
Investigating Milk Fat Globule Structure, Size, and Functionality after Thermal Processing and Homogenization of Human Milk

 , , ,
, , , 
Abstract
:1. Introduction
2. Methods and Materials
2.1. Human Milk Handling, Storage, and Composition
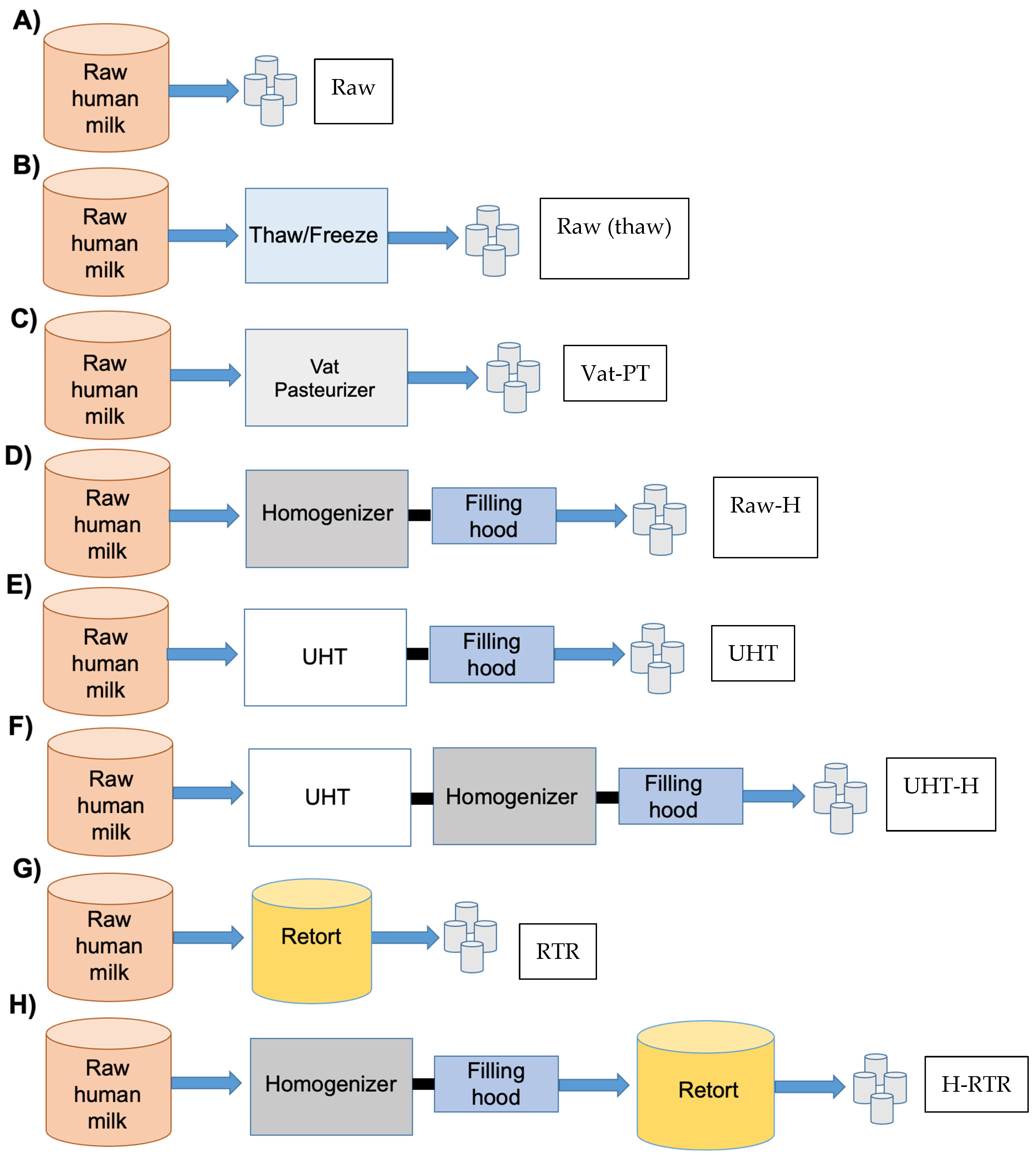
2.2. Human Milk Processing at the Industrial Scale
2.3. Milk Fat Globule Size Distribution
2.4. Zeta Potential of Milk Fat Globules
2.5. Microstructure of Milk Fat Globules
2.6. Color Parameters
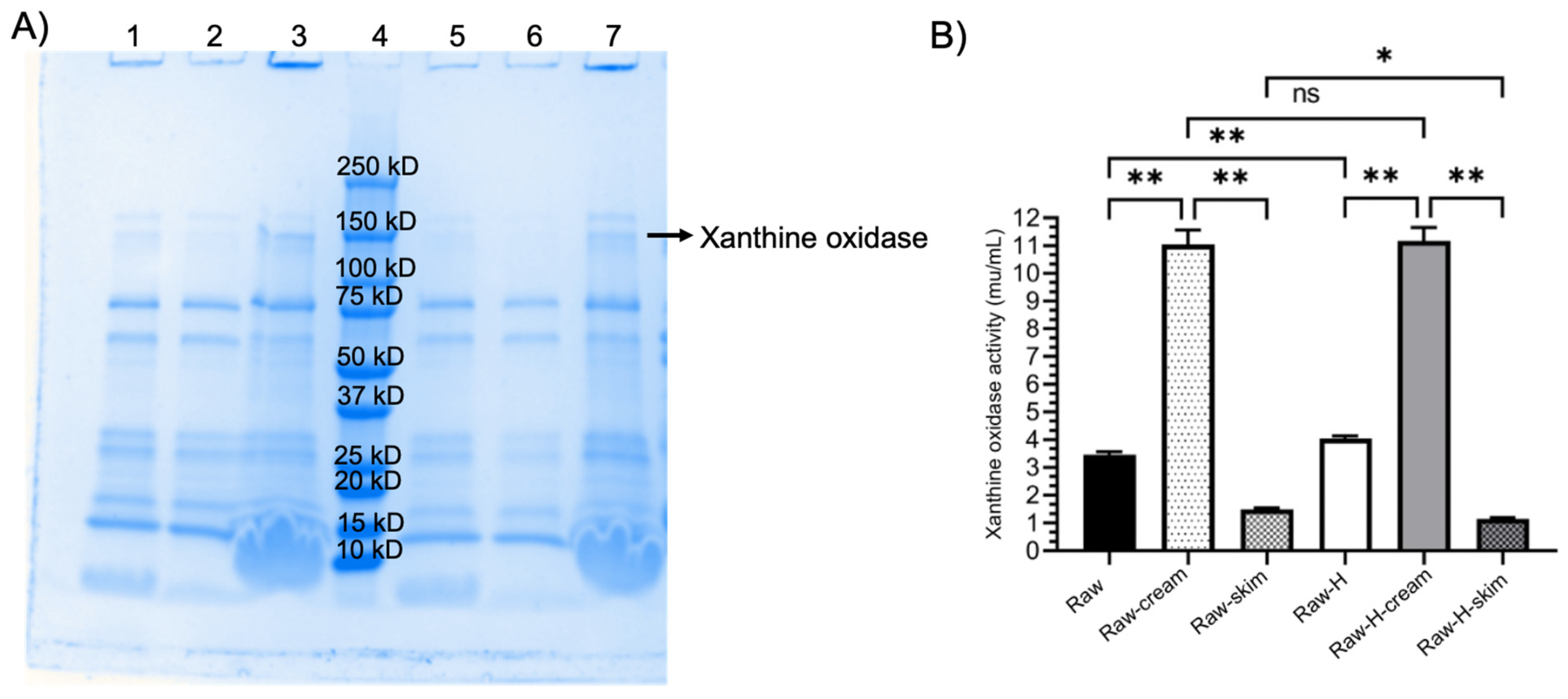
2.7. Xanthine Oxidase Activity
2.8. Distribution of MFGM Proteins in Fractions of Homogenized and Non-Homogenized Raw Human Milk by SDS-PAGE
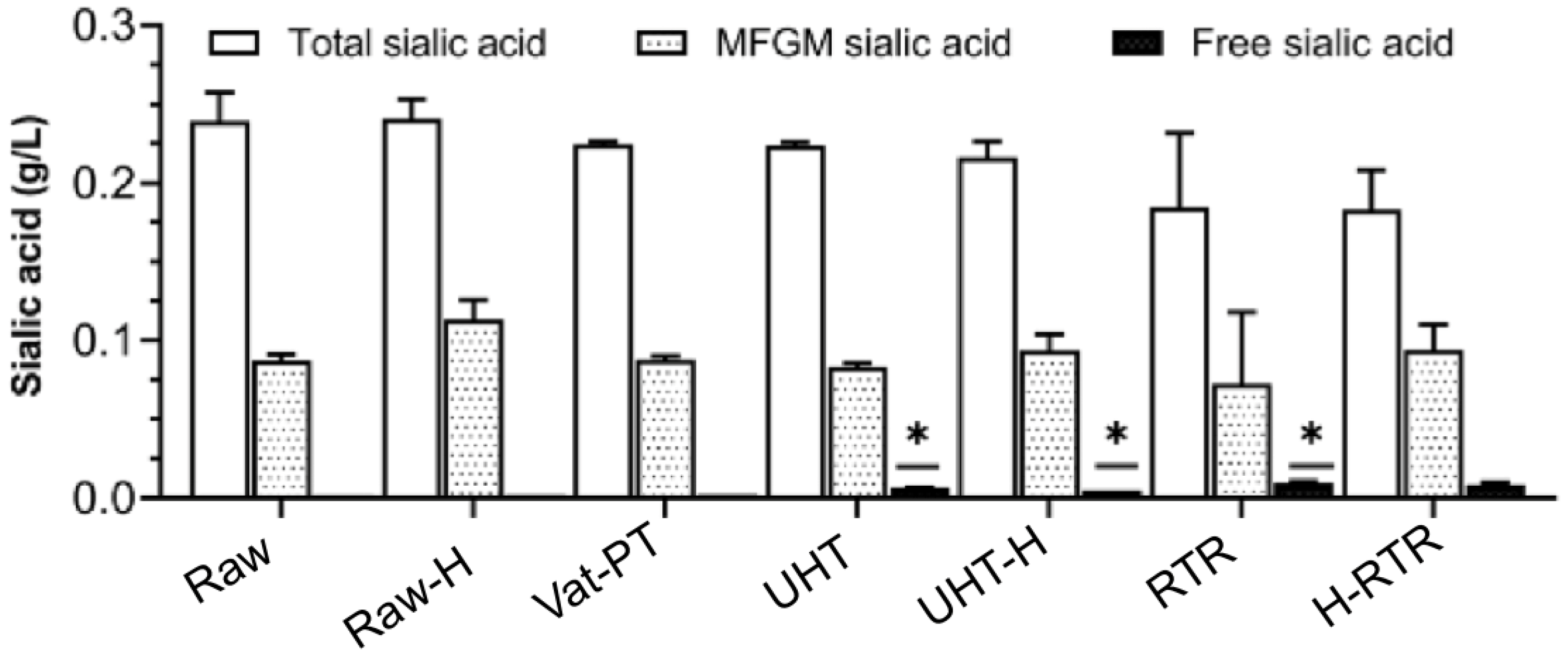
2.9. Sialic Acid Quantification
2.10. Lipolysis of Milk Fat Globules by the Pancreatic Lipase
3. Statistical Analysis
4. Results
4.1. Human Milk Composition
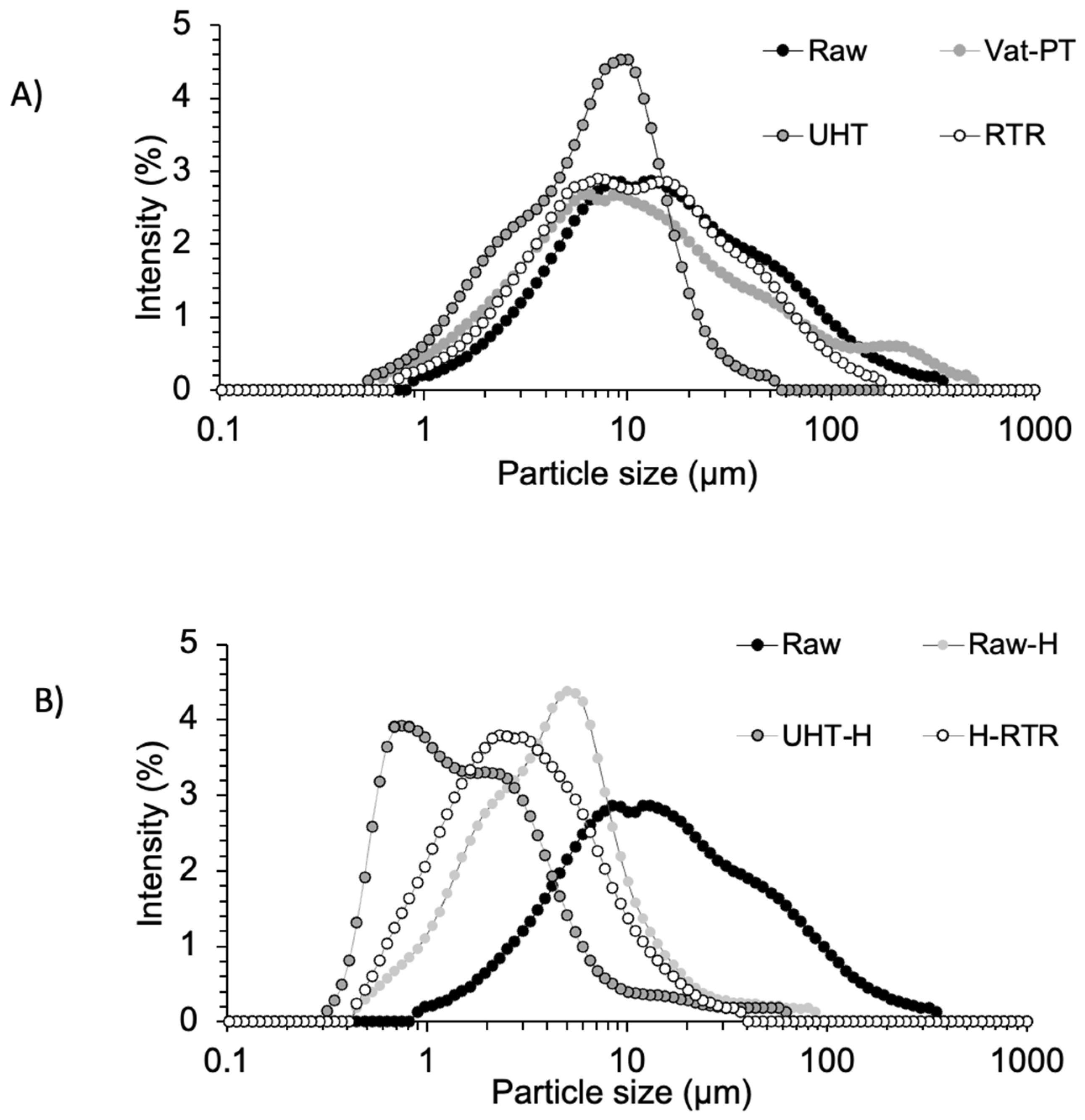
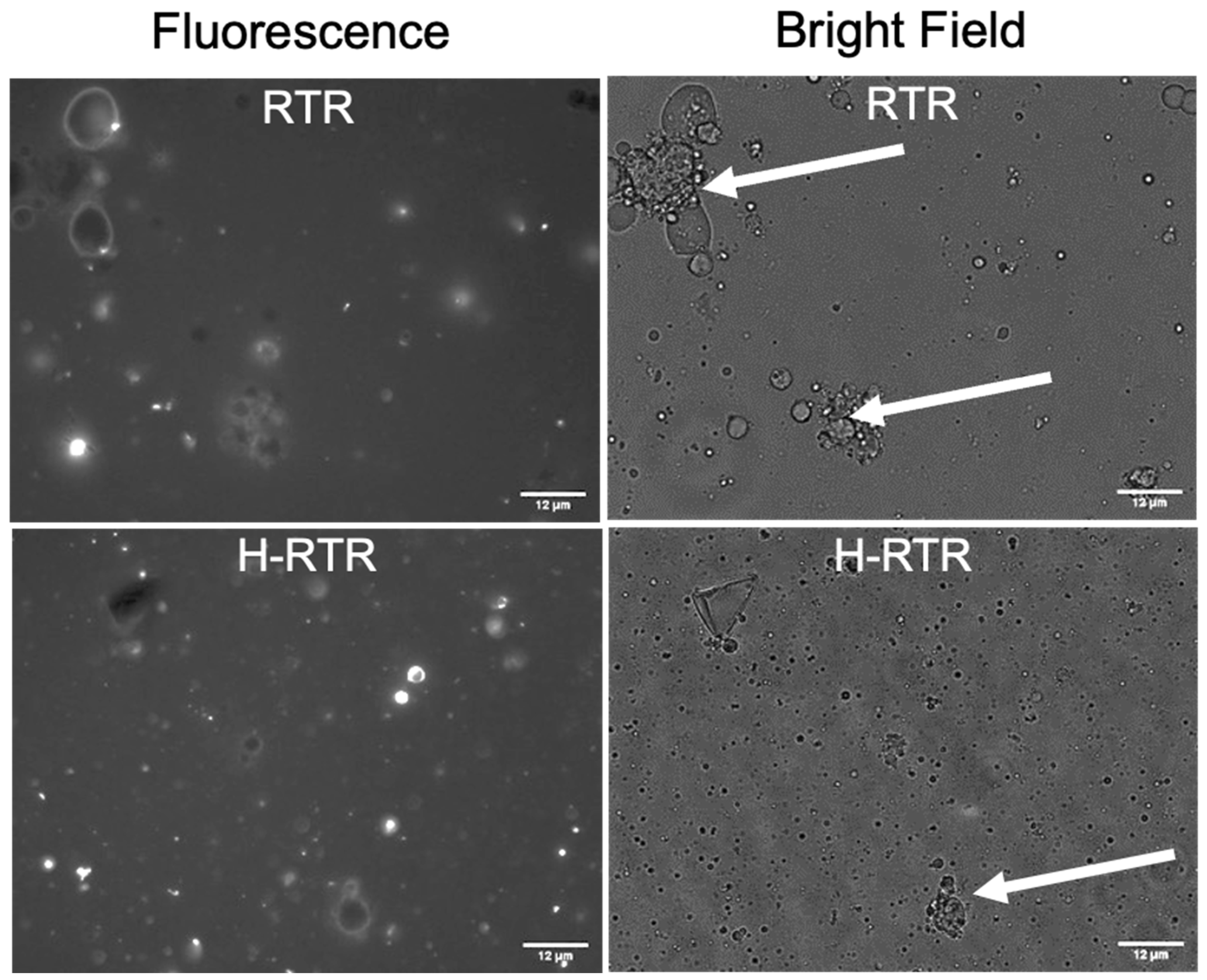
4.2. Changes in MFG Particle Size Distribution, Zeta Potential, and Membrane Induced by Homogenization and Thermal Processing
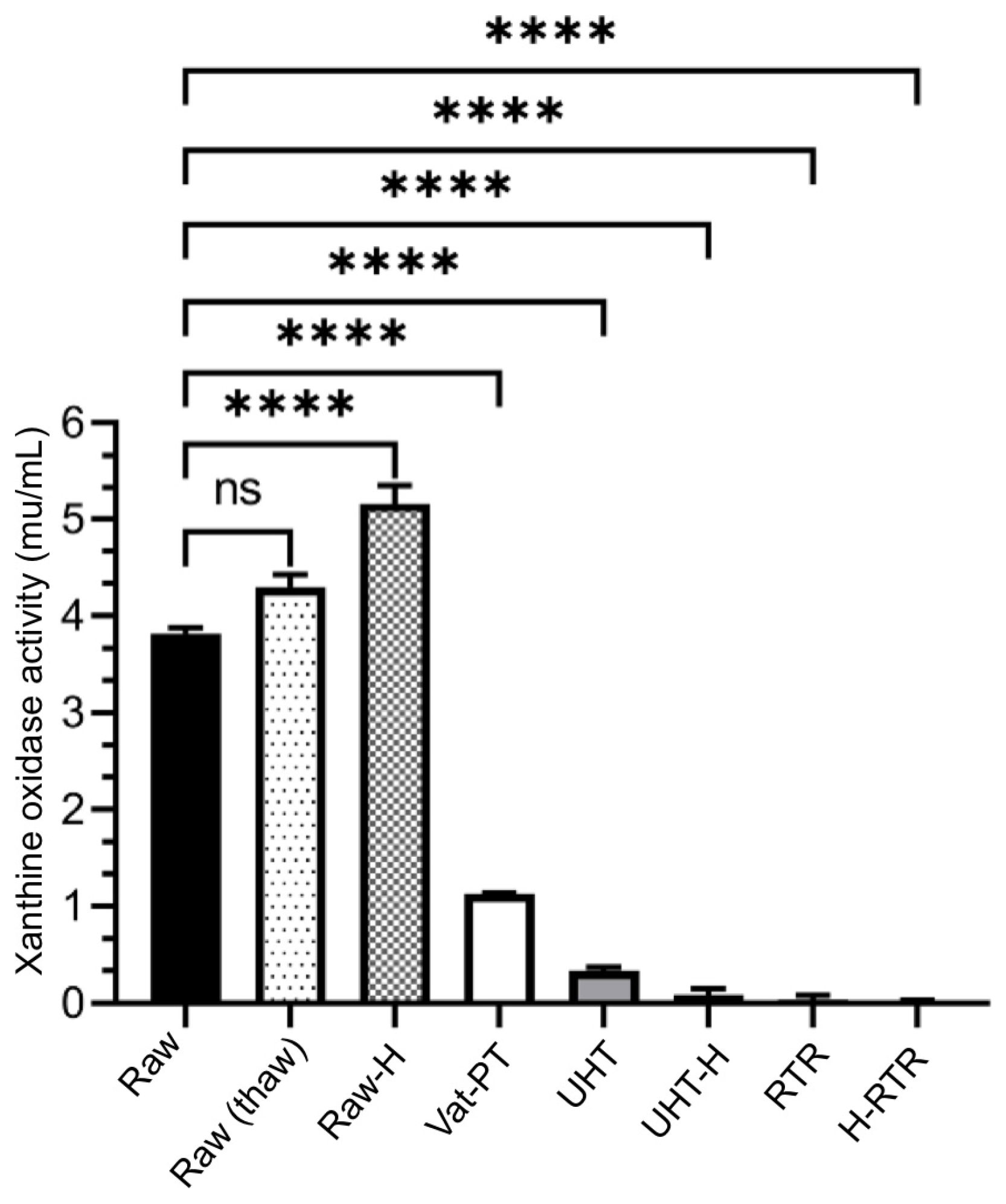
4.3. Xanthine Oxidase Activity
4.4. Distribution of MFGM Proteins in Fractions of Homogenized and Non-Homogenized Human Milk
4.5. Sialic Acid
4.6. Color Parameters
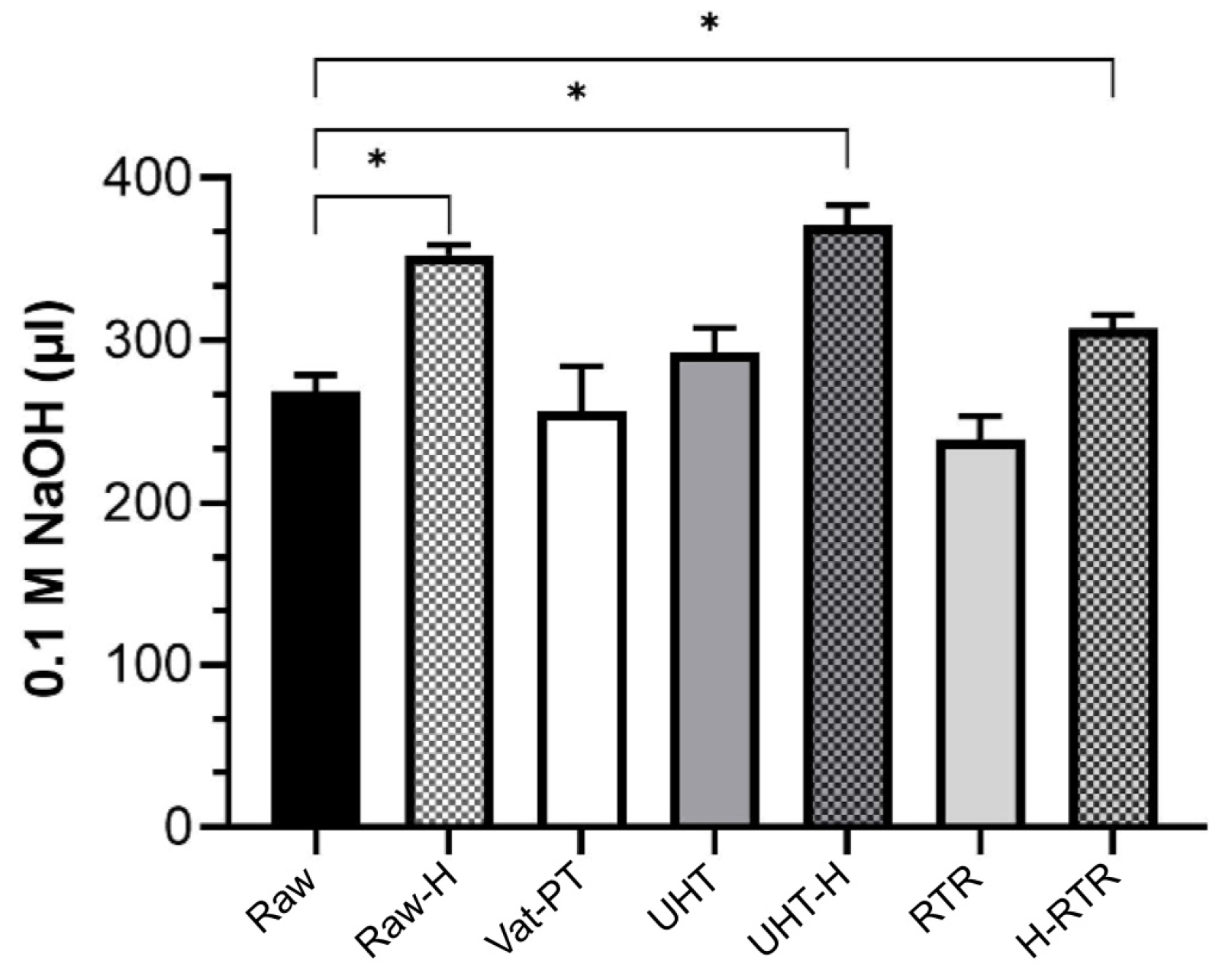
4.7. Lipolysis of Milk Fat Globules by the Pancreatic Lipase
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hinde, K.; German, J.B. Food in an Evolutionary Context: Insights from Mother’s Milk. J. Sci. Food Agric. 2012, 92, 2219–2223. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Human Milk: Maternal Dietary Lipids and Infant Development. Proc. Nutr. Soc. 2007, 66, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Heid, H.W.; Keenan, T.W. Intracellular Origin and Secretion of Milk Fat Globules. Eur. J. Cell Biol. 2005, 84, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Brink, L.R.; Lönnerdal, B. The Role of Milk Fat Globule Membranes in Behavior and Cognitive Function Using a Suckling Rat Pup Supplementation Model. J. Nutr. Biochem. 2018, 58, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Dewettinck, K.; Rombaut, R.; Thienpont, N.; Le, T.T.; Messens, K.; Van Camp, J. Nutritional and Technological Aspects of Milk Fat Globule Membrane Material. Int. Dairy J. 2008, 18, 436–457. [Google Scholar] [CrossRef]
- Fong, B.Y.; Norris, C.S.; MacGibbon, A.K.H. Protein and Lipid Composition of Bovine Milk-Fat-Globule Membrane. Int. Dairy J. 2007, 17, 275–288. [Google Scholar] [CrossRef]
- Lee, H.; Padhi, E.; Hasegawa, Y.; Larke, J.; Parenti, M.; Wang, A.; Hernell, O.; Lönnerdal, B.; Slupsky, C. Compositional Dynamics of the Milk Fat Globule and Its Role in Infant Development. Front. Pediatr. 2018, 6, 313. [Google Scholar] [CrossRef] [PubMed]
- Mather, I.H. A Review and Proposed Nomenclature for Major Proteins of the Milk-Fat Globule Membrane. J. Dairy Sci. 2000, 83, 203–247. [Google Scholar] [CrossRef] [PubMed]
- Al-Shehri, S.S.; Knox, C.L.; Liley, H.G.; Cowley, D.M.; Wright, J.R.; Henman, M.G.; Hewavitharana, A.K.; Charles, B.G.; Shaw, P.N.; Sweeney, E.L.; et al. Breastmilk-Saliva Interactions Boost Innate Immunity by Regulating the Oral Microbiome in Early Infancy. PLoS ONE 2015, 10, e0135047. [Google Scholar] [CrossRef]
- Ozturk, G.; Shah, I.M.; Mills, D.A.; German, J.B.; de Moura Bell, J.M.L.N. The Antimicrobial Activity of Bovine Milk Xanthine Oxidase. Int. Dairy J. 2020, 102, 104581. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; McVeagh, P.; Petocz, P.; Brand-Miller, J. Brain Ganglioside and Glycoprotein Sialic Acid in Breastfed Compared with Formula-Fed Infants. Am. J. Clin. Nutr. 2003, 78, 1024–1029. [Google Scholar] [CrossRef] [PubMed]
- Garrido, D.; Nwosu, C.; Ruiz-Moyano, S.; Aldredge, D.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Endo-β-N-Acetylglucosaminidases from Infant Gut-Associated Bifidobacteria Release Complex N-Glycans from Human Milk Glycoproteins. Mol. Cell. Proteom. 2012, 11, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Perrine, C.G.; Scanlon, K.S. Prevalence of Use of Human Milk in US Advanced Care Neonatal Units. Pediatrics 2013, 131, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Bylund, G. Dairy Processing Handbook; Tetra Pak Processing Systems: Lund, Sweden, 2003; ISBN 978-91-631-3427-2. [Google Scholar]
- Rowe, M.; Donaghy, J. Microbiological Aspects of Dairy Ingredients. In Dairy Ingredients for Food Processing; Chandan, R.C., Kilara, A., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 59–101. ISBN 978-0-470-95916-9. [Google Scholar]
- Chandan, R.C. Dairy Ingredients for Food Processing: An Overview. In Dairy Ingredients for Food Processing; Chandan, R.C., Kilara, A., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 3–33. ISBN 978-0-470-95916-9. [Google Scholar]
- Arslanoglu, S.; Bertino, E.; Tonetto, P.; De Nisi, G.; Ambruzzi, A.M.; Biasini, A.; Profeti, C.; Spreghini, M.R.; Moro, G.E. Guidelines for the Establishment and Operation of a Donor Human Milk Bank. J. Matern.-Fetal Neonatal Med. 2010, 23, 1–20. [Google Scholar] [CrossRef]
- Peila, C.; Moro, G.E.; Bertino, E.; Cavallarin, L.; Giribaldi, M.; Giuliani, F.; Cresi, F.; Coscia, A. The Effect of Holder Pasteurization on Nutrients and Biologically-Active Components in Donor Human Milk: A Review. Nutrients 2016, 8, 477. [Google Scholar] [CrossRef] [PubMed]
- Centre for Clinical Practice at NICE (UK). Donor Breast Milk Banks: The Operation of Donor Milk Bank Services; National Institute for Health and Clinical Excellence: Guidance; National Institute for Health and Clinical Excellence (UK): London, UK, 2010. [Google Scholar]
- Meredith-Dennis, L.; Xu, G.; Goonatilleke, E.; Lebrilla, C.B.; Underwood, M.A.; Smilowitz, J.T. Composition and Variation of Macronutrients, Immune Proteins, and Human Milk Oligosaccharides in Human Milk From Nonprofit and Commercial Milk Banks. J. Hum. Lact. 2018, 34, 120–129. [Google Scholar] [CrossRef] [PubMed]
- About Us. Available online: https://www.ni-q.com/about-us/ (accessed on 24 July 2023).
- Bertino, E.; Coppa, G.V.; Giuliani, F.; Coscia, A.; Gabrielli, O.; Sabatino, G.; Sgarrella, M.; Testa, T.; Zampini, L.; Fabris, C. Effects of Holder Pasteurization on Human Milk Oligosaccharides. Int. J. Immunopathol. Pharmacol. 2008, 21, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lee, N.A.; Duley, J.A.; Cowley, D.M.; Shaw, P.N.; Bansal, N. Comparing the Effects of Hydrostatic High-Pressure Processing vs Holder Pasteurisation on the Microbial, Biochemical and Digestion Properties of Donor Human Milk. Food Chem. 2022, 373, 131545. [Google Scholar] [CrossRef] [PubMed]
- Leite, J.A.S.; Robinson, R.C.; Salcedo, J.; Ract, J.N.R.; Quintal, V.S.; Tadini, C.C.; Barile, D. The Effect of Microwave-Assisted Heating on Bioactive and Immunological Compounds in Donor Human Milk. LWT 2022, 161, 113306. [Google Scholar] [CrossRef]
- Young, B.E.; Murphy, K.; Borman, L.L.; Heinrich, R.; Krebs, N.F. Milk Bank Pooling Practices Impact Concentrations and Variability of Bioactive Components of Donor Human Milk. Front. Nutr. 2020, 7, 579115. [Google Scholar] [CrossRef] [PubMed]
- Moltó-Puigmartí, C.; Permanyer, M.; Castellote, A.I.; López-Sabater, M.C. Effects of Pasteurisation and High-Pressure Processing on Vitamin C, Tocopherols and Fatty Acids in Mature Human Milk. Food Chem. 2011, 124, 697–702. [Google Scholar] [CrossRef]
- de Oliveira, S.C.; Bellanger, A.; Ménard, O.; Pladys, P.; Gouar, Y.L.; Henry, G.; Dirson, E.; Rousseau, F.; Carrière, F.; Dupont, D.; et al. Impact of Homogenization of Pasteurized Human Milk on Gastric Digestion in the Preterm Infant: A Randomized Controlled Trial. Clin. Nutr. ESPEN 2017, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, S.C.; Deglaire, A.; Ménard, O.; Bellanger, A.; Rousseau, F.; Henry, G.; Dirson, E.; Carrière, F.; Dupont, D.; Bourlieu, C. Holder Pasteurization Impacts the Proteolysis, Lipolysis and Disintegration of Human Milk under in Vitro Dynamic Term Newborn Digestion. Food Res. Int. 2016, 88, 263–275. [Google Scholar] [CrossRef]
- HMBANA Publications. Available online: https://www.hmbana.org/about-us/our-work/publications.html (accessed on 11 January 2024).
- AOAC 972.16-1972(1996); Fat, Lactose, Protein, and Solids in Milk—$14.75: AOAC Official Method. 1996. Available online: http://www.aoacofficialmethod.org/index.php?main_page=product_info&cPath=1&products_id=38 (accessed on 27 January 2024).
- Michalski, M.-C.; Briard, V.; Michel, F. Optical Parameters of Milk Fat Globules for Laser Light Scattering Measurements. Lait 2001, 81, 787–796. [Google Scholar] [CrossRef]
- Alshehab, M.; Reis, M.G.; Day, L.; Nitin, N. Milk Fat Globules, a Novel Carrier for Delivery of Exogenous Cholecalciferol. Food Res. Int. 2019, 126, 108579. [Google Scholar] [CrossRef] [PubMed]
- McGuire, R.G. Reporting of Objective Color Measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef]
- Milovanovic, B.; Djekic, I.; Miocinovic, J.; Djordjevic, V.; Lorenzo, J.M.; Barba, F.J.; Mörlein, D.; Tomasevic, I. What Is the Color of Milk and Dairy Products and How Is It Measured? Foods 2020, 9, 1629. [Google Scholar] [CrossRef] [PubMed]
- Hurum, D.C.; Rohrer, J.S. Determination of Sialic Acids in Infant Formula by Chromatographic Methods: A Comparison of High-Performance Anion-Exchange Chromatography with Pulsed Amperometric Detection and Ultra-High-Performance Liquid Chromatography Methods. J. Dairy Sci. 2012, 95, 1152–1161. [Google Scholar] [CrossRef] [PubMed]
- Grundy, M.M.L.; Abrahamse, E.; Almgren, A.; Alminger, M.; Andres, A.; Ariëns, R.M.C.; Bastiaan-Net, S.; Bourlieu-Lacanal, C.; Brodkorb, A.; Bronze, M.R.; et al. INFOGEST Inter-Laboratory Recommendations for Assaying Gastric and Pancreatic Lipases Activities Prior to in Vitro Digestion Studies. J. Funct. Foods 2021, 82, 104497. [Google Scholar] [CrossRef]
- Berton, A.; Rouvellac, S.; Robert, B.; Rousseau, F.; Lopez, C.; Crenon, I. Effect of the Size and Interface Composition of Milk Fat Globules on Their in Vitro Digestion by the Human Pancreatic Lipase: Native versus Homogenized Milk Fat Globules. Food Hydrocoll. 2012, 29, 123–134. [Google Scholar] [CrossRef]
- McClements, D.J.; Li, Y. Structured Emulsion-Based Delivery Systems: Controlling the Digestion and Release of Lipophilic Food Components. Adv. Colloid Interface Sci. 2010, 159, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.W.; Moon, T.-W.; Dylewski, D.P. Lipid Globules Retain Globule Membrane Material after Homogenization. J. Dairy Sci. 1983, 66, 196–203. [Google Scholar] [CrossRef]
- Michalski, M.C.; Briard, V.; Michel, F.; Tasson, F.; Poulain, P. Size Distribution of Fat Globules in Human Colostrum, Breast Milk, and Infant Formula. J. Dairy Sci. 2005, 88, 1927–1940. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Ménard, O. Human Milk Fat Globules: Polar Lipid Composition and in Situ Structural Investigations Revealing the Heterogeneous Distribution of Proteins and the Lateral Segregation of Sphingomyelin in the Biological Membrane. Colloids Surf. B Biointerfaces 2011, 83, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Nebbia, S.; Giribaldi, M.; Cavallarin, L.; Bertino, E.; Coscia, A.; Briard-Bion, V.; Ossemond, J.; Henry, G.; Ménard, O.; Dupont, D.; et al. Differential Impact of Holder and High Temperature Short Time Pasteurization on the Dynamic in Vitro Digestion of Human Milk in a Preterm Newborn Model. Food Chem. 2020, 328, 127126. [Google Scholar] [CrossRef] [PubMed]
- Dhungana, P.; Truong, T.; Bansal, N.; Bhandari, B. Apparent Thermal and UHT Stability of Native, Homogenized and Recombined Creams with Different Average Fat Globule Sizes. Food Res. Int. 2019, 123, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Wiking, L.; Gregersen, S.B.; Hansen, S.F.; Hammershøj, M. Heat-Induced Changes in Milk Fat and Milk Fat Globules and Its Derived Effects on Acid Dairy Gelation—A Review. Int. Dairy J. 2022, 127, 105213. [Google Scholar] [CrossRef]
- Liang, N.; Koh, J.; Kim, B.J.; Ozturk, G.; Barile, D.; Dallas, D.C. Structural and Functional Changes of Bioactive Proteins in Donor Human Milk Treated by Vat-Pasteurization, Retort Sterilization, Ultra-High-Temperature Sterilization, Freeze-Thawing and Homogenization. Front. Nutr. 2022, 9, 926814. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Koh, J.; Liang, N.; Yang, J.; Ozturk, G.; Barile, D.; Dallas, D.C. Effect of Vat Pasteurization, Ultra-High Temperature Sterilization, Retort Sterilization and Homogenization on Soluble Proteins in Donor Human Milk Detected via Proteomics. LWT 2023, 182, 114842. [Google Scholar] [CrossRef]
- Jukkola, A.; Hokkanen, S.; Kämäräinen, T.; Partanen, R.; Heino, A.; Rojas, O.J. Changes in Milk Fat Globules and Membrane Lipids under the Shear Fields of Microfiltration and Centrifugation. J. Membr. Sci. 2019, 573, 218–225. [Google Scholar] [CrossRef]
- Van Boekel, M.A.J.S. Effect of Heating on Maillard Reactions in Milk. Food Chem. 1998, 62, 403–414. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, L.; Tian, S.; Li, X.; Hussain, M.; Li, C.; Zhang, L.; Zhang, Q.; Leng, Y.; Jiang, S.; et al. Comparative Analysis of Interfacial Composition and Structure of Fat Globules in Human Milk and Infant Formulas. Food Hydrocoll. 2022, 124, 107290. [Google Scholar] [CrossRef]
- Zhao, L.; Du, M.; Mao, X. Change in Interfacial Properties of Milk Fat Globules by Homogenization and Thermal Processing Plays a Key Role in Their in Vitro Gastrointestinal Digestion. Food Hydrocoll. 2019, 96, 331–342. [Google Scholar] [CrossRef]
- Pan, Y.; Liu, Y.; Zhao, J.; Cui, L.; Li, X.; Liu, L.; Kouame, K.J.E.-P.; Wang, Z.; Tan, X.; Jiang, Y.; et al. Simulated in Vitro Infant Digestion and Lipidomic Analysis to Explore How the Milk Fat Globule Membrane Modulates Fat Digestion. Food Chem. 2024, 447, 139008. [Google Scholar] [CrossRef] [PubMed]
- Bourlieu, C.; Ménard, O.; De La Chevasnerie, A.; Sams, L.; Rousseau, F.; Madec, M.-N.; Robert, B.; Deglaire, A.; Pezennec, S.; Bouhallab, S.; et al. The Structure of Infant Formulas Impacts Their Lipolysis, Proteolysis and Disintegration during in Vitro Gastric Digestion. Food Chem. 2015, 182, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Liu, S.; Zhang, X.; Li, X.; Liu, L.; Hao, D.; Cui, L.; Ma, C.; Dang, X.; Xu, Y.; et al. Influence of Pasteurization and Spray Drying on the Fat Digestion Behavior of Human Milk Fat Analog Emulsion: A Simulated in Vitro Infant Digestion Study. J. Sci. Food Agric. 2024. Epub ahead of printing. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Diameter (μm) | Vol % | Width | Zeta Potential (mV) |
|---|---|---|---|---|
| Raw | 13.55 ± 0.10 | 100 | 44.94 ± 1.32 | −16.00 ± 0.50 |
| Vat-PT | 9.79 ± 0.38 | 100 | 38.83 ± 5.37 | −16.50 ± 0.20 |
| UHT | 7.13 ± 0.34 | 100 | 11.55 ± 0.50 | −24.60 ± 0.90 |
| RTR | 10.79 ± 0.78 | 100 | 27.76 ± 2.61 | −20.10 ± 0.20 |
| Raw-H | 3.90 ± 0.04 | 100 | 7.74 ± 0.54 | −23.10 ± 0.30 |
| UHT-H | 2.06 ± 0.44 | 100 | 4.86 ± 0.85 | −24.40 ± 0.50 |
| H-RTR | 3.90 ± 1.40 | 100 | 9.41 ± 4.54 | −21.20 ± 0.20 |
| Samples | L* | a* | b* |
|---|---|---|---|
| Raw | 80.94 ± 0.06 **** | −4.15 ± 0.00 **** | 9.76 ± 0.07 **** |
| Raw (thaw) | 81.42 ± 0.04 **** | −3.89 ± 0.01 **** | 10.16 ± 0.12 **** |
| Raw-H | 86.72 ± 0.01 **** | −3.00 ± 0.00 **** | 9.68 ± 0.01 ns |
| Vat-PT | 80.06 ± 0.04 **** | −3.66 ± 0.01 **** | 8.64 ± 0.11 **** |
| UHT | 82.71 ± 0.64 **** | −3.68 ± 0.00 **** | 10.79 ± 0.63 **** |
| UHT-H | 87.56 ± 0.25 **** | −3.00 ±0.00 **** | 9.27 ± 0.13 **** |
| RTR | 82.23 ± 2.81 **** | −2.29 ± 0.00 **** | 15.07 ± 0.66 **** |
| H-RTR | 85.11 ± 1.08 **** | −1.66 ± 0.04 **** | 13.27 ± 0.37 **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozturk, G.; Paviani, B.; Rai, R.; Robinson, R.C.; Durham, S.D.; Baller, M.I.; Wang, A.; Nitin, N.; Barile, D. Investigating Milk Fat Globule Structure, Size, and Functionality after Thermal Processing and Homogenization of Human Milk. Foods 2024, 13, 1242. https://doi.org/10.3390/foods13081242
Ozturk G, Paviani B, Rai R, Robinson RC, Durham SD, Baller MI, Wang A, Nitin N, Barile D. Investigating Milk Fat Globule Structure, Size, and Functionality after Thermal Processing and Homogenization of Human Milk. Foods. 2024; 13(8):1242. https://doi.org/10.3390/foods13081242
Chicago/Turabian StyleOzturk, Gulustan, Bruna Paviani, Rewa Rai, Randall C. Robinson, Sierra D. Durham, Mara I. Baller, Aidong Wang, Nitin Nitin, and Daniela Barile. 2024. "Investigating Milk Fat Globule Structure, Size, and Functionality after Thermal Processing and Homogenization of Human Milk" Foods 13, no. 8: 1242. https://doi.org/10.3390/foods13081242
APA StyleOzturk, G., Paviani, B., Rai, R., Robinson, R. C., Durham, S. D., Baller, M. I., Wang, A., Nitin, N., & Barile, D. (2024). Investigating Milk Fat Globule Structure, Size, and Functionality after Thermal Processing and Homogenization of Human Milk. Foods, 13(8), 1242. https://doi.org/10.3390/foods13081242