Comparative Phytochemical Analyses and Metabolic Profiling of Different Phenotypes of Chinese Cabbage (Brassica Rapa ssp. Pekinensis)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. GC-TOFMS Analysis
2.3. Glucosinolate Extraction and HPLC Analysis
2.4. Phenolic Compound Extraction and HPLC Analysis
2.5. Carotenoid Extraction and HPLC Analysis
2.6. Chlorophyll Contents Analysis
2.7. Statistical Analysis
3. Results
3.1. Morphological Variation between Oval and Rectangular Chinese Cabbage
3.2. Metabolite-Specific Profiling in Oval and Rectangular Chinese Cabbage
3.3. Glucosinolates in the Oval and Rectangular Chinese Cabbage
3.4. Phenolics in the Oval and Rectangular Chinese Cabbage
3.5. Carotenoids and Chlorophylls in the Oval and Rectangular Chinese Cabbage
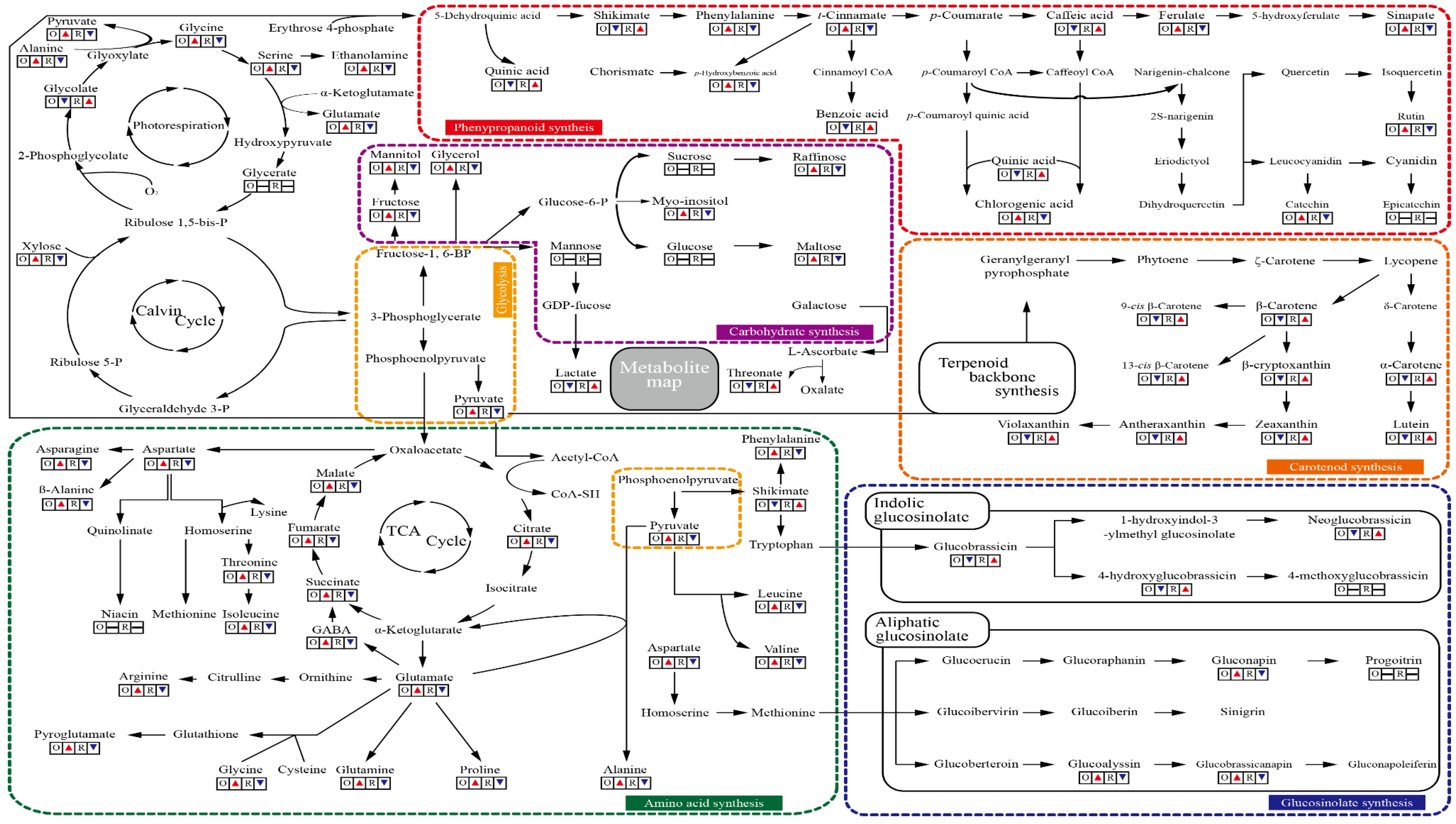
3.6. A metabolite Map Comparing Primary and Secondary Metabolites in Oval and Rectangular Chinese Cabbage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kim, Y.; Li, X.; Kim, S.-J.; Kim, H.; Lee, J.; Kim, H.; Park, S. MYB transcription factors regulate glucosinolate biosynthesis in different organs of Chinese cabbage (Brassica rapa ssp. pekinensis). Molecules 2013, 18, 8682–8695. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, M.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Health-affecting compounds in Brassicaceae. Compr. Rev. Food Sci. Food Saf. 2009, 8, 31–43. [Google Scholar] [CrossRef]
- Björkman, M.; Klingen, I.; Birch, A.N.; Bones, A.M.; Bruce, T.J.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E. Phytochemicals of Brassicaceae in plant protection and human health—Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Al-Gendy, A.; El-Gindi, O.; Hafez, A.S.; Ateya, A. Glucosinolates, volatile constituents and biological activities of Erysimum corinthium Boiss. (Brassicaceae). Food Chem. 2010, 118, 519–524. [Google Scholar] [CrossRef]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Kljak, K.; Grbeša, D. Carotenoid content and antioxidant activity of hexane extracts from selected Croatian corn hybrids. Food Chem. 2015, 167, 402–408. [Google Scholar] [CrossRef]
- Howitt, C.A.; Pogson, B.J. Carotenoid accumulation and function in seeds and non-green tissues. Plant Cell Environ. 2006, 29, 435–445. [Google Scholar] [CrossRef]
- Koornneef, M. Genetic aspects of abscisic acid. In A Genetic Approach to Plant Biochemistry; Blonstein, A.D., King, P.J., Eds.; Springer: Vienna, Austria, 1986; pp. 35–54. [Google Scholar]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Baek, S.-A.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of Lycoris radiata. Biology 2019, 8, 63. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, N.S.; Park, J.S.; Lee, S.Y.; Lee, J.-W.; Park, S.U. Effects of light-emitting diodes on the accumulation of glucosinolates and phenolic compounds in sprouting canola (Brassica napus L.). Foods 2019, 8, 76. [Google Scholar] [CrossRef]
- Lee, M.-K.; Chun, J.-H.; Byeon, D.H.; Chung, S.-O.; Park, S.U.; Park, S.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, Y.-P.; Kim, S.-J. Variation of glucosinolates in 62 varieties of Chinese cabbage (Brassica rapa L. ssp. pekinensis) and their antioxidant activity. LWT Food Sci. Technol. 2014, 58, 93–101. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, Y.B.; Li, X.; Choi, S.R.; Park, S.; Park, J.S.; Lim, Y.P.; Park, S.U. Accumulation of phenylpropanoids by white, blue, and red light irradiation and their organ-specific distribution in Chinese cabbage (Brassica rapa ssp. pekinensis). J. Agric. Food Chem. 2015, 63, 6772–6778. [Google Scholar] [CrossRef] [PubMed]
- Tuan, P.A.; Kim, J.K.; Lee, J.; Park, W.T.; Kwon, D.Y.; Kim, Y.B.; Kim, H.H.; Kim, H.R.; Park, S.U. Analysis of carotenoid accumulation and expression of carotenoid biosynthesis genes in different organs of Chinese cabbage (Brassica rapa subsp. pekinensis). EXCLI J. 2012, 11, 508–516. [Google Scholar] [PubMed]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4-3. [Google Scholar] [CrossRef]
- Chun, J.-H.; Kim, N.-H.; Seo, M.-S.; Jin, M.; Park, S.U.; Arasu, M.V.; Kim, S.-J.; Al-Dhabi, N.A. Molecular characterization of glucosinolates and carotenoid biosynthetic genes in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Saudi J. Biol. Sci. 2018, 25, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Chu, S.M.; Kim, S.J.; Lee, D.J.; Lee, S.Y.; Lim, S.H.; Ha, S.-H.; Kweon, S.J.; Cho, H.S. Variation of glucosinolates in vegetable crops of Brassica rapa L. ssp. pekinensis. Food Chem. 2010, 119, 423–428. [Google Scholar] [CrossRef]
- Baek, S.-A.; Jung, Y.-H.; Lim, S.-H.; Park, S.U.; Kim, J.K. Metabolic profiling in Chinese cabbage (Brassica rapa L. subsp. pekinensis) cultivars reveals that glucosinolate content is correlated with carotenoid content. J. Agric. Food Chem. 2016, 64, 4426–4434. [Google Scholar] [CrossRef]
- Watanabe, M.; Musumi, K.; Ayugase, J. Carotenoid pigment composition, polyphenol content, and antioxidant activities of extracts from orange-colored Chinese cabbage. LWT Food Sci. Technol. 2011, 44, 1971–1975. [Google Scholar] [CrossRef]
- Jiang, N.; Chung, S.-O.; Lee, J.; Ryu, D.; Lim, Y.P.; Park, S.; Lee, C.; Song, J.; Kim, K.; Park, J.-T. Increase of phenolic compounds in new Chinese cabbage cultivar with red phenotype. Hortic. Environ. Biotechnol. 2013, 54, 82–88. [Google Scholar] [CrossRef]
- Lee, H.; Oh, I.-N.; Kim, J.; Jung, D.; Cuong, N.P.; Kim, Y.; Lee, J.; Kwon, O.; Park, S.U.; Lim, Y. Phenolic compound profiles and their seasonal variations in new red-phenotype head-forming Chinese cabbages. LWT Food Sci. Technol. 2018, 90, 433–439. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Baskar, T.B.; Park, Y.E.; Park, J.S.; Lee, S.Y.; Park, S.U. In vitro antioxidant and antimicrobial properties of flower, leaf, and stem extracts of Korean mint. Antioxidants 2019, 8, 75. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Park, S.-Y.; Lee, S.Y.; Kim, J.K.; Park, S.U. Analysis of metabolites in white flowers of Magnolia denudata Desr. and violet flowers of Magnolia liliiflora Desr. Molecules 2018, 23, 1558. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Park, C.H.; Yang, J.; Yeo, H.J.; Kim, T.J.; Kim, J.K.; Park, S.U. Molecular characterization of anthocyanin and betulinic acid biosynthesis in red and white mulberry fruits using high-throughput sequencing. Food Chem. 2019, 279, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Morgan, A.M.A.; Park, B.B.; Lee, S.Y.; Lee, S.; Kim, J.K.; Park, S.U. Metabolic analysis of four cultivars of Liriope platyphylla. Metabolites 2019, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental abiotic factors on the content of saponins in plants. Phytochem. Rev. 2011, 10, 471–491. [Google Scholar] [CrossRef]
- Guo, L.; Wang, S.; Zhang, J.; Yang, G.; Zhao, M.; Ma, W.; Zhang, X.; Li, X.; Han, B.; Chen, N. Effects of ecological factors on secondary metabolites and inorganic elements of Scutellaria baicalensis and analysis of geoherblism. Sci. China Life Sci. 2013, 56, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Markom, M.; Hasan, M.; Daud, W.R.W.; Singh, H.; Jahim, J.M. Extraction of hydrolysable tannins from Phyllanthus niruri Linn.: Effects of solvents and extraction methods. Sep. Purif. Technol. 2007, 52, 487–496. [Google Scholar] [CrossRef]
- Zulak, K.G.; Weljie, A.M.; Vogel, H.J.; Facchini, P.J. Quantitative 1 H NMR metabolomics reveals extensive metabolic reprogramming of primary and secondary metabolism in elicitor-treated opium poppy cell cultures. BMC Plant Biol. 2008, 8, 5. [Google Scholar] [CrossRef]
- Zheng, Z.L. Carbon and nitrogen nutrient balance signaling in plants. Plant Signal. Behav. 2009, 4, 584–591. [Google Scholar] [CrossRef]
- Aharoni, A.; Galili, G. Metabolic engineering of the plant primary–secondary metabolism interface. Curr. Opin. Biotechnol. 2011, 22, 239–244. [Google Scholar] [CrossRef]
- Zakhleniuk, O.V.; Raines, C.A.; Lloyd, J.C. pho3: A phosphorus-deficient mutant of Arabidopsis thaliana (L.) Heynh. Planta 2001, 212, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Ferri, M.; Righetti, L.; Tassoni, A. Increasing sucrose concentrations promote phenylpropanoid biosynthesis in grapevine cell cultures. J. Plant Physiol. 2011, 168, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Sakuta, M.; Hirano, H.; Kakegawa, K.; Suda, J.; Hirose, M.; Joy, R.W., IV; Sugiyama, M.; Komamine, A. Regulatory mechanisms of biosynthesis of betacyanin and anthocyanin in relation to cell division activity in suspension cultures. Plant Cell Tissue Organ Cult. 1994, 38, 167–169. [Google Scholar] [CrossRef]
- Howles, P.A.; Sewalt, V.J.H.; Paiva, N.L.; Elkind, Y.; Bate, N.J.; Lamb, C.; Dixon, R.A. Overexpression of L-phenylalanine ammonia-lyase in transgenic tobacco plants reveals control points for flux into phenylpropanoid biosynthesis. Plant Physiol. 1996, 112, 1617–1624. [Google Scholar] [CrossRef] [Green Version]
- Blount, J.W.; Korth, K.L.; Masoud, S.A.; Rasmussen, S.; Lamb, C.; Dixon, R.A. Altering expression of cinnamic acid 4-hydroxylase in transgenic plants provides evidence for a feedback loop at the entry point into the phenylpropanoid pathway. Plant Physiol. 2000, 122, 107–116. [Google Scholar] [CrossRef]
- Muir, S.R.; Collins, G.J.; Robinson, S.; Hughes, S.; Bovy, A.; de Vos, C.H.R.; van Tunen, A.J.; Verhoeyen, M.E. Overexpression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001, 19, 470–474. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Glucosinolate (μg g−1 Dry Weight) | Oval Chinese Cabbage | Rectangular Chinese Cabbage |
|---|---|---|
| Progoitrin | 298.40 ± 3.89 | 300.07 ± 11.68 |
| Glucoalyssin | 123.5 ± 44.52 *,1 | 94.98 ± 13.55 |
| Gluconapin | 610.21 ± 7.47 *** | 204.56 ± 0.00 |
| 4-hydroxyglucobrassicin | 48.99 ± 9.29 | 109.73 ± 4.64 *** |
| Glucobrassicanapin | 263.35 ± 3.87 *** | 108.38 ± 0.00 |
| Glucobrassicin | 652.00 ± 22.42 | 935.06 ± 76.24 *** |
| 4-methoxyglucobrassicin | 767.00 ± 57.42 | 666.10 ± 43.06 |
| Neoglucobrassicin | 133.29 ± 4.78 | 158.92 ± 9.57 * |
| Total 2 | 2896.78 ± 113.67 * | 2577.79 ± 158.74 |
| Phenolics (μg g−1 Dry Weight) | Oval Chinese Cabbage | Rectangular Chinese Cabbage |
|---|---|---|
| p-hydroxybenzoic acid | 63.41 ± 3.58 **,1 | 47.94 ± 3.03 |
| Catechin hydrate | 11.34 ± 1.06 | n.d 2 |
| Chlorogenic acid | 37.53 ± 0.75 * | 34.86 ± 1.41 |
| Caffeic acid | 5.39 ± 0.39 | 8.00 ± 0.55 *** |
| Epicatechin | 34.24 ± 7.42 | 38.17 ± 0.50 |
| Ferulate | 67.97 ± 1.42 *** | 34.81 ± 1.16 |
| Benzoic acid | n.d | 29.97 ± 1.59 |
| Rutin | 5.11 ± 0.29 | n.d |
| t-cinnamate | 0.99 ± 0.22 | 0.59 ± 0.03 |
| Total 3 | 226.53 ± 15.63 * | 194.34 ± 4.94 |
| Carotenoid (μg g−1 Dry Weight) | Oval Chinese Cabbage | Rectangular Chinese Cabbage |
|---|---|---|
| Violaxanthin | 82.17 ± 1.81 | 134.35 ± 3.19 ***,1 |
| Antheraxanthin | 24.42 ± 0.10 | 43.06 ± 0.13 *** |
| Lutein | 350.06 ± 11.96 | 839.06 ± 9.20 *** |
| Zeaxanthin | 12.34 ± 0.38 | 17.31 ± 0.24 *** |
| β-Cryptoxanthin | 1.72 ± 0.00 | 5.27 ± 0.58 *** |
| 13-cis-β-Carotene | 8.04 ± 0.63 | 19.56 ± 1.77 *** |
| α-Carotene | 2.81 ± 0.07 | 3.91 ± 0.53 * |
| β-Carotene | 251.15 ± 5.65 | 561.94 ± 54.72 *** |
| 9-cis-β-Carotene | 9.04 ± 0.18 | 18.67 ± 2.15 *** |
| Total 2 | 741.73 ± 20.75 | 1,643.13 ± 72.48 *** |
| Chlorophyll (μg g-1 Dry Weight) | Oval Chinese Cabbage | Rectangular Chinese Cabbage |
|---|---|---|
| Chlorophyll a | 434.54 ± 23.91 | 460.47 ± 23.91 |
| Chlorophyll b | 325.42 ± 118.01 | 831.70 ± 110.20 ***,1 |
| Total 2 | 836.48 ± 104.85 | 1332.22 ± 146.51 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.H.; Yeo, H.J.; Park, S.-Y.; Kim, J.K.; Park, S.U. Comparative Phytochemical Analyses and Metabolic Profiling of Different Phenotypes of Chinese Cabbage (Brassica Rapa ssp. Pekinensis). Foods 2019, 8, 587. https://doi.org/10.3390/foods8110587
Park CH, Yeo HJ, Park S-Y, Kim JK, Park SU. Comparative Phytochemical Analyses and Metabolic Profiling of Different Phenotypes of Chinese Cabbage (Brassica Rapa ssp. Pekinensis). Foods. 2019; 8(11):587. https://doi.org/10.3390/foods8110587
Chicago/Turabian StylePark, Chang Ha, Hyeon Ji Yeo, Soo-Yun Park, Jae Kwang Kim, and Sang Un Park. 2019. "Comparative Phytochemical Analyses and Metabolic Profiling of Different Phenotypes of Chinese Cabbage (Brassica Rapa ssp. Pekinensis)" Foods 8, no. 11: 587. https://doi.org/10.3390/foods8110587
APA StylePark, C. H., Yeo, H. J., Park, S.-Y., Kim, J. K., & Park, S. U. (2019). Comparative Phytochemical Analyses and Metabolic Profiling of Different Phenotypes of Chinese Cabbage (Brassica Rapa ssp. Pekinensis). Foods, 8(11), 587. https://doi.org/10.3390/foods8110587