,6-nonadienal were between 1.10–7.49 μg L
−1 and 0–3.44 μg L
−1, accordingly, beyond their smell thresholds of 0.08 and 0.02 μg L
−1 [
13,
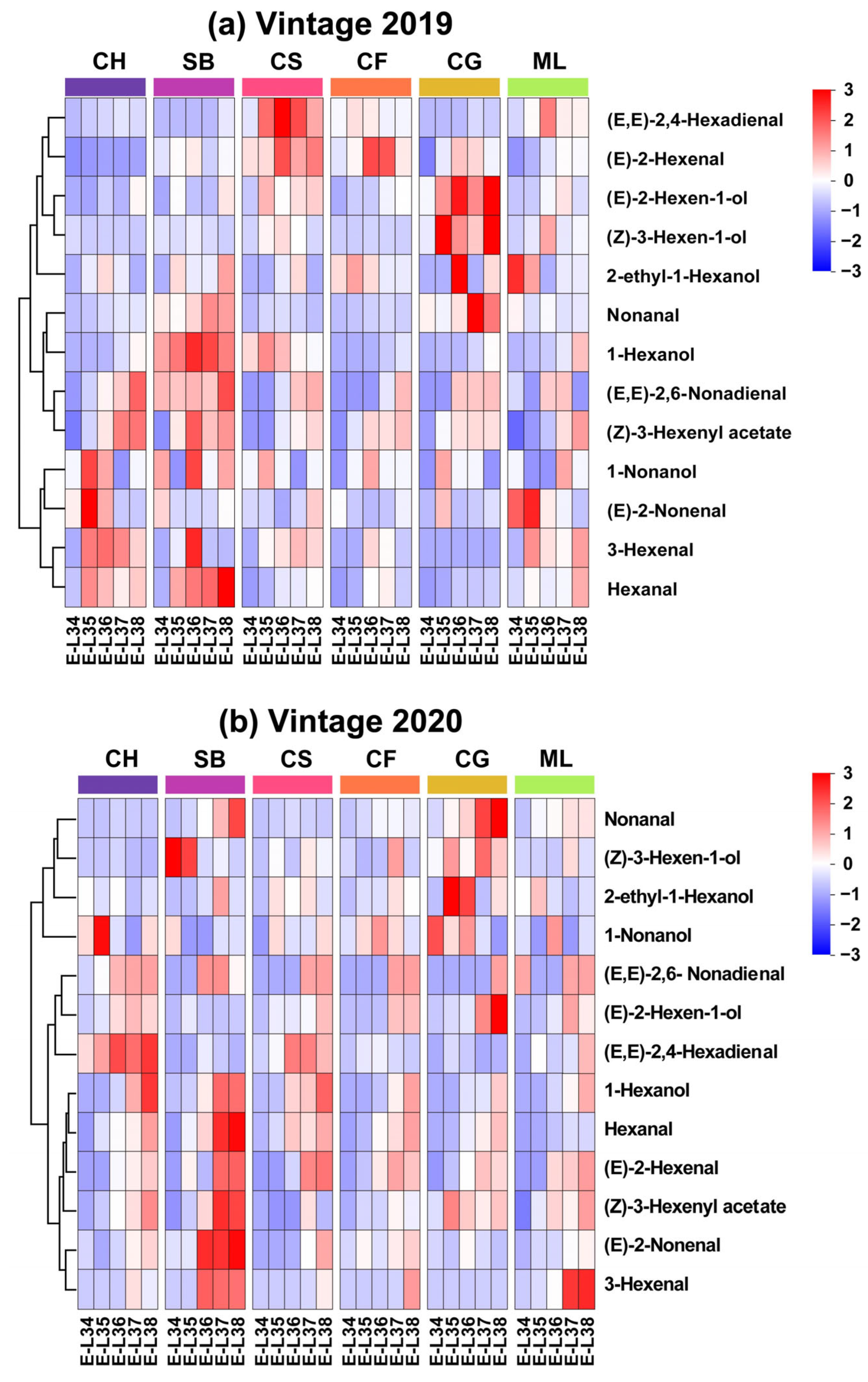
35]. The evolution of the GLV profiles in 2019 and 2020, respectively, are shown in
Figure 3a,b. Our findings suggest that the development of the GLV profile varies not only among grape varieties but also among different vintages of the same variety suggested that this variation might be attributed to genetic factors and environmental influences. Below, we describe the variations in the GLV profiles of the six grape varietals as they progress through the stages of berry maturation.
The heatmap of concentrations reveals that GLV profiles are grouped into three distinct clusters. In 2019, 3-hexenal and hexanal exhibited a trend of increasing and then decreasing in Cabernet Franc and Cabernet Gernischt. A similar evolutionary pattern was observed in the remaining four grape varieties, with higher concentrations during the ripening period and lower concentrations at harvest (
Figure 3a). 1-hexanol, (E, E)-2,6-nonadienal and (Z)-3-hexenyl acetate showed consistent patterns of rise in all six types, reaching their highest levels at the mature stage as they grew and developed. Some GLVs existed only at specific stages in typical varieties. For instance, the levels of (E)-2-hexenal and (E, E)-2,4-hexadienal are lower across the varieties at E-L 34. Regarding (E)-2-Hexen-1-ol and (Z)-3-Hexen-1-ol, Cabernet Gernischt exhibited the highest abundance during berry development, while Chardonnay had the lowest. In 2020, hexanal, 1-hexanol, (Z)-3-hexenyl acetate, and 3-hexenal across the six varieties matured into the same category, gradually increasing in content with growth and development, and peaking near maturity across the examined varieties (
Figure 3b). Similar to the year 2019, the contents of 1-hexanol, hexanal and (Z)-3-hexenyl acetate increased with growth and development in 2020, peaking at the mature stage. However, (E)-2-nonenal and 3-hexenal had inconsistent evolution patterns over the two years.
The differences in the accumulation patterns of C6 and C9 aldehydes among different grape varieties highlight the complex interplay of genetic and environmental factors that influence grape aroma characteristics. As reported by Wang et al. [
30], the total content of C6 aldehydes was greater in 2020, indicating a strong environmental impact, possibly related to climatic conditions. This observation is crucial for grape growers and winemakers, as it implies that annual climatic variations can significantly affect the aromatic compounds in grapes, thus influencing the quality of the wines. Furthermore, the variation in the peak accumulation stages of different grape varieties underscores the importance of determining the exact timing of grape harvesting to optimize the aroma profile. This knowledge can aid in developing targeted viticultural practices to enhance the aromatic characteristics of wines.
3.4. GLV Evolution Pattern Recognition
GLVs are mostly produced through the LOX-HPL pathway, and their production is influenced by grape farming and environmental conditions [
6]. Genetic profile plays a major role in regulating GLV synthesis [
36]. Prior research has shown that C6 volatile components differ among various fresh grape cultivars [
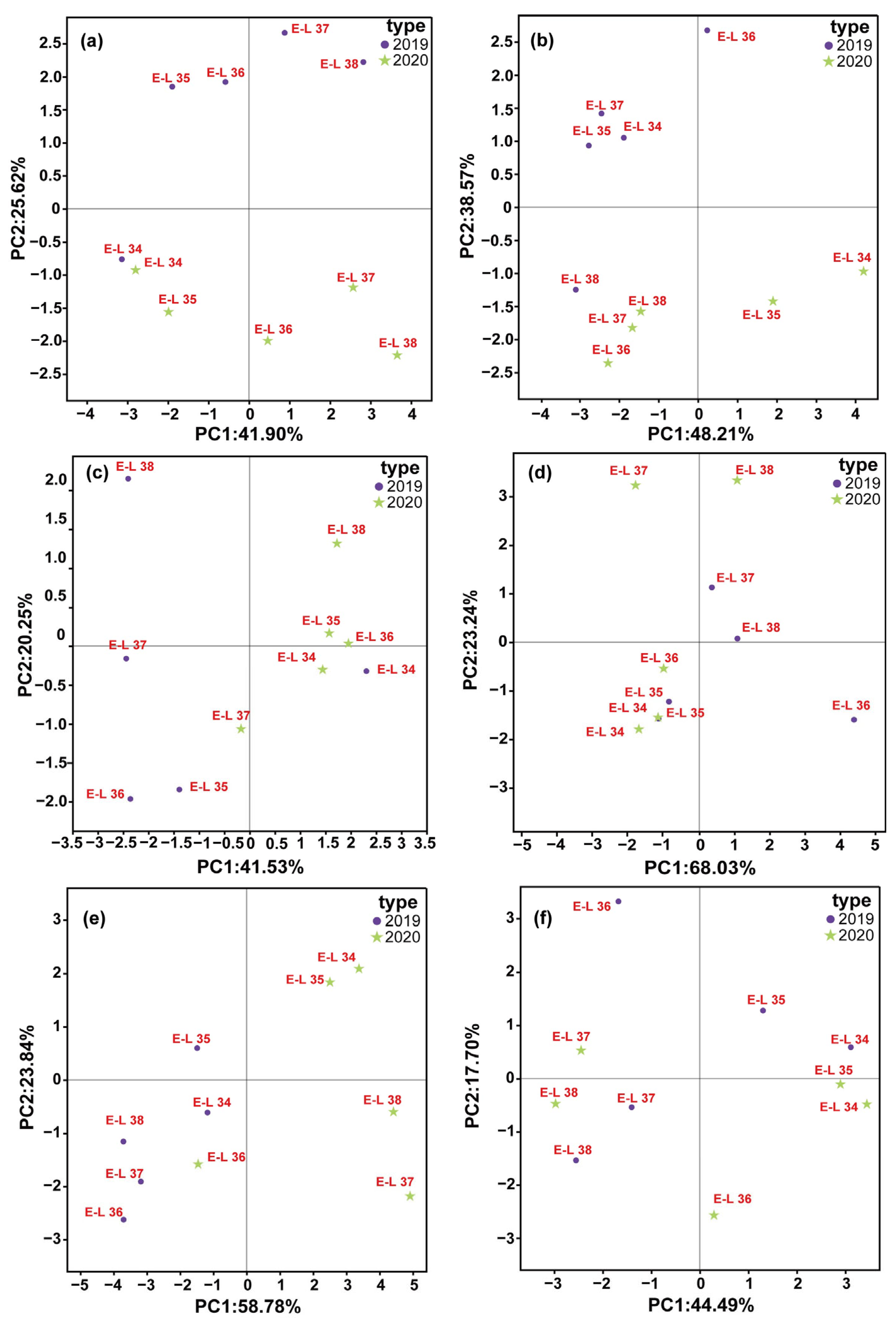
37]. There is limited research on the development of GLV profiles in a diverse range of wine grapes in the Loess Plateau region. Therefore, principal component analysis (PCA) was applied to the GLV profiles of six wine grape varietals during a two-year period, as shown in
Figure 4.
The synthesis of GLVs is controlled by environmental and viticultural conditions, as well as the genetic traits of the grape variety [
9]. PCA was utilized to examine the variations in GLV components across different cultivars during fruit development (
Figure 4a–f). The first two functions accounted for over 60.00% of the variance across all the types. The PCA results indicated that the GLV components of the six varieties exhibited noticeable differences at various growth stages. Furthermore, analysis of berry GLVs profiles showed significant differences between the E-L34 and E-L38 period across the six varieties. Previous studies on fresh cultivation varieties also indicated that grapes have distinctive C6 volatile contents at harvest, which aligns with our findings [
38].
Building upon the PCA findings, we subsequently performed a Partial Least Squares Discriminant Analysis (PLS-DA) to more effectively discern the variations in GLV profiles during the maturation process of grape berries across two vintages, 2019 and 2020 (
Figure S1a,b). In both vintages, the first principal component (PC1) and the second principal component (PC2) represent substantial proportions of the variation, with PC1 accounting for 20.4% in 2019 and 33.2% in 2020, while PC2 captures 8.6% and 15.7%, respectively. The score plots illustrate a distinct segregation of samples along PC1, indicating a pronounced variance in GLV profiles as a function of the maturation stage.
To further investigate the impact of specific GLVs on the separation observed in the PLS-DA, Variable Importance in Projection (VIP) scores derived from a PLS-DA were utilized (
Figure S1c,d). Compounds with VIP scores greater than 1 are considered significant contributors to the model. In 2019, 1-hexanol, hexanal, (E, E)-2,6-nonadienal, and 3-hexenal were identified as significant due to their VIP scores surpassing this threshold. Similarly, in 2020, 3-hexenal, (E)-2-nonenal, 1-nonanol, and (E)-2-hexen-1-ol were distinguished for their substantial contributions, as reflected by the VIP scores above. Notably, 3-hexenal exhibited high VIP scores in both years, highlighting their influence on the grape variety classification. Such analysis is crucial for identifying biochemical markers impacting grape quality and wine flavor.
3.5. Aroma Activity Analysis
Odor activity value (OAV) was calculated by assessing the ratio of the compound concentration to its odor threshold in water [
8]. Components with an OAV of more than 1 made a substantial contribution to the grape scent, as detailed in
Tables S3–S8. Among all the mature berries from 2019 and 2020, there were nearly 7 and 8 GLV compounds, respectively, with an OAV > 1. Out of the six fully developed grape types assessed in both years, three odor-active GLVs with OAV exceeding 30 were identified as 3-hexenal, (E)-2-nonenal, and (E, E)-2,6-nonadienal, which are likely to play a substantial role in grape scent.
In 2019, Chardonnay at E-L 35 had the highest (E)-2-nonenal OAV, followed by Marselan. Sauvignon Blanc had a greater (E, E)-2,6-nonadienal OAV at harvest, with mature Chardonnay and Cabernet Sauvignon berries having (E, E)-2,6-nonadienal OAVs of 157.50 and 111.12, respectively. The compound (E, E)-2,6-nonadienal had the highest OAV among all the mature berries in 2020, followed by (E)-2-nonenal and 3-hexenal. Mature Cabernet Sauvignon had the highest (E, E)-2,6-nonadienal OAV, with mature Sauvignon Blanc having higher (E)-2-nonenal and hexanal OAVs, which likely had significant impact on the creation of aroma.
Descriptors are commonly employed to describe volatile substances in odor compounds. Aroma compound descriptors are classified into six kinds: sweet, fruity, green, citrus, chemical, and floral [
39]. The aroma profiles of the six grape types differed, with mature Sauvignon Blanc berries exhibiting the most pronounced green aroma due to elevated levels of hexanal. The odor Cabernet Sauvignon, Cabernet Gernischt, and Cabernet Franc peaked at maturity. The aroma profile of the berries changed as they matured, affected by the changing seasons. In 2020, green notes were more pronounced than other odors in all six grape varieties. The aroma characteristics of the six grape varieties generally increased gradually with growth and development. During harvest, Sauvignon Blanc grapes exhibited the strongest green aroma.
This research contributes to a deeper understanding of the chemical basis of grape aroma and flavor, which is crucial in the field of oenology. Furthermore, investigating the effects of these compounds on the final product quality will be valuable. Although our study provides significant insights, it is limited to a few grape varieties. Extending this research to a broader range of varieties could provide a more comprehensive understanding of the variability in aroma compounds.
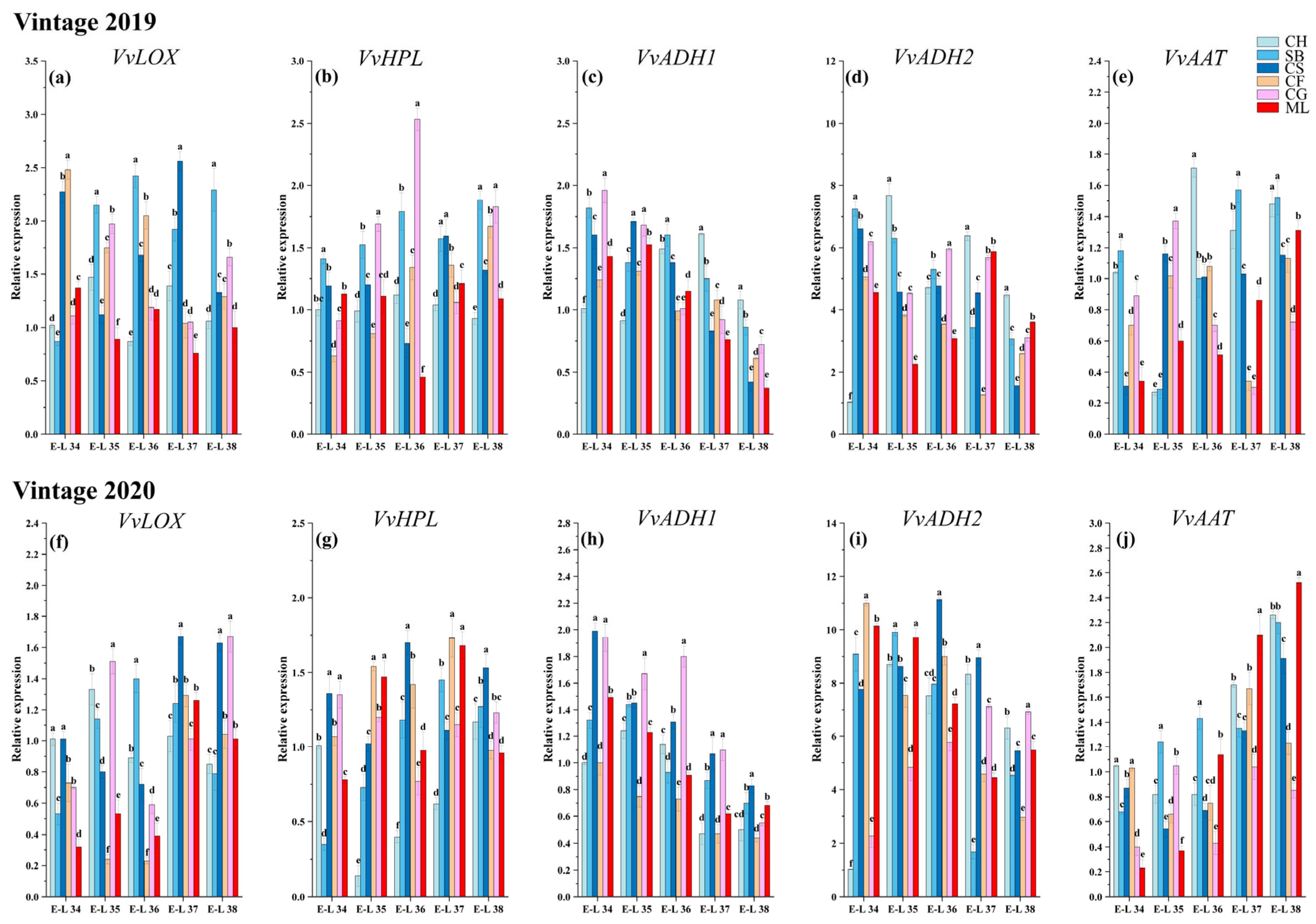
3.6. Expression of Genes in LOX-HPL Pathway
Considering the discernible variation in the concentrations of most GLV compounds across different sampling times and grape varieties, coupled with the fact that C6/C9 volatile compounds are directly synthesized via the LOX-HPL pathway, our study concentrated on analyzing genes that encode the enzymes functional in the LOX, HPL, ADH, and AAT within this pathway. This analysis was performed using RT-qPCR. The levels of gene expression related to the accumulation of GLVs are shown in
Figure 5.
Lipoxygenase (LOX) catalyzes the synthesis of hydroperoxides, which are direct precursors for aldehyde synthesis [
16]. The expression patterns of the
VvLOX genes varied among the six types during development (
Figure 5a,f). In this study, the activity of the
VvLOX gene exhibited fluctuating trends throughout the growth and development stages of six grape varieties in 2019, while showing a notably upward trend overall in 2020. This indicates that the varieties’ responses to environmental or developmental factors may vary between years, suggesting a nuanced interaction between genetic expression and external conditions. The study found that the activity of the
VvLOX gene gradually increased during the growth and development of the six grape varieties, especially in 2020. In 2019, the highest expression of the
VvLOX genes was detected in Cabernet Franc and Cabernet Sauvignon during E-L 34. In Sauvignon Blanc, Cabernet Sauvignon, and Cabernet Gernischt, the expression peaked during the ripening stages. This discovery implied that a cultivar-specific temporal expression pattern could influence the timing and intensity of aroma compound synthesis. In 2020, the highest expression of the
VvLOX gene was detected in Cabernet Sauvignon and Chardonnay during E-L 34. In the early maturation stages, E-L 34, 35, and 36, the expression of
VvLOX in Cabernet Franc and Marselan was significantly lower than that in the other varieties. During the ripening stages, the
VvLOX gene exhibited markedly elevated expression in Cabernet Sauvignon and Cabernet Gernischt relative to other grape varieties. Despite this, the levels of C6/C9 aldehydes in these two varieties did not significantly surpass those in other grapes at the same stages. This observation indicates a potential delay in the
VvLOX-mediated regulation of C6/C9 aldehyde accumulation in Cabernet Sauvignon and Cabernet Gernischt (
Figure 2 and
Figure 6).
Hydroperoxide lyase (HPL) breaks down fatty acid hydroperoxides into C6/C9 aldehydes [
6,
40]. Over the two years, the levels of
VvHPL in Chardonnay and Sauvignon Blanc generally increased with growth and development, while those in the other varieties generally increased initially and then decreased during the maturation period, suggesting genetic differences in the regulation of the LOX-HPL pathway (
Figure 5b,g). In 2019, during E-L 34, the highest expression of
VvHPL occurred in Sauvignon Blanc, followed by Cabernet Sauvignon and Marselan, which could impact the early formation of aroma compounds. In the early maturation stages E-L 35 and 36, the levels of
VvHPL in Cabernet Gernischt were markedly greater than those in other varieties. At E-L 38, the expression in Sauvignon Blanc and Cabernet Gernischt was notably higher compared to other varietals. This is consistent with the development of aldehydes (
Figure 2). In 2020, during the E-L 34 stage, the expression of
VvHPL in Cabernet Sauvignon and Cabernet France was significantly greater than that in the other varieties. At the mature stage, the expression in Cabernet Sauvignon was significantly greater than that in the other varieties. Contrasting this, the 2020 vintage revealed a marked deviation in this trend within the Chardonnay variety, where gene expression levels first decreased dramatically before showing a notable increase. The initial decrease in gene expression could be attributed to the rapid rise in temperatures and the subsequent stress conditions experienced by the vines. As the season progressed, an increase in humidity and rainfall, alongside stable sunshine hours, may have created conditions conducive to the recovery and enhancement of
VvHPL gene activity, particularly at the later stages of grape development. Chardonnay and Sauvignon Blanc show a positive correlation with (E, E)-2,4-hexadienal and hexanal, in contrast to Marselan, which shows a negative correlation with hexanal. This pattern suggests the presence of distinct
VvHPL regulatory mechanisms in different grape varieties.
Alcohol dehydrogenase (ADH) catalyzes the synthesis of alcohols [
31]. The transcript levels of
VvADH1 and
VvADH2 showed an overall declining trend in the six varieties over the two years. Qian et al. [
23] found that the levels of
VvADH in Muscat Tchervine, Gewurztraminer and Syrah decreased after the veraison stage, similar to the results of our studies. In 2019, the
VvADH1 gene exhibited the highest expression in Chardonnay at harvest, while Marselan having the lowest expression throughout the developmental process (
Figure 5c). In 2020, the expression of the
VvADH1 gene was greatest in Chardonnay at harvest, while Cabernet Franc exhibited relatively low
VvADH1 gene expression throughout the developmental process (
Figure 5h).
VvADH2 is the most abundant among all the
VvADHs. In 2019, at harvest, Chardonnay exhibited the highest
VvADH2 expression (
Figure 5d). In 2020, Chardonnay, Sauvignon Blanc and Cabernet Sauvignon exhibited a trend of increasing and then decreasing expression levels during the growth and development process. Cabernet Gernischt exhibited an increase in
VvADH2 expression, while the other varieties generally exhibited a gradual decrease in
VvADH2 expression with growth and development (
Figure 5i). Among them, Cabernet Sauvignon exhibited the highest expression, and Cabernet Franc exhibited the lowest.
Alcohol acyltransferase (AAT) catalyzes the conversion of low-molecular-weight alcohols into esters [
11]. Over two years,
VvAAT gene expression in Cabernet Gernischt decreased with maturity, while, in other varieties, it generally increased with growth. In 2019, Chardonnay exhibited the highest
VvAAT gene expression at E-L 36 phases (
Figure 5e). The expression of the
VvAAT gene in Sauvignon Blanc was significantly greater than that in the other varieties at mature stage.
VvAAT gene expression in the later stages of maturation in 2020 was significantly greater than that in 2019, possibly due to the lower rainfall in 2020 (
Figure 5j). At the mature stage, the expression of the
VvAAT gene in Mareslan was significantly greater than that in the other varieties. This finding suggested that a cultivar-specific temporal expression pattern could influence the timing and intensity of aroma compound synthesis. In the berries of Chardonnay and Sauvignon Blanc, a pronounced transcription of
VvAAT correlates with elevated levels of C6 esters. This indicates a significant positive relationship between
VvAAT expression and C6 ester accumulation in these grape varieties.
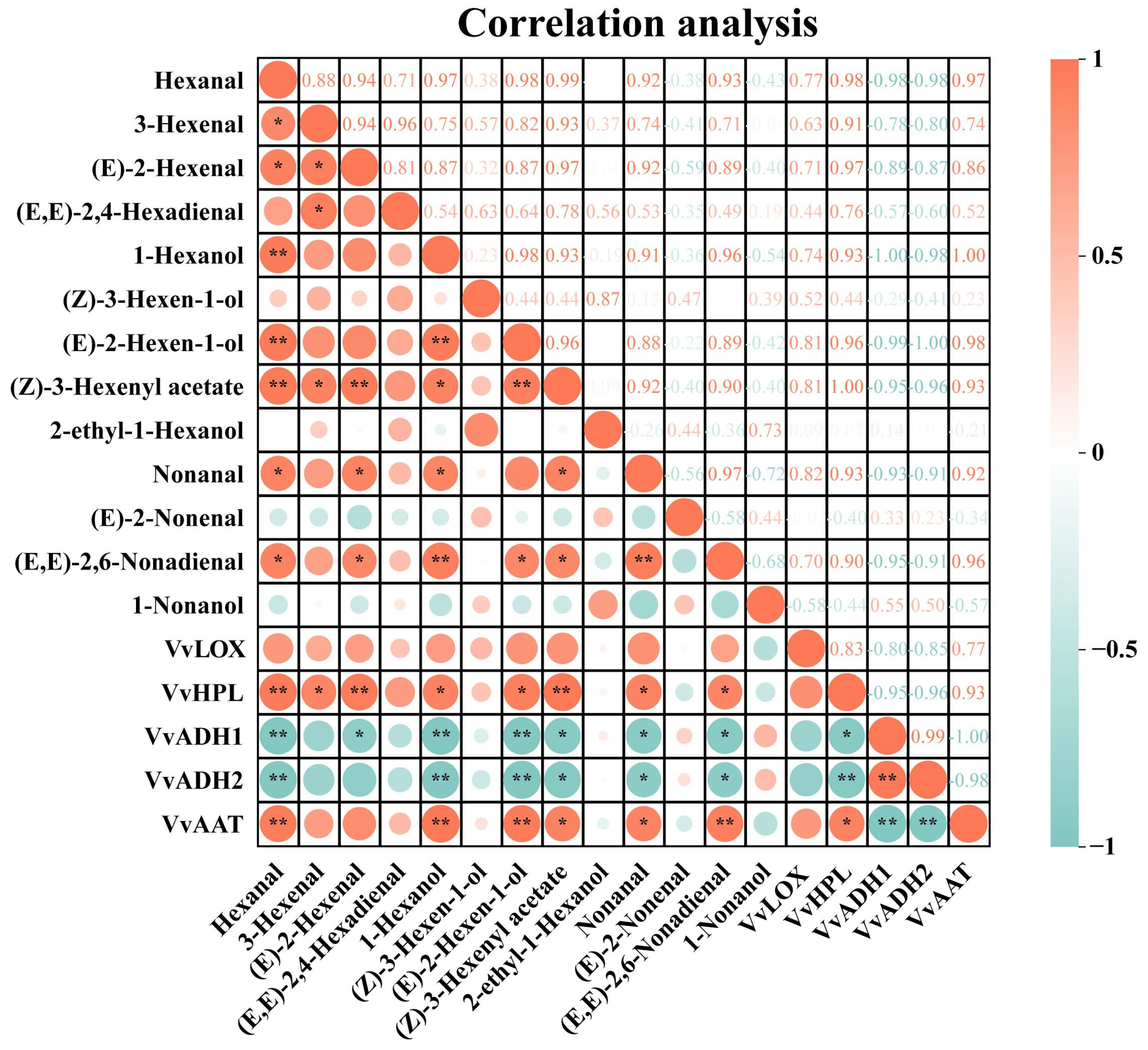
3.7. Correlation Analysis
To improve our understanding of these results, we conducted a correlation analysis between the expression of C6/C9 aromatic substances and LOX-HPL pathway genes in grapes, as shown in
Figure 6. Among the six wine grape varieties,
VvLOX exhibited a positive correlation with most C6/C9 volatile compounds, although the correlation was not significant.
VvHPL exhibited a highly significant positive correlation with hexanal, (Z)-3-hexenyl acetate, and (E)-2-hexenal. It was also significantly positively correlated with 3-hexenal, (E)-2-hexen-1-ol, 1-hexanol, nonanal, and (E, E)-2,6-nonadienal. Both
VvADH1 and
VvADH2 were strongly negatively correlated with hexanal, (E)-2-hexen-1-ol, and 1-hexanol. These genes were significantly negatively correlated with (E, E)-2,6-nonadienal, (Z)-3-hexenyl acetate, and nonanal.
VvAAT had a highly significant positive correlation with 1-hexanol, hexanal, (E, E)-2,6-nonadienal and a significant positive correlation with (Z)-3-hexenyl acetate and nonanal. This discovery suggests a strong connection between the activity of these LOX-HPL pathway genes and the build-up of C6 volatiles. Additionally, hexanal was significantly positively correlated with 3-hexenal, 1-hexanol, and (E, E)-2,6-nonadienal.
Recent studies have demonstrated that viticultural practices and varietal differences regulate the transcription of genes in the LOX-HPL pathway [
41,
42]. Specifically, gene transcription levels within the LOX-HPL pathway vary among wine grape varieties and are closely linked to the production of C6/C9 volatile compounds. Notably, Syrah grapes exhibit a higher concentration of C6 aldehydes, suggesting that increased expression of
VvLOX and
VvHPL genes contributes to this heightened accumulation [
43]. Ju et al. [
9] further highlights the variability in C6 compound concentrations across six varieties of prickly grapes, a variation that correlates with the expression levels of the
VdLOX and
VdHPL genes.
Our findings further reveal that (E)-2-hexenal, hexanal, and (E)-2-hexen-1-ol are most abundant in six grape varieties (
Figure 3). There is a particularly strong positive correlation between the levels of hexanal and (E)-2-hexenal and the expression of
VvHPL. These findings point to
VvHPL as a potential key gene influencing varietal differences in GLV profiles.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





