
Effects of Different Phenolic Compounds on the Redox State of Myoglobin and Prevention of Discoloration, Lipid and Protein Oxidation of Refrigerated Longtail Tuna (Thunnus tonggol) Slices



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Collection of Tuna Sample
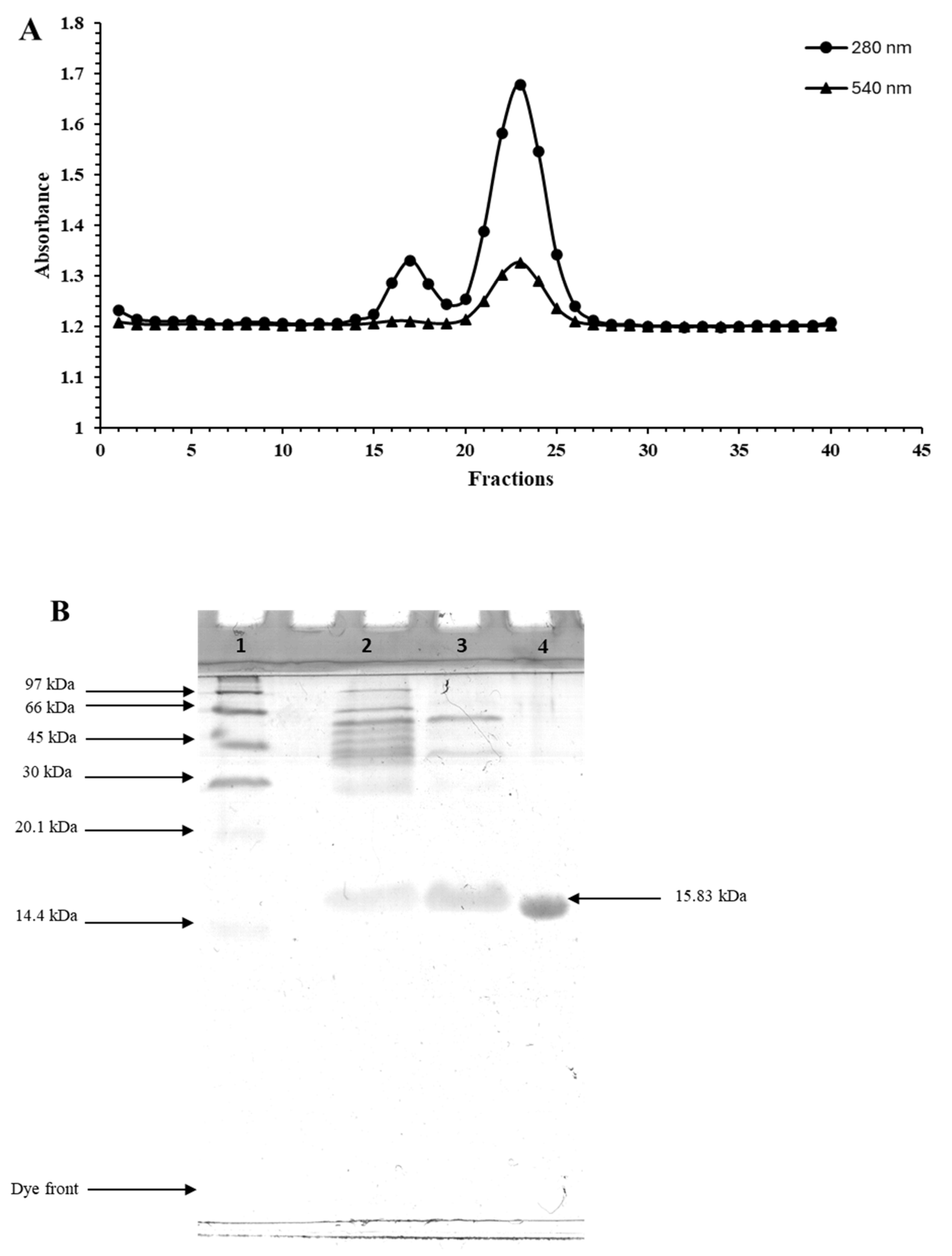
2.3. Purification and Characterization of Myoglobin from Longtail Tuna
2.3.1. Sodium Dodecyl-Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.3.2. Preparation of metMb
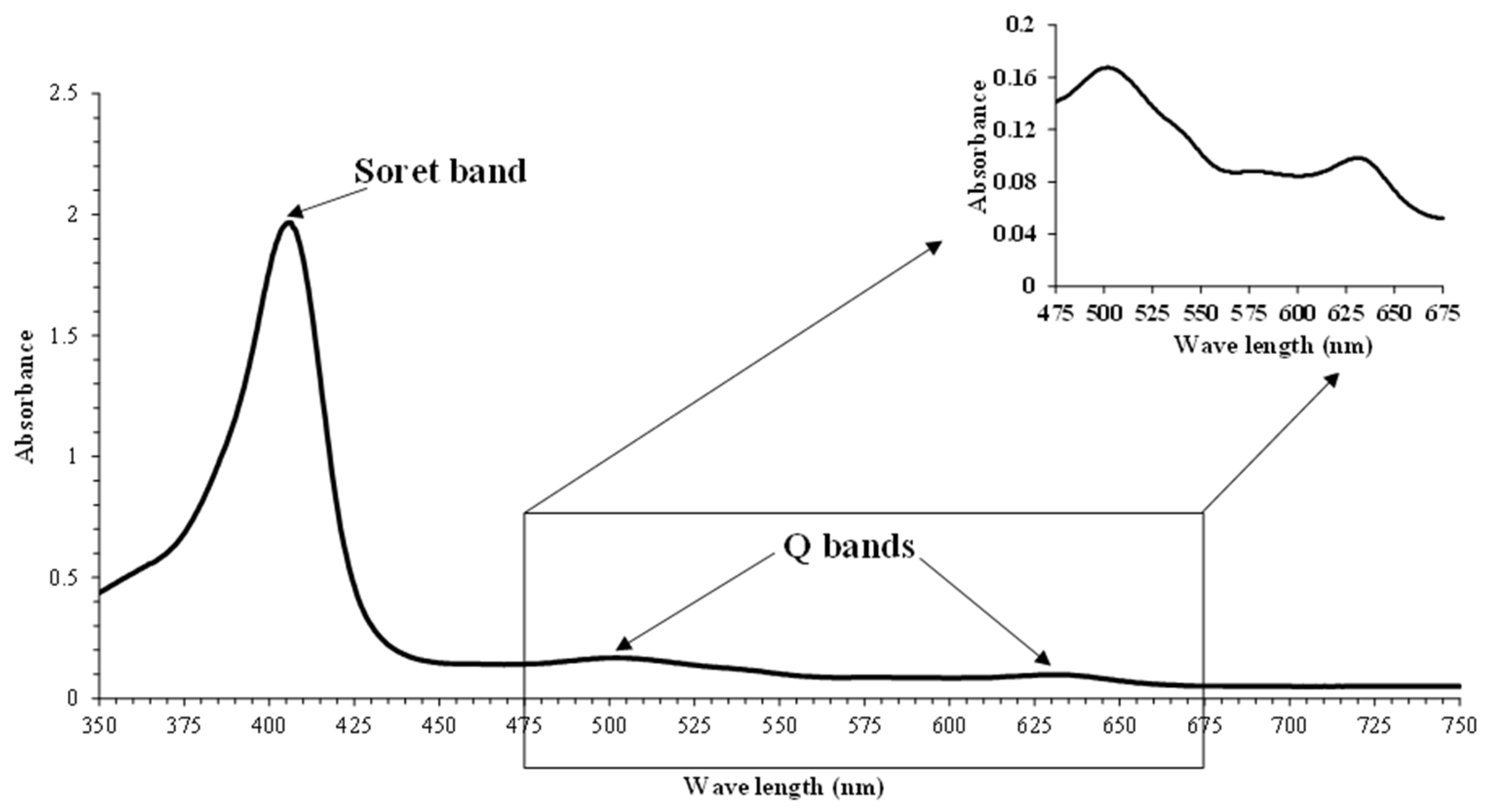
2.3.3. Absorption Spectra and Proportion of Mb Forms
2.4. Effect of Phenolic Compounds on the Proportion of Mb Forms in metMb Solution
2.4.1. Absorption Spectra and Proportion of Mb Forms
2.4.2. Tryptophan Fluorescence
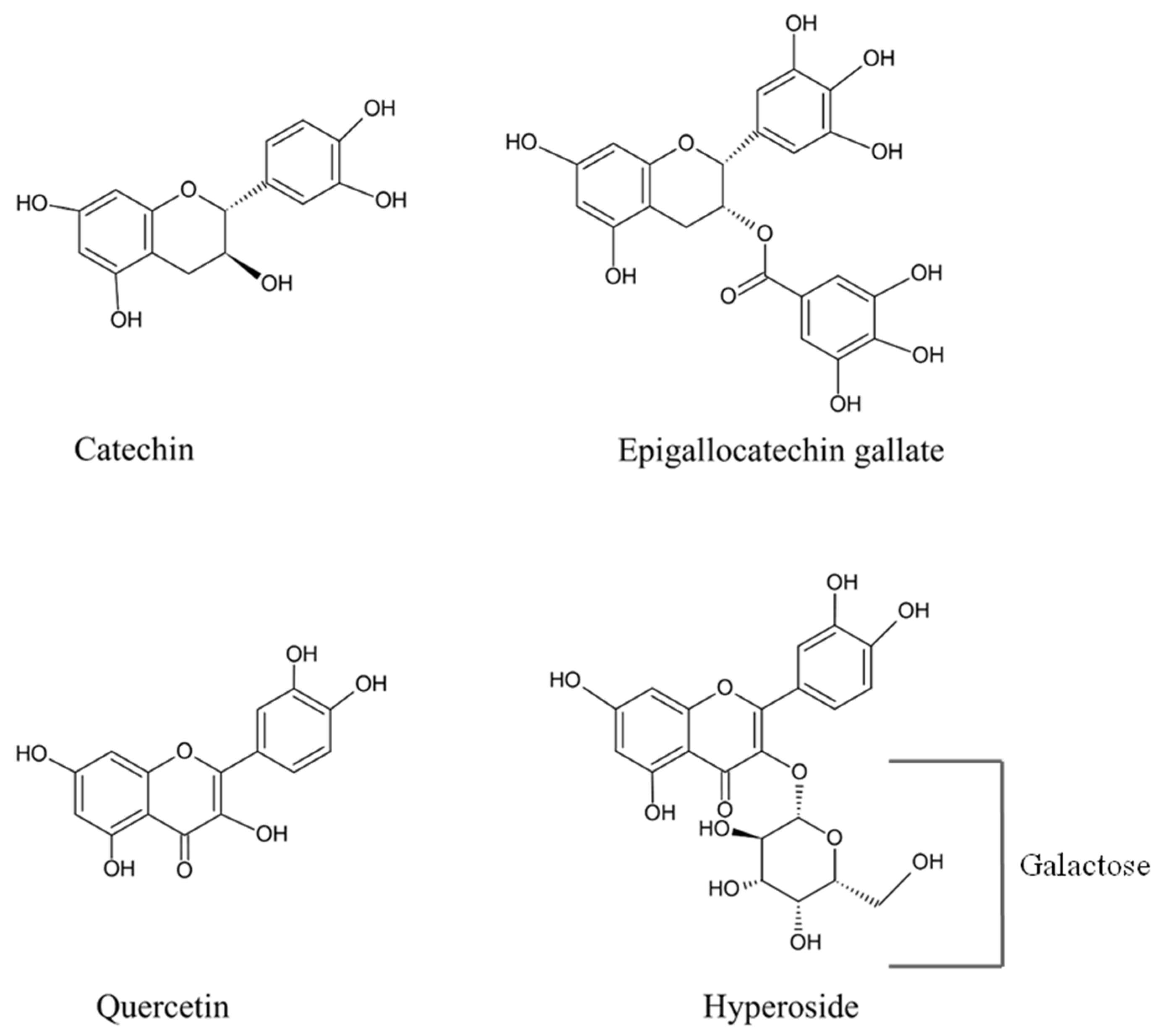
2.5. Effect of EGCG and Quercetin on Mb, Color, and Lipid/Protein Oxidation of Refrigerated Longtail Tuna Slices
2.5.1. Preparation of Tuna Slices Treated with EGCG or Quercetin
Myoglobin Forms and Color
Lipid Oxidation
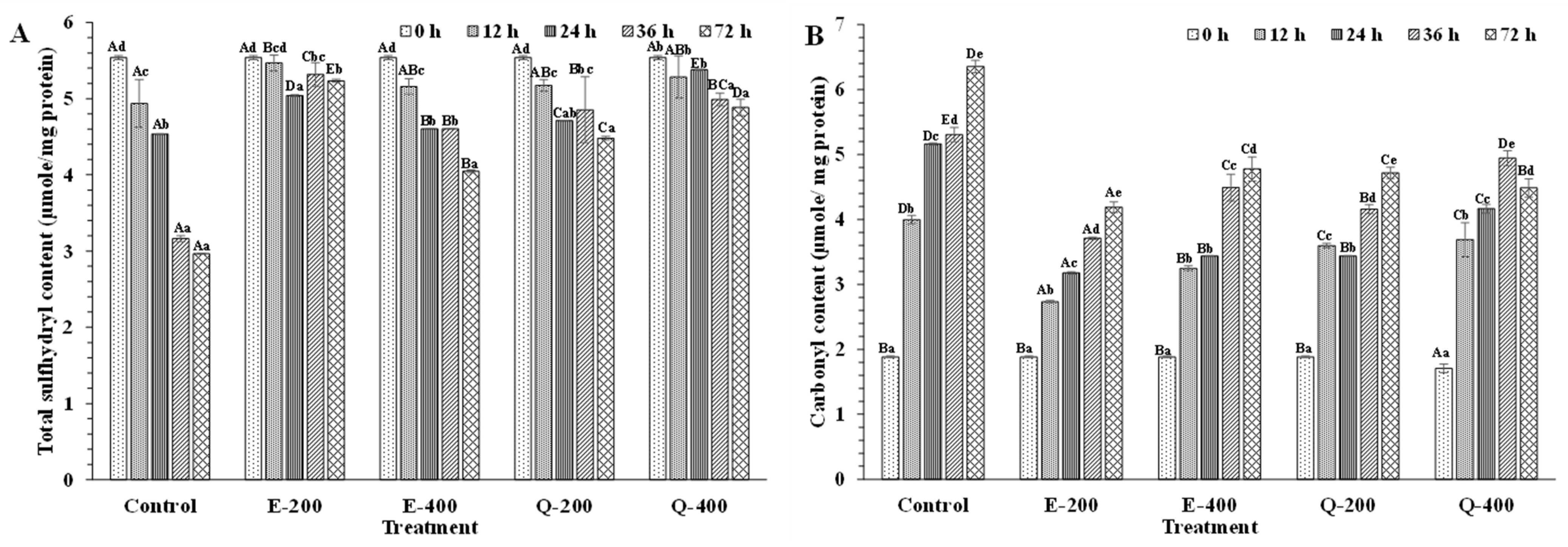
Total Sulfhydryl Content
Protein Carbonyl Content
Fatty Acid Composition
2.6. Statistical Analysis
3. Results and Discussion
3.1. Purification of Myoglobin from Longtail Tuna Dark Muscle
3.2. Absorption Spectra and Proportion of metMb Solutions
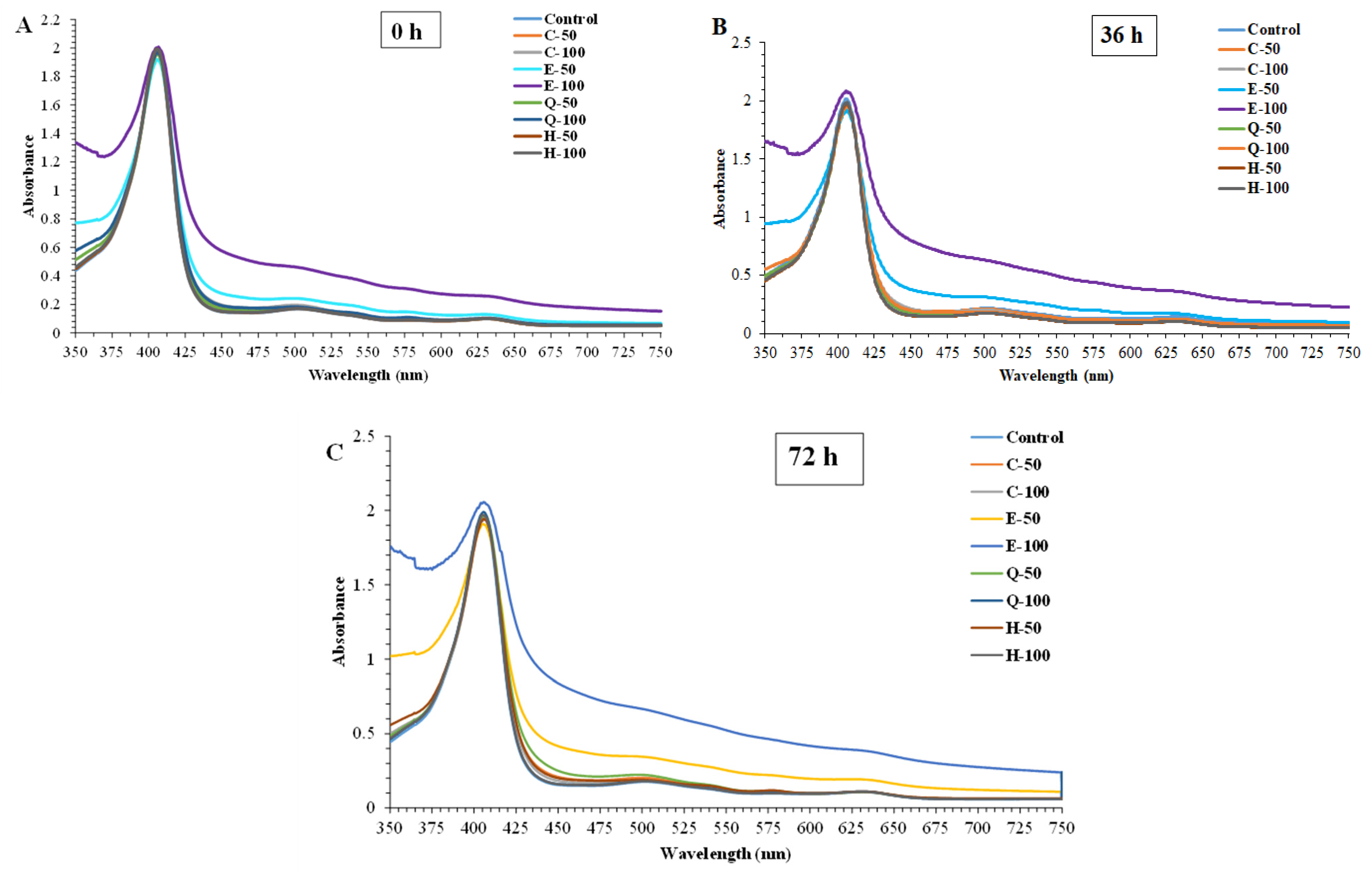
3.3. Effect of Phenolic Compounds on Absorption Spectra and Proportion of Different Forms in metMb Solution
3.3.1. Absorption Spectra
3.3.2. Proportion of Mb Forms
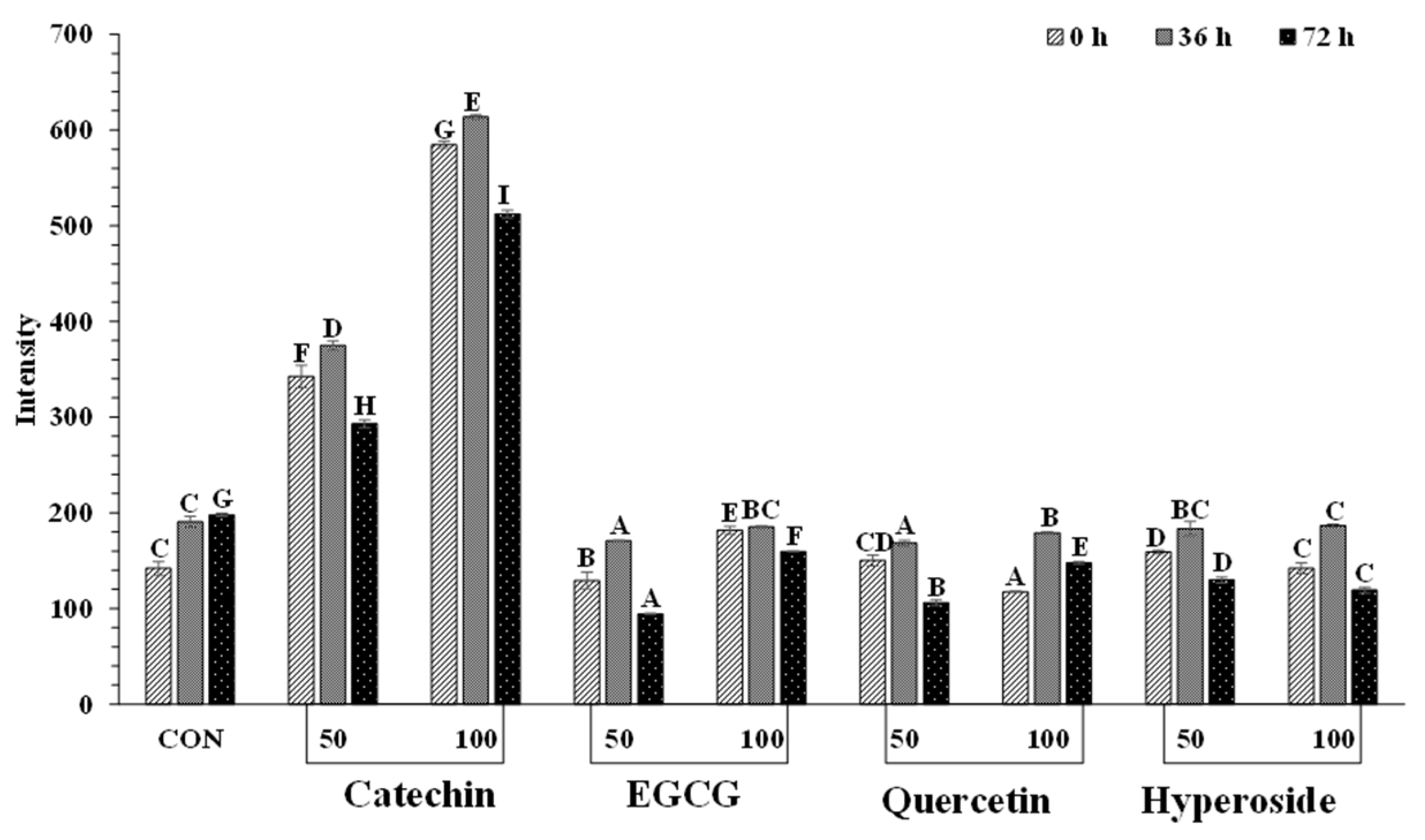
3.3.3. Tryptophan Fluorescence Intensity
3.4. Effect of EGCG and Quercetin on Color, Mb, and Quality of Longtail Tuna Slices during Refrigerated Storage
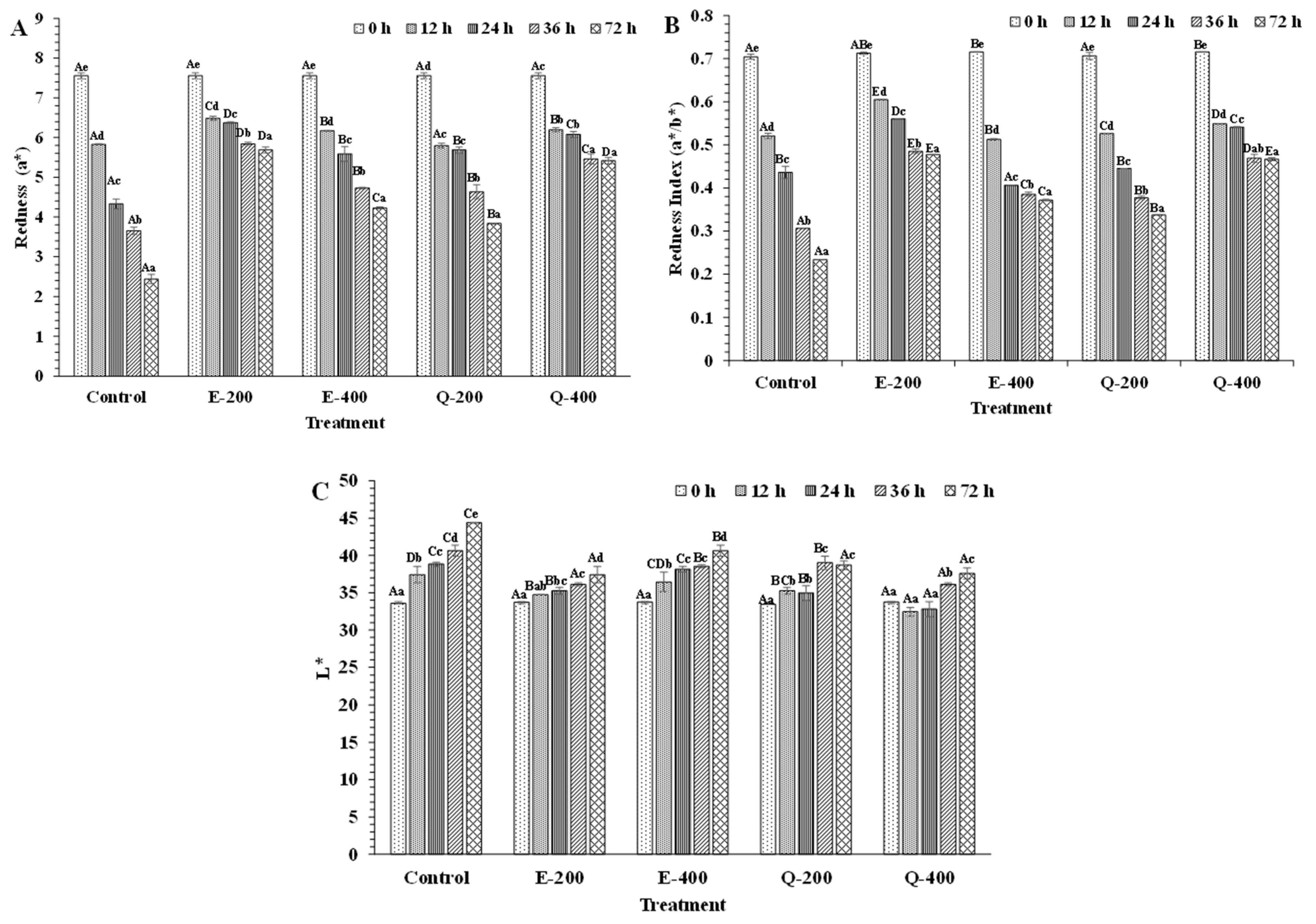
3.4.1. Color
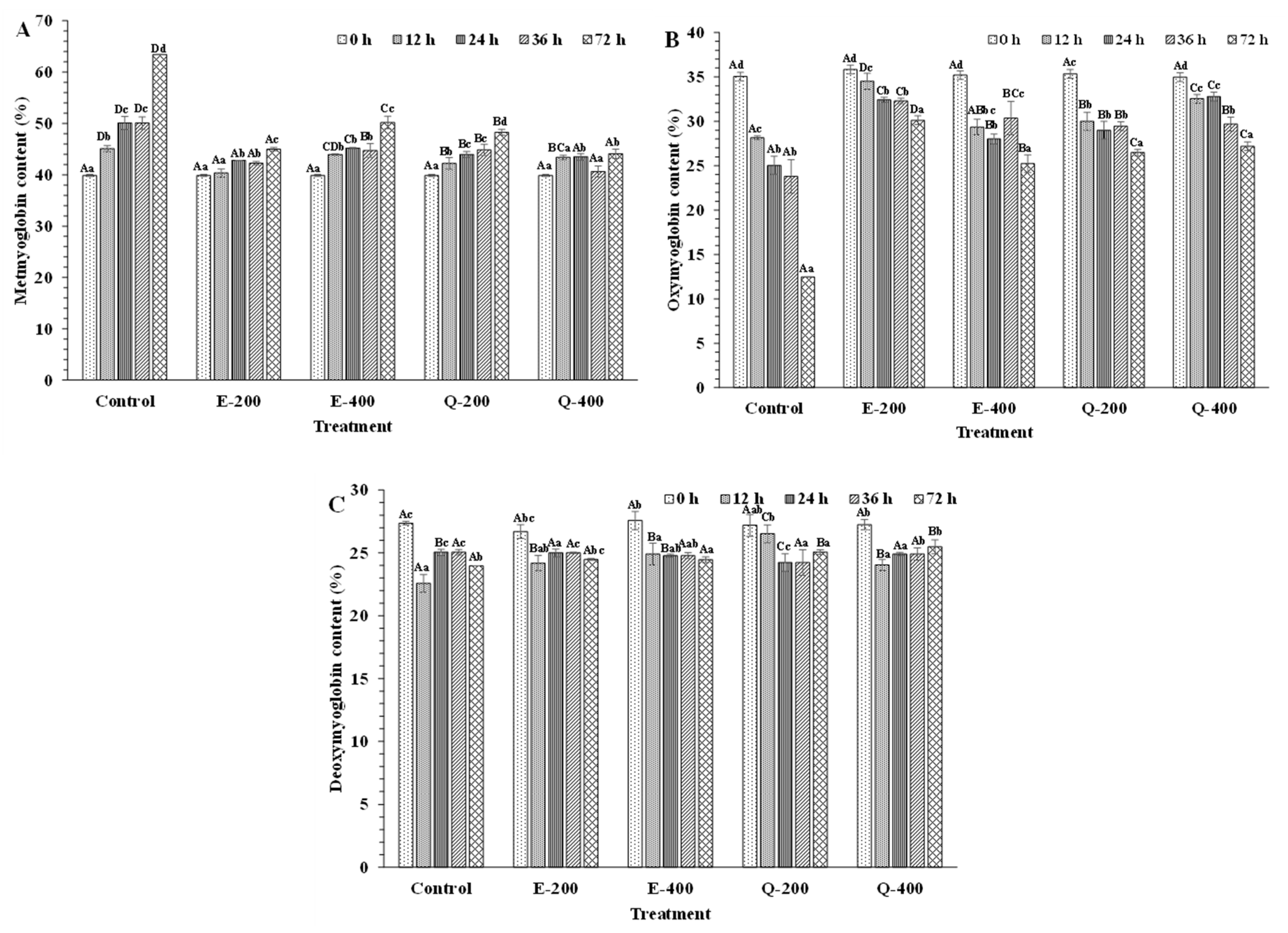
3.4.2. Mb Derivatives
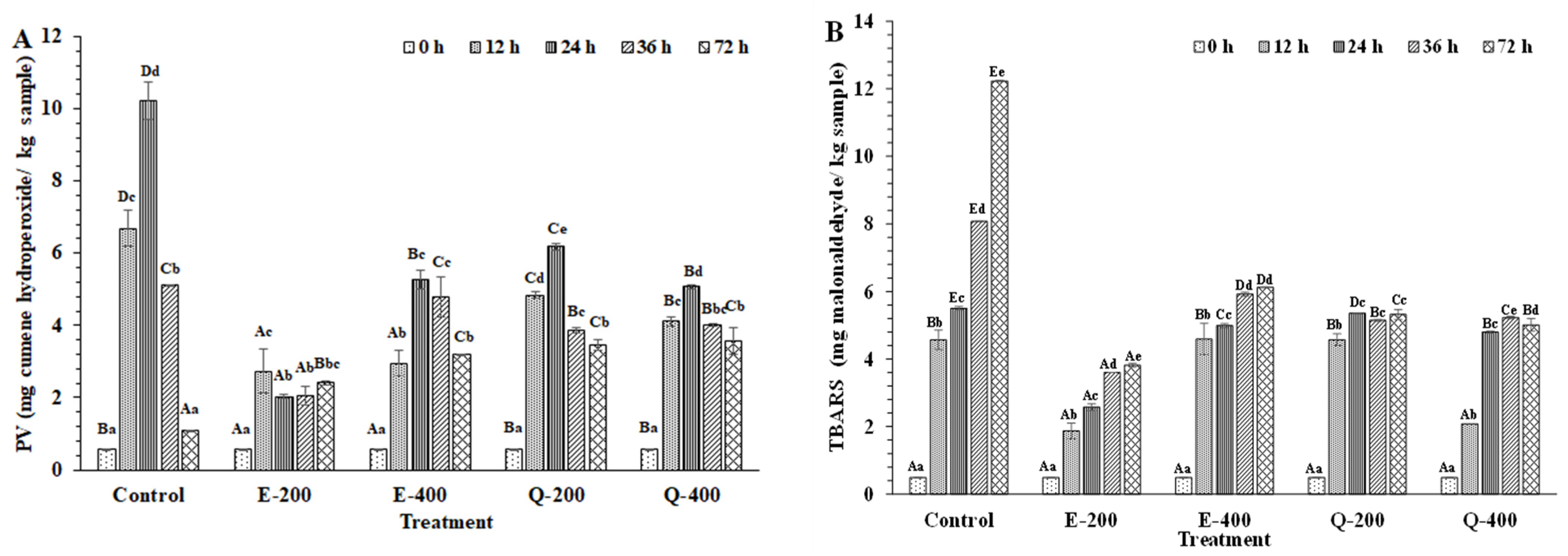
3.4.3. Lipid Oxidation
3.4.4. Protein Oxidation
3.4.5. Fatty Acid Profile
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). GLOBEFISH Highlights—International Markets for Fisheries and Aquaculture Products—Second Issue 2023, with January–December 2022 Statistics; Globefish Highlights; FAO: Rome, Italy, 2023. [Google Scholar] [CrossRef]
- Blanc, M.; Desurmont, A.; Beverly, S. Onboard Handling of Sashimi-Grade Tuna. A Practical Guide for Crew Members; Secretariat of the Pacific Community: Noumea, New Caledonia, 2005; p. 24. [Google Scholar]
- Faustman, C.; Cassens, R.G. The Biochemical Basis for Discoloration in Fresh Meat: A Review. J. Musc. Foods 1990, 1, 217–243. [Google Scholar] [CrossRef]
- Tang, J.; Faustman, C.; Hoagland, T.A. Krzywicki Revisited: Equations for Spectrophotometric Determination of Myoglobin Redox Forms in Aqueous Meat Extracts. J. Food Sci. 2004, 69, C717–C720. [Google Scholar] [CrossRef]
- King, D.A.; Hunt, M.C.; Barbut, S.; Claus, J.R.; Cornforth, D.P.; Joseph, P.; Kim, Y.H.B.; Lindahl, G.; Mancini, R.A.; Nair, M.N. American Meat Science Association Guidelines for Meat Color Measurement. Meat Musc. Biol. 2023, 6, 12473. [Google Scholar] [CrossRef]
- Thiansilakul, Y.; Benjakul, S.; Park, S.Y.; Richards, M.P. Characteristics of Myoglobin and Haemoglobin-Mediated Lipid Oxidation in Washed Mince from Bighead Carp (Hypophthalmichthys nobilis). Food Chem. 2012, 132, 892–900. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, X.; Zhou, G. Covalent Chemical Modification of Myofibrillar Proteins to Improve Their Gelation Properties: A Systematic Review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 924–959. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.P.; Joseph, P. Myoglobin Chemistry and Meat Color. Annu. Rev. Food Sci. Technol. 2013, 4, 79–99. [Google Scholar] [CrossRef]
- Cornforth, D.P.; Jayasingh, P. Chemical and Physical Characteristics of Meat/Color and Pigment; Elsevier Science Ltd.: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Wang, R.; Hu, X.; Agyekumwaa, A.K.; Li, X.; Xiao, X.; Yu, Y. Synergistic Effect of Kojic Acid and Tea Polyphenols on Bacterial Inhibition and Quality Maintenance of Refrigerated Sea Bass (Lateolabrax japonicus) Fillets. LWT-Food Sci. Technol. 2021, 137, 110452. [Google Scholar] [CrossRef]
- Singh, A.; Mittal, A.; Benjakul, S. Undesirable Discoloration in Edible Fish Muscle: Impact of Indigenous Pigments, Chemical Reactions, Processing, and Its Prevention. Compr. Rev. Food Sci. Food Saf. 2022, 21, 580–603. [Google Scholar] [CrossRef]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef]
- Zhu, W.; Han, M.; Bu, Y.; Li, X.; Yi, S.; Xu, Y.; Li, J. Plant Polyphenols Regulating Myoglobin Oxidation and Color Stability in Red Meat and Certain Fish: A Review. Crit. Rev. Food Sci. Nutr. 2024, 64, 2276–2288. [Google Scholar] [CrossRef]
- Czubinski, J.; Dwiecki, K. A Review of Methods Used for Investigation of Protein–Phenolic Compound Interactions. Int. J. Food Sci. Technol. 2017, 52, 573–585. [Google Scholar] [CrossRef]
- Liu, F.; Sun, C.; Yang, W.; Yuan, F.; Gao, Y. Structural Characterization and Functional Evaluation of Lactoferrin–Polyphenol Conjugates Formed by Free-Radical Graft Copolymerization. RSC Adv. 2015, 5, 15641–15651. [Google Scholar] [CrossRef]
- Buitimea-Cantúa, N.E.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Phenolic–Protein Interactions: Effects on Food Properties and Health Benefits. J. Med. Food 2018, 21, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Shahidi, F. Lipophilised Epigallocatechin Gallate (EGCG) Derivatives and Their Antioxidant Potential in Food and Biological Systems. Food Chem. 2012, 131, 22–30. [Google Scholar] [CrossRef]
- Nikoo, M.; Regenstein, J.M.; Ahmadi Gavlighi, H. Antioxidant and Antimicrobial Activities of (-)-Epigallocatechin-3-gallate (EGCG) and Its Potential to Preserve the Quality and Safety of Foods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 732–753. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, M.; Xu, Y.-F.; Feng, Y.; Che, J.-P.; Wang, G.-C.; Zheng, J.-H. Combination of Quercetin and Hyperoside Has Anticancer Effects on Renal Cancer Cells through Inhibition of Oncogenic microRNA-27a. Oncol. Rep. 2014, 31, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Zamora, R.; Hidalgo, F.J. The Triple Defensive Barrier of Phenolic Compounds against the Lipid Oxidation-Induced Damage in Food Products. Trends Food Sci. Technol. 2016, 54, 165–174. [Google Scholar] [CrossRef]
- Dragoev, S.G. Lipid Peroxidation in Muscle Foods: Impact on Quality, Safety and Human Health. Foods 2024, 13, 797. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Benjakul, S.; Zhang, B.; Deng, S.; Mittal, A. Effect of Squid Pen Chitooligosaccharide in Conjugation with Different Modified Atmospheric Packaging Conditions on Color and Storage Stability of Tuna Slices. Food Control 2021, 125, 108013. [Google Scholar] [CrossRef]
- Lee, S.; Joo, S.T.; Alderton, A.L.; Hill, D.W.; Faustman, C. Oxymyoglobin and Lipid Oxidation in Yellowfin Tuna (Thunnus albacares) Loins. J. Food Sci. 2003, 68, 1664–1668. [Google Scholar] [CrossRef]
- Chung, C.; Baier, S.; McClements, D.J.; Decker, E.A. Stabilization of Myoglobin from Different Species (Produced by Cellular Agriculture) Using Food-Grade Natural and Synthetic Antioxidants. Food Res. Int. 2024, 178, 113965. [Google Scholar] [CrossRef] [PubMed]
- Trout, G.R.; Gutzke, D.A. A Simple, Rapid Preparative Method for Isolating and Purifying Oxymyoglobin. Meat Sci. 1996, 43, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Swatland, H.J. A Review of Meat Spectrophotometry (300 to 800 nm). Can. Inst. Food Sci. Technol. J. 1989, 22, 390–402. [Google Scholar] [CrossRef]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Changes of Pigments and Color in Sardine (Sardinella gibbosa) and Mackerel (Rastrelliger kanagurta) Muscle during Iced Storage. Food Chem. 2005, 93, 607–617. [Google Scholar] [CrossRef]
- Thiansilakul, Y.; Benjakul, S.; Richards, M.P. Isolation, Characterisation and Stability of Myoglobin from Eastern Little Tuna (Euthynnus affinis) Dark Muscle. Food Chem. 2011, 124, 254–261. [Google Scholar] [CrossRef]
- Singh, A.; Benjakul, S.; Zhou, P.; Zhang, B.; Deng, S. Effect of Squid Pen Chitooligosaccharide and Epigallocatechin Gallate on Discoloration and Shelf-Life of Yellowfin Tuna Slices during Refrigerated Storage. Food Chem. 2021, 351, 129296. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chiu, E.; Huang, J. Color and Gel-forming Properties of Horse Mackerel (Trachurus japonicus) as Related to Washing Conditions. J. Food Sci. 1997, 62, 985–991. [Google Scholar] [CrossRef]
- Arfat, Y.A.; Benjakul, S.; Vongkamjan, K.; Sumpavapol, P.; Yarnpakdee, S. Shelf-Life Extension of Refrigerated Sea Bass Slices Wrapped with Fish Protein Isolate/Fish Skin Gelatin-ZnO Nanocomposite Film Incorporated with Basil Leaf Essential Oil. J. Food Sci. Technol. 2015, 52, 6182–6193. [Google Scholar] [CrossRef]
- Benjakul, S.; Bauer, F. Biochemical and Physicochemical Changes in Catfish (Silurus glanis Linne) Muscle as Influenced by Different Freeze–Thaw Cycles. Food Chem. 2001, 72, 207–217. [Google Scholar] [CrossRef]
- Sriket, P.; Benjakul, S.; Visessanguan, W.; Kijroongrojana, K. Comparative Studies on the Effect of the Freeze–Thawing Process on the Physicochemical Properties and Microstructures of Black Tiger Shrimp (Penaeus monodon) and White Shrimp (Penaeus vannamei) Muscle. Food Chem. 2007, 104, 113–121. [Google Scholar] [CrossRef]
- Nikoo, M.; Benjakul, S.; Xu, X. Antioxidant and Cryoprotective Effects of Amur Sturgeon Skin Gelatin Hydrolysate in Unwashed Fish Mince. Food Chem. 2015, 181, 295–303. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Raju, N.; Benjakul, S. Use of Beta Cyclodextrin to Remove Cholesterol and Increase Astaxanthin Content in Shrimp Oil. Eur. J. Lipid Sci. Technol. 2020, 122, 1900242. [Google Scholar] [CrossRef]
- Thiansilakul, Y.; Benjakul, S.; Richards, M.P. Effect of Phenolic Compounds in Combination with Modified Atmospheric Packaging on Inhibition of Quality Losses of Refrigerated Eastern Little Tuna Slices. LWT-Food Sci. Technol. 2013, 50, 146–152. [Google Scholar] [CrossRef]
- Fosmire, G.J.; Brown, W.D. Yellowfin Tuna (Thunnus albacares) Myoglobin: Characterization and Comparative Stability. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1976, 55, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Ueki, N.; Ochiai, Y. Primary Structure and Thermostability of Bigeye Tuna Myoglobin in Relation to Those of Other Scombridae Fish. Fish. Sci. 2004, 70, 875–884. [Google Scholar] [CrossRef]
- Yamaguchi, K. Properties of Mackerel and Sardine Myoglobins. Bull. Jpn. Soc. Sci. Fish. 1979, 45, 1335–1339. [Google Scholar] [CrossRef]
- Chotichayapong, C.; Sattayasai, N.; Kanzawa, N.; Tamiya, T.; Tsuchiya, T.; Chanthai, S. Purification, Peptide Mapping and Spectroscopic Characterization of Myoglobin from Striped Snake-Head Fish (Ophicephalus striatus). Orient. J. Chem. 2016, 32, 181. [Google Scholar] [CrossRef]
- Renerre, M.; Anton, M.; Gatellier, P. Autoxidation of Purified Myoglobin from Two Bovine Muscles. Meat Sci. 1992, 32, 331–342. [Google Scholar] [CrossRef]
- Satterlee, L.D.; Zachariah, N.Y. Porcine and Ovine Myoglobin: Isolation, Purification, Characterization and Stability. J. Food Sci. 1972, 37, 909–912. [Google Scholar] [CrossRef]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Lee, S.; Faustman, C. The Effect of Freezing and Aldehydes on the Interaction between Fish Myoglobin and Myofibrillar Proteins. J. Agric. Food Chem. 2007, 55, 4562–4568. [Google Scholar] [CrossRef] [PubMed]
- Smulevich, G.; Droghetti, E.; Focardi, C.; Coletta, M.; Ciaccio, C.; Nocentini, M. A Rapid Spectroscopic Method to Detect the Fraudulent Treatment of Tuna Fish with Carbon Monoxide. Food Chem. 2007, 101, 1071–1077. [Google Scholar] [CrossRef]
- Chen, W.-L.; Chow, C.-J. Studies on The Physicochemical Properties of Milkfish Myoglobin. J. Food Biochem. 2001, 25, 157–174. [Google Scholar] [CrossRef]
- Nurilmala, M.; Ushio, H.; Ochiai, Y. pH- and Temperature-Dependent Denaturation Profiles of Tuna Myoglobin. Fish. Sci. 2018, 84, 579–587. [Google Scholar] [CrossRef]
- Ke, Z.; Bai, Y.; Zhu, H.; Xiang, X.; Liu, S.; Zhou, X.; Ding, Y. Characteristics of Myoglobin Degradation by Cold Plasma and Its Pro-Oxidative Activity on Lipid in Washed Fish Muscle. Food Chem. 2022, 389, 132972. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.R.; Zipser, M.W.; Watts, B.M. The Use of Reflectance Spectrophotometry for the Assay of Raw Meat Pigments. J. Food Sci. 1965, 30, 464–469. [Google Scholar] [CrossRef]
- Krzywicki, K. Assessment of Relative Content of Myoglobin, Oxymyoglobin and Metmyoglobin at the Surface of Beef. Meat Sci. 1979, 3, 1–10. [Google Scholar] [CrossRef]
- Engler, N.; Ostermann, A.; Gassmann, A.; Lamb, D.C.; Prusakov, V.E.; Schott, J.; Schweitzer-Stenner, R.; Parak, F.G. Protein Dynamics in an Intermediate State of Myoglobin: Optical Absorption, Resonance Raman Spectroscopy, and X-ray Structure Analysis. Biophys. J. 2000, 78, 2081–2092. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, R.N.; Machado, A.E.H.; Piovesan, E.; Gonçalves, P.J.; De Paula, R.; Cavaleiro, J.A.S.; Neto, N.B. Influence of the Solvent-Porphyrin Interaction on the UV–Vis Absorption of Free Base Imidazol Cationic Porphyrin. Dye. Pigment. 2014, 100, 73–78. [Google Scholar] [CrossRef]
- Kingsbury, C.J.; Senge, M.O. The Shape of Porphyrins. Coord. Chem. Rev. 2021, 431, 213760. [Google Scholar] [CrossRef]
- Baron, C.P.; Skibsted, L.H.; Andersen, H.J. Concentration Effects in Myoglobin-Catalyzed Peroxidation of Linoleate. J. Agric. Food Chem. 2002, 50, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.H.; Benjakul, S.; Sae-leaw, T.; Balange, A.K.; Maqsood, S. Protein–Polyphenol Conjugates: Antioxidant Property, Functionalities and Their Applications. Trends Food Sci. Technol. 2019, 91, 507–517. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S. Phenolic Compounds of Green Tea: Health Benefits and Technological Application in Food. Asian Pac. J. Trop. Biomed. 2016, 6, 709–719. [Google Scholar] [CrossRef]
- Predescu, C.; Papuc, C.; Petcu, C.; Goran, G.; Rus, A.E. The Effect of Some Polyphenols on Minced Pork during Refrigeration Compared with Ascorbic Acid. Bull. UASVM Food Sci. Technol. 2018, 75, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Faustman, C.; Sun, Q.; Mancini, R.; Suman, S.P. Myoglobin and Lipid Oxidation Interactions: Mechanistic Bases and Control. Meat Sci. 2010, 86, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Copeland, R.A. Spectroscopic Probes of Protein Structure. In Methods for Protein Analysis; Springer: Boston, MA, USA, 1994; pp. 161–198. ISBN 978-1-4757-1507-1. [Google Scholar]
- Chanthai, S.; Ogawa, M.; Tamiya, T.; Tsuchiya, T. Studies on Thermal Denaturation of Fish Myoglobins Using Differential Scanning Calorimetry, Circular Dichroism, and Tryptophan Fluorescence. Fish. Sci. 1996, 62, 927–932. [Google Scholar] [CrossRef]
- Sirangelo, I.; Tavassi, S.; Martelli, P.L.; Casadio, R.; Irace, G. The Effect of Tryptophanyl Substitution on Folding and Structure of Myoglobin. Eur. J. Biochem. 2000, 267, 3937–3945. [Google Scholar] [CrossRef] [PubMed]
- Snyder, H.E. Measurement of Discoloration in Fresh Beef. J. Food Sci. 1964, 29, 535–539. [Google Scholar] [CrossRef]
- Dickson, K.A. Locomotor Muscle of High-Performance Fishes: What Do Comparisons of Tunas with Ectothermic Sister Taxa Reveal? Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1996, 113, 39–49. [Google Scholar] [CrossRef]
- Wu, H.; Yin, J.; Xiao, S.; Zhang, J.; Richards, M.P. Quercetin as an Inhibitor of Hemoglobin-Mediated Lipid Oxidation: Mechanisms of Action and Use of Molecular Docking. Food Chem. 2022, 384, 132473. [Google Scholar] [CrossRef]
- Awad, H.M.; Boersma, M.G.; Vervoort, J.; Rietjens, I.M. Peroxidase-Catalyzed Formation of Quercetin Quinone Methide–Glutathione Adducts. Arch. Biochem. Biophys. 2000, 378, 224–233. [Google Scholar] [CrossRef]
- Ito, N.; Komiyama, N.H.; Fermi, G. Structure of Deoxyhaemoglobin of the Antarctic Fish Pagothenia bernacchii with an Analysis of the Structural Basis of the Root Effect by Comparison of the Liganded and Unliganded Haemoglobin Structures. J. Mol. Biol. 1995, 250, 648–658. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A Review on Protein–Phenolic Interactions and Associated Changes. Int. Food Res. J. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Stepanic, V.; Gasparovic, A.C.; Troselj, K.G.; Amic, D.; Zarkovic, N. Selected Attributes of Polyphenols in Targeting Oxidative Stress in Cancer. Curr. Top. Med. Chem. 2015, 15, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Cherviakovsky, E.M.; Bolibrukh, D.A.; Baranovsky, A.V.; Vlasova, T.M.; Kurchenko, V.P.; Gilep, A.A.; Usanov, S.A. Oxidative Modification of Quercetin by Hemeproteins. Biochem. Biophys. Res. Commun. 2006, 342, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Achike, F.I.; Murugan, D.D. Quercetin and Antioxidant Potential in Diabetes. In Diabetes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 293–302. [Google Scholar]
- Masniyom, P.; Benjakul, S.; Visessanguan, W. Shelf-life Extension of Refrigerated Seabass Slices under Modified Atmosphere Packaging. J. Sci. Food Agric. 2002, 82, 873–880. [Google Scholar] [CrossRef]
- Chiou, T.-K.; Pong, C.-Y.; Nieh, F.-P.; Jiang, S.-T. Effect of Met-Myoglobin Reductase on the Color Stability of Blue Fin Tuna during Refrigerated Storage. Fish. Sci. 2001, 67, 694–702. [Google Scholar] [CrossRef]
- Viana, F.M.; Wang, Y.; Li, S.; Conte-Junior, C.A.; Chen, J.; Zhu, H.; Suman, S.P.; Conte-Junior, C.A.; Suman, S.P. Thermal Instability Induced by 4-Hydroxy-2-Nonenal in Beef Myoglobin. Meat Mus. Biol. 2020. [Google Scholar] [CrossRef]
- Bekhit, A.E.D.; Faustman, C. Metmyoglobin Reducing Activity. Meat Sci. 2005, 71, 407–439. [Google Scholar] [CrossRef]
- Ramanathan, R.; Nair, M.N.; Hunt, M.C.; Suman, S.P. Mitochondrial Functionality and Beef Colour: A Review of Recent Research. S. Afr. J. Anim. Sci. 2019, 49, 9–19. [Google Scholar] [CrossRef]
- Seyfert, M.; Mancini, R.A.; Hunt, M.C.; Tang, J.; Faustman, C.; Garcia, M. Color Stability, Reducing Activity, and Cytochrome C Oxidase Activity of Five Bovine Muscles. J. Agric. Food Chem. 2006, 54, 8919–8925. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Zhong, Y. Measurement of Antioxidant Activity. J. Funct. Food 2015, 18, 757–781. [Google Scholar] [CrossRef]
- Nawar, W.W. Thermal Degradation of Lipids. J. Agric. Food Chem. 1969, 17, 18–21. [Google Scholar] [CrossRef]
- Vieira, S.A.; Zhang, G.; Decker, E.A. Biological Implications of Lipid Oxidation Products. J. Am. Oil Chem. Soc. 2017, 94, 339–351. [Google Scholar] [CrossRef]
- Wei, P.; Zhu, K.; Cao, J.; Dong, Y.; Li, M.; Shen, X.; Duan, Z.; Li, C. The Inhibition Mechanism of the Texture Deterioration of Tilapia Fillets during Partial Freezing after Treatment with Polyphenols. Food Chem. 2021, 335, 127647. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, M.; Mujumdar, A.S. Technological Innovations or Advancement in Detecting Frozen and Thawed Meat Quality: A Review. Crit. Rev. Food Sci. Nutr. 2023, 63, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Bu, Y.; Han, M.; Tan, G.; Zhu, W.; Li, X.; Li, J. Changes in Quality Characteristics of Southern Bluefin Tuna (Thunnus maccoyii) during Refrigerated Storage and Their Correlation with Color Stability. LWT-Food Sci. Technol. 2022, 154, 112715. [Google Scholar] [CrossRef]
- Ma, J.; Wang, X.; Li, Q.; Zhang, L.; Wang, Z.; Han, L.; Yu, Q. Oxidation of Myofibrillar Protein and Crosslinking Behavior during Processing of Traditional Air-Dried Yak (Bos grunniens) Meat in Relation to Digestibility. LWT-Food Sci. Technol. 2021, 142, 110984. [Google Scholar] [CrossRef]
- Jiang, S.; Xue, D.; Zhang, Z.; Shan, K.; Ke, W.; Zhang, M.; Zhao, D.; Nian, Y.; Xu, X.; Zhou, G. Effect of Sous-Vide Cooking on the Quality and Digestion Characteristics of Braised Pork. Food Chem. 2022, 375, 131683. [Google Scholar] [CrossRef]
- Matsuura, F. Studies on the Autoxidation Velocity of Fish Myoglobin. Nippon Suisan Gakkaishi 1962, 28, 210–216. [Google Scholar] [CrossRef]
- Bazargani-Gilani, B.; Aliakbarlu, J.; Tajik, H. Effect of Pomegranate Juice Dipping and Chitosan Coating Enriched with Zataria multiflora Boiss Essential Oil on the Shelf-Life of Chicken Meat during Refrigerated Storage. Innov. Food Sci. Emerg. Technol. 2015, 29, 280–287. [Google Scholar] [CrossRef]
- Hematyar, N.; Rustad, T.; Sampels, S.; Kastrup Dalsgaard, T. Relationship between Lipid and Protein Oxidation in Fish. Aquac. Res. 2019, 50, 1393–1403. [Google Scholar] [CrossRef]
- Huang, G.; Tian, Z.; Xue, L.; Chen, Q.; Wang, D.; Yi, Y.; Li, M.; Sun, S.; Sui, X. Effect of Interaction between EGCG and Soy β-Conglycinin/Glycinin on Protein Structure. Food Sci. 2022, 43, 1–6. [Google Scholar]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Mittal, A.; Singh, A.; Benjakul, S. The Impact of Chitosan Film or Chitosan/Chitosan–Epigallocatechin Gallate Conjugate Composite Film on the Quality Changes of Asian Sea Bass (Lates calcarifer) Slices Stored in Air or under Vacuum Packaging. Int. J. Food Sci. Technol. 2021, 56, 6025–6038. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | Formula | Fresh | Control | E-200 | E-400 | Q-200 | Q-400 |
|---|---|---|---|---|---|---|---|
| Lauric acid | C12:0 | 0.13 ± 0.02 b | 0.08 ± 0.02 cd | 0.06 ± 0 d | 0.59 ± 0.01 a | 0.09 ± 0.02 c | 0.09 ± 0.01 c |
| Myristic acid | C14:0 | 1.58 ± 0.03 d | 2.03 ± 0.17 c | 2.91 ± 0.01 a | 2.16 ± 0.02 c | 2.55 ± 0.28 b | 2.27 ± 0.15 c |
| Pentadecanoic acid | C15:0 | 0.51 ± 0.1 c | 0.68 ± 0.07 b | 0.87 ± 0 a | 0.62 ± 0.01 bc | 0.92 ± 0.1 a | 0.67 ± 0.1 b |
| Palmitic acid | C16:0 | 21.34 ± 0.15 c | 25.61 ± 0.32 a | 26.45 ± 0.11 a | 23.18 ± 0.05 bc | 26.17 ± 0.92 a | 23.57 ± 1 bc |
| Heptadecanoic acid | C17:0 | 0.82 ± 0 c | 0.97 ± 0.09 b | 1.11 ± 0 b | 0.81 ± 0.07 c | 1.46 ± 0.15 a | 1.52 ± 0.05 a |
| Stearic acid | C18:0 | 7.91 ± 0.05 bc | 9.16 ± 0.86 b | 0.09 ± 0.01 bc | 7.62 ± 0.06 c | 11.67 ± 1.45 a | 8.19 ± 0.1 bc |
| Arachidic acid | C20:0 | 0.23 ± 0.09 b | 0.31 ± 0.06 b | 0.97 ± 0.56 a | 0.33 ± 0.01 b | 0.58 ± 0.06 ab | 0.26 ± 0.01 b |
| Docosanoic acid | C22:0 | 0.26 ± 0.09 bc | 0.14 ± 0.02 cd | 0.29 ± 0.08 ab | 0.23 ± 0.09 bcd | 0.4 ± 0.04 a | 0.13 ± 0.02 d |
| Lignoceric acid | C24:0 | 0.28 ± 0.03 bcd | 0.29 ± 0.03 bc | 0.25 ± 0 cd | 0.24 ± 0.01 d | 0.39 ± 0.03 a | 0.3 ± 0.02 b |
| SFA | 33.06 ± 0.51 d | 39.27 ± 3.36 bc | 41 ± 0.50 ab | 35.78 ± 0.02 c | 44.23 ± 5.041 a | 37 ± 1.23 bc | |
| Palmitoleic acid | C16:1 | 3.41 ± 0 d | 4.88 ± 0.45 a | 4.61 ± 0.04 ab | 4.48 ± 0.07 ab | 3.83 ± 0.49 cd | 4.1 ± 0.15 bc |
| Cis-10-Heptadecenoic acid | C17:1 | 0.09 ± 0.03 b | 0.09 ± 0.02 b | 0.13 ± 0 a | 0.15 ± 0.01 a | 0.12 ± 0.02 ab | 0.05 ± 0 c |
| Oleic acid | C18:1 n9c | 15.11 ± 0.07 ab | 17.86 ± 1.7 a | 9.15 ± 0.13 c | 17 ± 0.14 a | 9.09 ± 1.03 c | 15.9 ± 0.5 ab |
| Elaidic acid | C18:1n9t | 0.12 ± 0.01 ab | 0.08 ± 0.04 bcd | 0.07 ± 0.02 cd | 0.05 ± 0.03 d | 0.11 ± 0.01 bc | 0.15 ± 0 a |
| Cis-11-Eicosenoic acid | C120:1 | 0.24 ± 0.18 a | 0.1 ± 0.03 a | 0.67 ± 0.61 a | 0.45 ± 0.4 a | 0.11 ± 0 a | 0.64 ± 0.05 a |
| Erucic acid | C22:1 n9 | 0.05 ± 0 a | 0.19 ± 0.14 a | 0.08 ± 0.01 a | 0.18 ± 0.12 a | 0.12 ± 0.08 a | 0.17 ± 0.02 a |
| Nervonic acid | C24:1 | 0.33 ± 0.04 bc | 0.34 ± 0.05 bc | 0.44 ± 0.02 a | 0.3 ± 0 c | 0.39 ± 0.06 ab | 0.35 ± 0.02 bc |
| MUFA | 19.35 ± 0.18 abc | 23.54 ± 1.99 a | 15.15 ± 9.82 bc | 22.61 ± 0.42 ab | 13.77 ± 1.52 c | 21.36 ± 0.40 abc | |
| Linoleic acid | C18:2n6c | 1.86 ± 0.03 b | 1.74 ± 0.18 b | 9.92 ± 0.42 a | 2.16 ± 0.62 b | 1.61 ± 0.25 b | 1.72 ± 0.05 b |
| γ-Linolenic acid | C18:3n6 | 0.05 ± 0.03 b | 0.11 ± 0.01 b | 0.25 ± 0.15 ab | 0.45 ± 0.4 a | 0.1 ± 0.01 b | 0.05 ± 0 b |
| α-Linolenic acid (ALA) | C18:3n3 | 0.15 ± 0.09 a | 0.46 ± 0.03 a | 0.47 ± 0.42 a | 0.22 ± 0.15 a | 0.53 ± 0.35 a | 0.29 ± 0.01 a |
| Eicosadienoic acid | C20:2 | 0.32 ± 0.18 c | 0.36 ± 0.1 c | 0.39 ± 0.06 c | 0.32 ± 0.09 c | 0.99 ± 0.16 a | 0.75 ± 0.1 b |
| Eicosatetraenoic acid | C20:4 | 6.37 ± 0.05 a | 6.04 ± 0.58 ab | 3.63 ± 0.01 d | 4.88 ± 0.07 c | 4.94 ± 0.66 c | 5.44 ± 0.5 bc |
| Eicosapentaenoic acid (EPA) | C20:5n3 | 5.02 ± 0.02 b | 5.52 ± 0.48 b | 5.49 ± 0.03 b | 4.9 ± 0.22 b | 7.3 ± 0.88 a | 5.11 ± 0 b |
| Docosahexaenoic acid (DHA) | C22:6n3 | 33.83 ± 0.09 a | 22.94 ± 0.81 d | 23.7 ± 0.31 d | 28.67 ± 0.22 ab | 26.51 ± 0.87 ac | 28.82 ± 0.84 ab |
| PUFA | 47.6 ± 0.32 a | 37.17 ± 5.63 b | 43.85 ± 9.32 ab | 41.6 ± 0.44 ab | 41.98 ± 6.56 ab | 42.18 ± 0.29 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palanisamy, S.; Singh, A.; Zhang, B.; Zhao, Q.; Benjakul, S. Effects of Different Phenolic Compounds on the Redox State of Myoglobin and Prevention of Discoloration, Lipid and Protein Oxidation of Refrigerated Longtail Tuna (Thunnus tonggol) Slices. Foods 2024, 13, 1238. https://doi.org/10.3390/foods13081238
Palanisamy S, Singh A, Zhang B, Zhao Q, Benjakul S. Effects of Different Phenolic Compounds on the Redox State of Myoglobin and Prevention of Discoloration, Lipid and Protein Oxidation of Refrigerated Longtail Tuna (Thunnus tonggol) Slices. Foods. 2024; 13(8):1238. https://doi.org/10.3390/foods13081238
Chicago/Turabian StylePalanisamy, Suguna, Avtar Singh, Bin Zhang, Qiancheng Zhao, and Soottawat Benjakul. 2024. "Effects of Different Phenolic Compounds on the Redox State of Myoglobin and Prevention of Discoloration, Lipid and Protein Oxidation of Refrigerated Longtail Tuna (Thunnus tonggol) Slices" Foods 13, no. 8: 1238. https://doi.org/10.3390/foods13081238
APA StylePalanisamy, S., Singh, A., Zhang, B., Zhao, Q., & Benjakul, S. (2024). Effects of Different Phenolic Compounds on the Redox State of Myoglobin and Prevention of Discoloration, Lipid and Protein Oxidation of Refrigerated Longtail Tuna (Thunnus tonggol) Slices. Foods, 13(8), 1238. https://doi.org/10.3390/foods13081238