Sprouted Barley Flour as a Nutritious and Functional Ingredient

 ,
, 

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Barley Grains
2.3. Germination
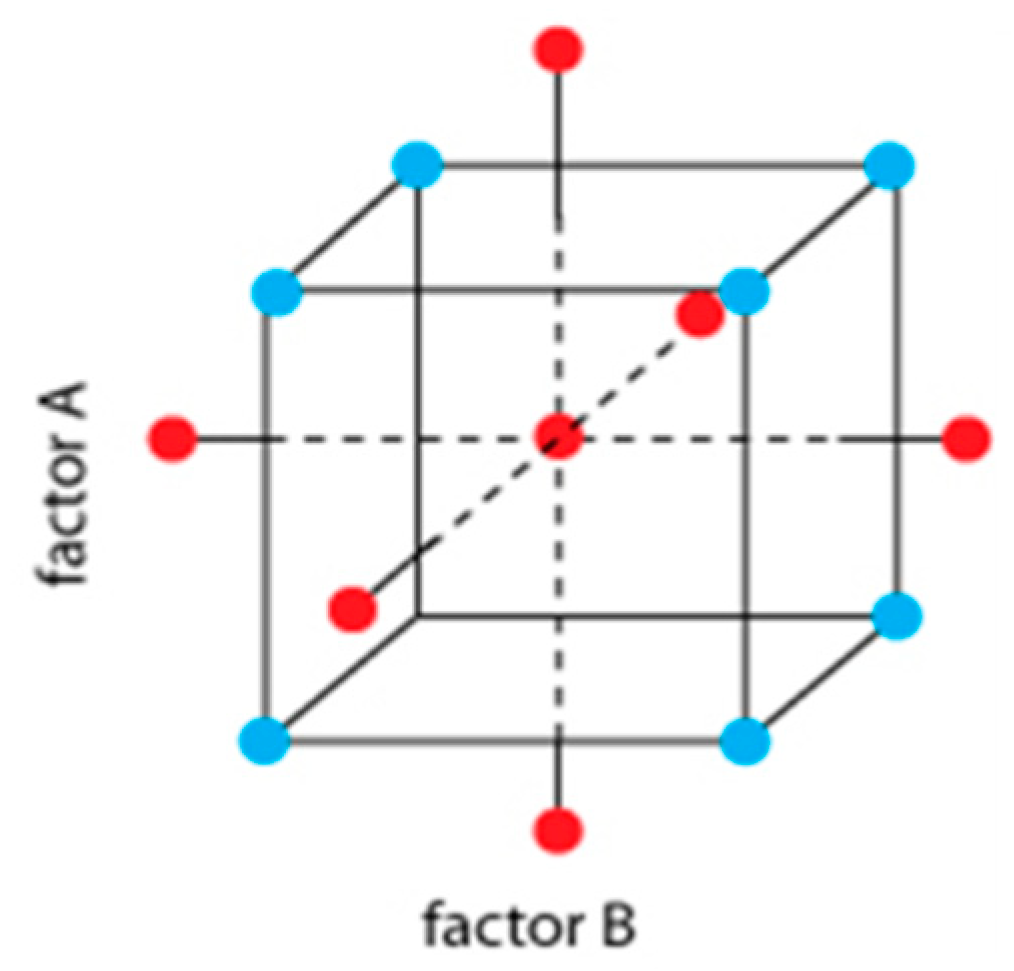
2.4. Experimental Design
2.5. Proximate and Fatty Acid Compositions
2.6. Content of Vitamin C
2.7. Content of Thiamine and Riboflavin
2.8. Content of β-Glucan
2.9. Content of γ-Aminobutyric Acid (GABA)
2.10. Total Phenolic Content Determined by the Folin–Ciocalteu Method
2.11. Content and Profile of Free and Bound Phenolic Compounds
2.12. Total Antioxidant Activity
2.12.1. DPPH Assays (Classical and Q-Versions)
2.12.2. TEAC Assay
2.12.3. ORAC Assay
2.12.4. FRAP Assay
2.13. Glycemic Index
2.14. Colorimetric Characterizationof Germinated Barley Flours
2.15. Sensory Analysis
2.16. Statistical Analysis
3. Results
3.1. Effect of Germination on Proximate Composition of Sprouted Barley Flours
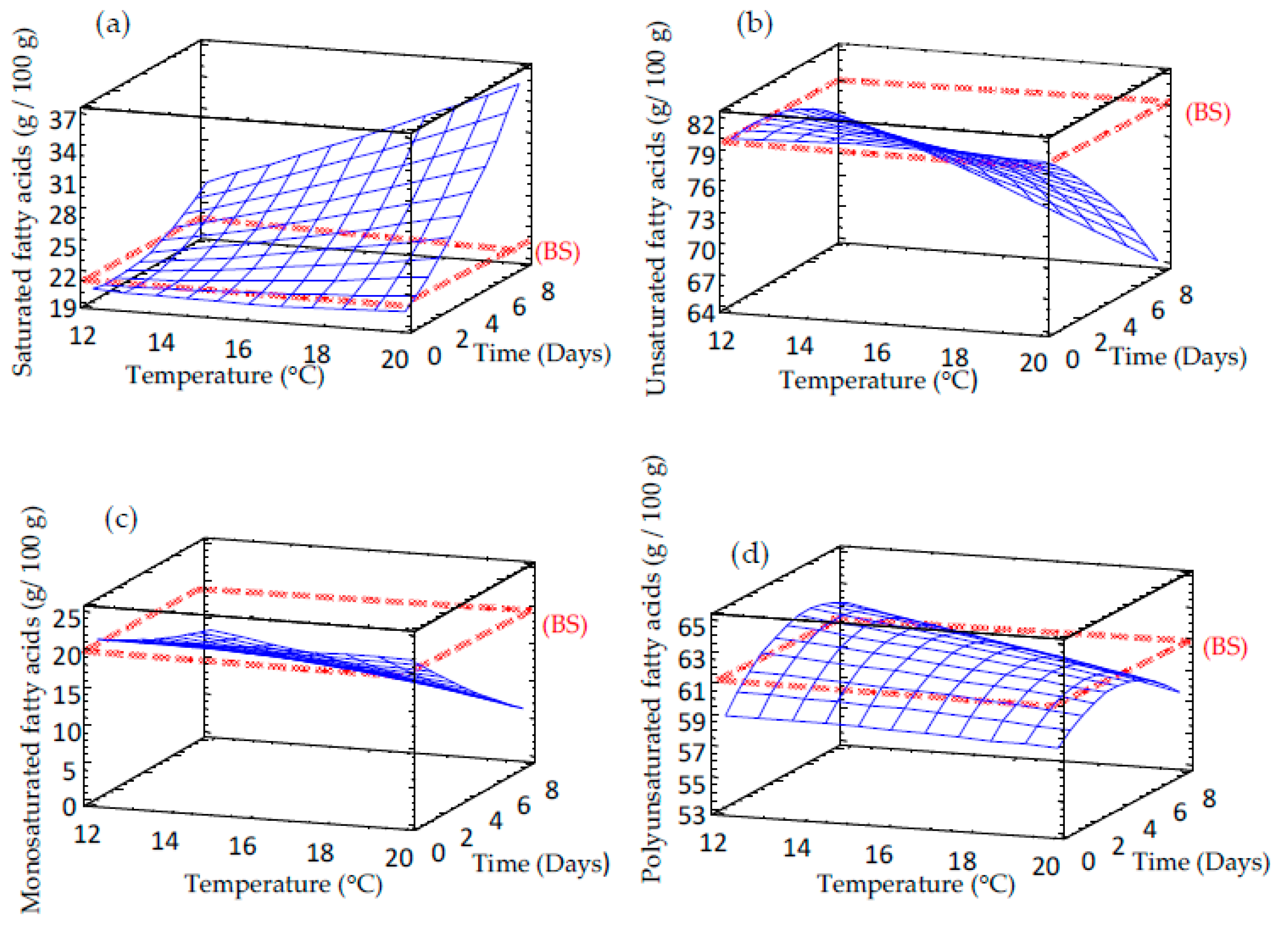
3.2. Effect of Germination on Fatty Acid Content of Sprouted Barley Flours
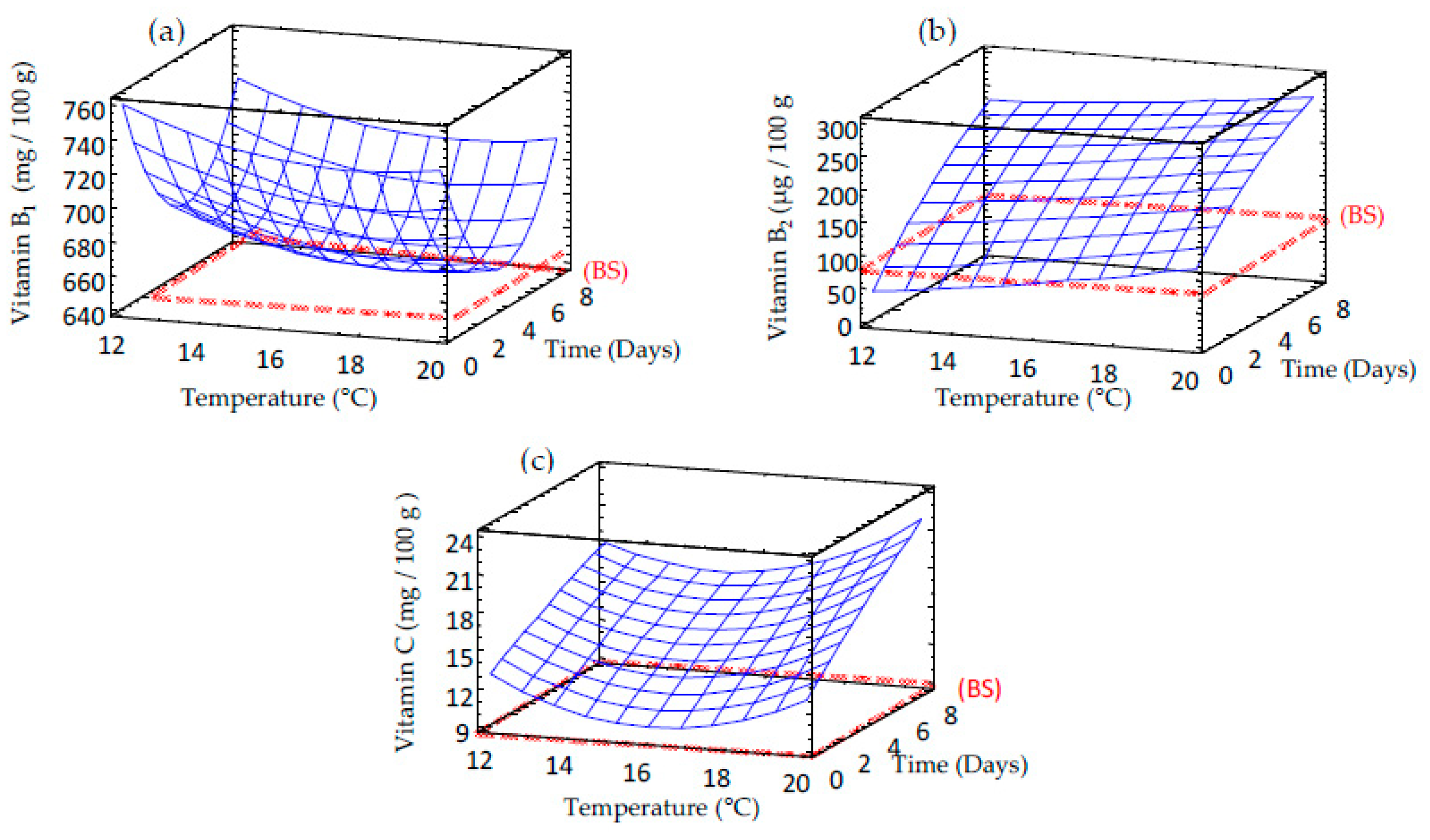
3.3. Effect of Germination on Vitamin Content of Sprouted Barley Flours
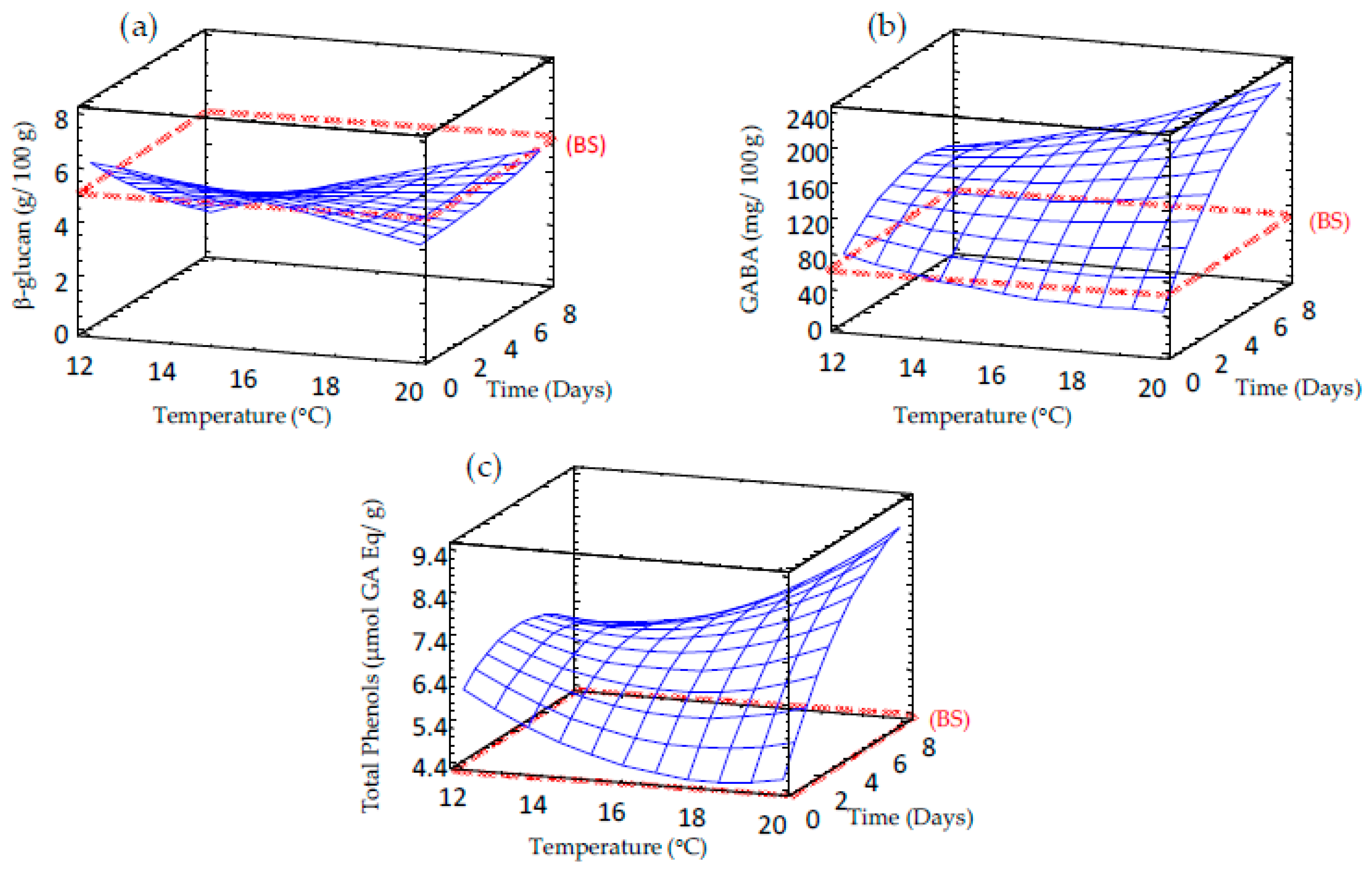
3.4. Effect of Germination on β-Glucan Content of Sprouted Barley Flours
3.5. Effect of Germination on GABA Content of Sprouted Barley Flours
3.6. Effect of Germination on Total Phenolic Content of Sprouted Barley Flours
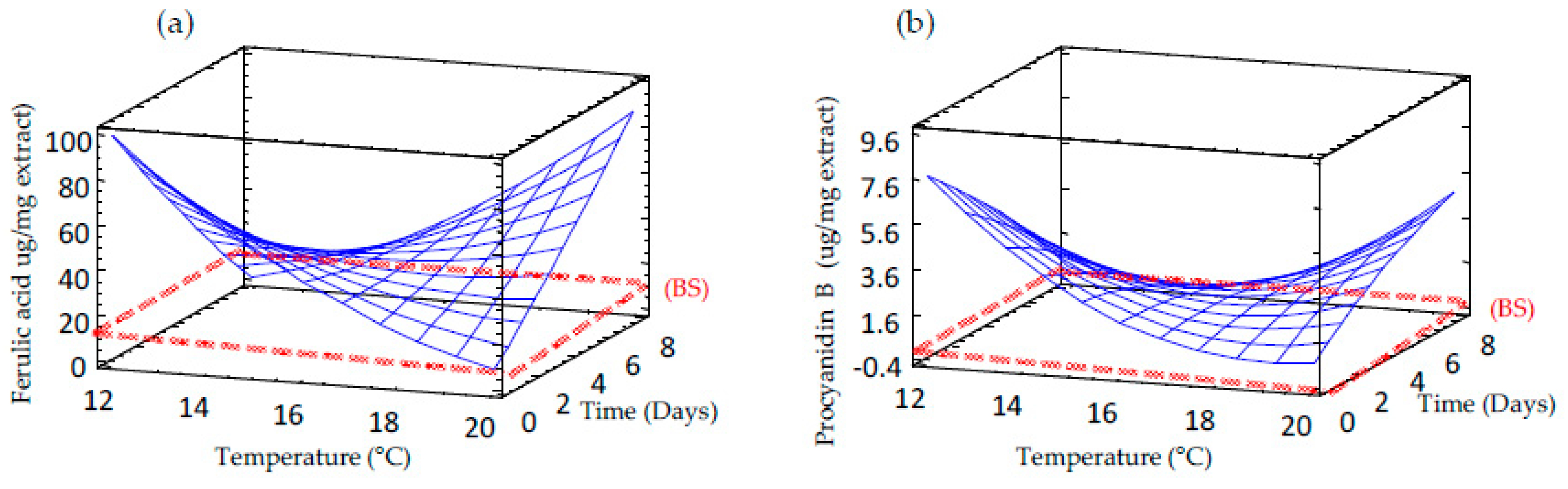
3.7. Effect of Germination on Free and Bound Phenolic Compounds Profile of Sprouted Barley Flours
3.8. Effect of Germination on Antioxidant Activity of Sprouted Barley Flours
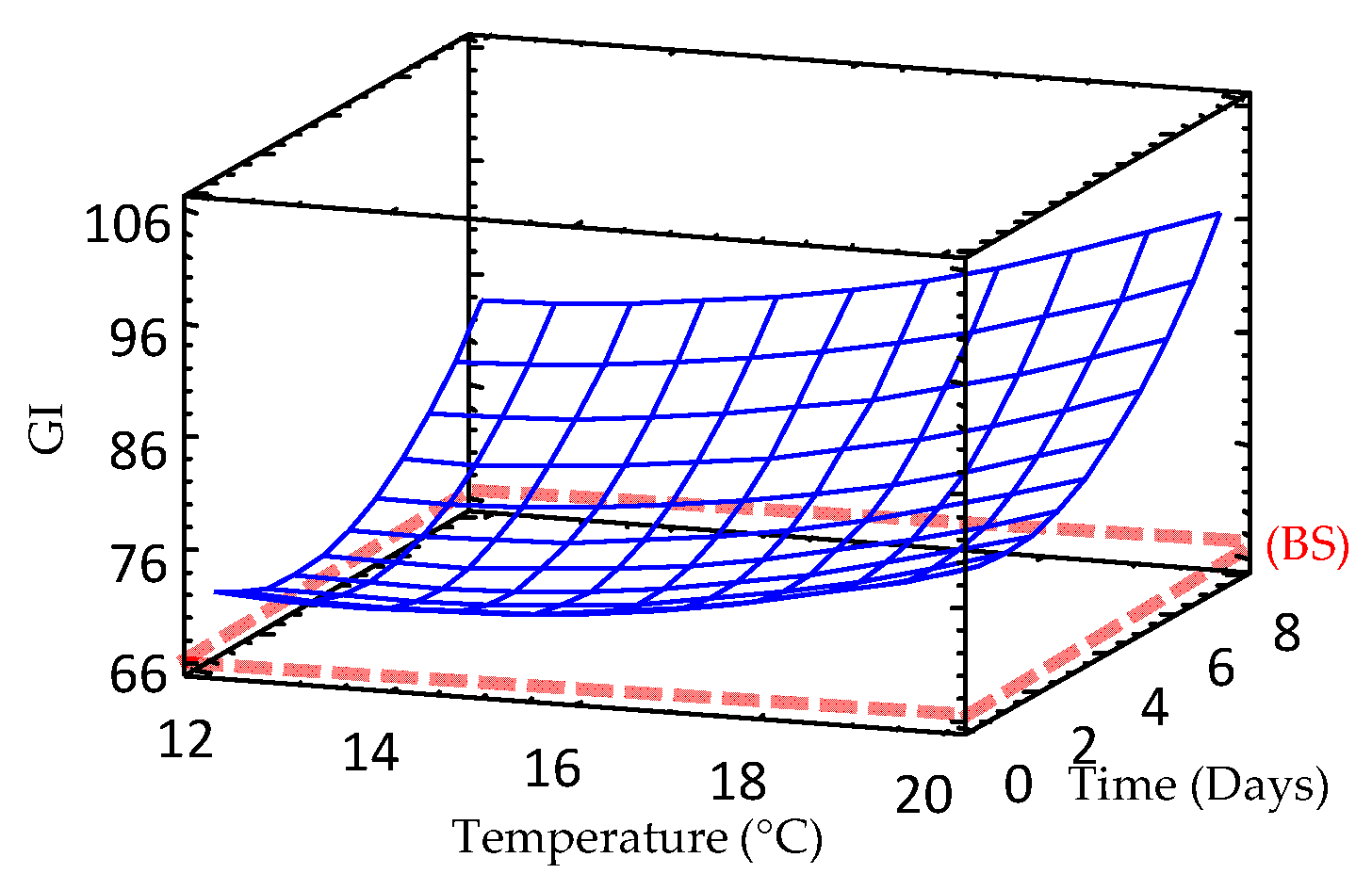
3.9. Effect of Germination on Glycemic Index (GI) of Sprouted Barley Flours
3.10. Effect of Germination on the Colour and Sensory Properties of Sprouted Barley Flours
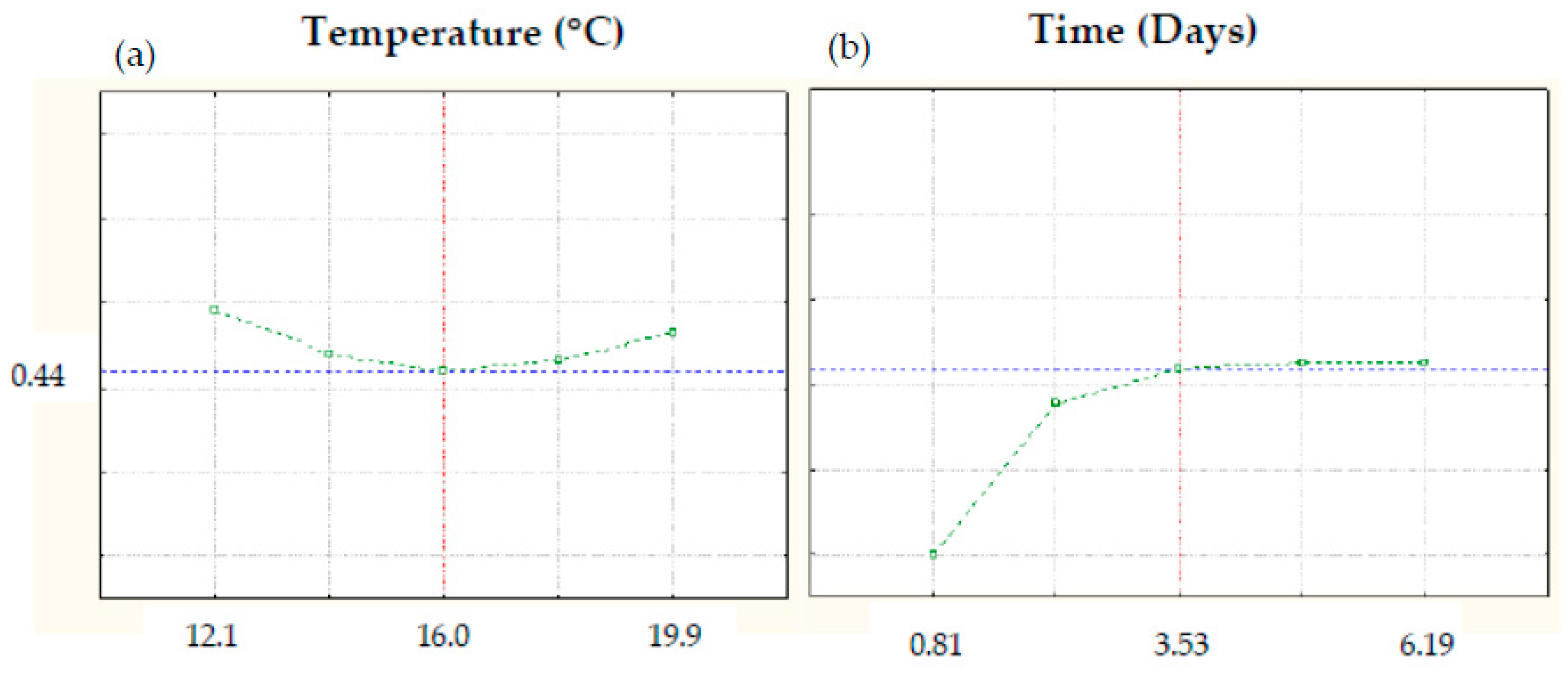
3.11. Multivariate Optimization of Germination Conditions to Obtain High-Quality Germinated Barley Flours
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Madakemohekar, A.; Prasad, L.C.; Pal, J.P.; Prasad, R. Estimation of combining ability and heterosis for yield contributing traits in exotic and indigenous crosses of barley (Hordeum vulgare L.). Res. Crop. 2018, 19, 264–270. [Google Scholar] [CrossRef]
- Martínez, M.; Motilva, M.J.; de las Hazas, M.C.L.; Romero, M.P.; Vaculova, K.; Ludwig, I.A. Phytochemical composition and β-glucan content of barley genotypes from two different geographic origins for human health food production. Food Chem. 2018, 245, 61–70. [Google Scholar] [CrossRef]
- Commission (EU) Regulation No 1048/2012 of 8 November 2012: On the Authorisation of a Health Claim Made on Foods and Referring to the Reduction of Disease Risk. Available online: https://eurex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2012:310:0038:0040:EN:PDF (accessed on 17 February 2020).
- Ötman, E.M.; Frid, A.H.; Groop, L.C.; Bjorck, I.M.E. A dietary exchange of common bread for tailored bread of low glycaemic index and rich in dietary fibre improved insulin economy in young women with impaired glucose tolerance. Eur. J. Clin. Nut. 2005, 60, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Šimić, G.; Horvat, D.; Lalić, A.; Komlenić, D.K.; Abiĉić, I.; Zdunić, Z. Distribution of β-glucan, phenolic acids, and proteins as functional phytonutrients of hull-less barley grain. Foods 2019, 8, 680. [Google Scholar] [CrossRef] [Green Version]
- Madakemohekar, A.; Prasad, L.C.; Prasad, R. Generation mean analysis in barley (Hordeum vulgare L.) under drought stress condition. Plant Arch. 2018, 18, 917–922. [Google Scholar]
- Holtekjølen, A.K.; Olsen, H.R.; Færgestad, E.M.; Uhlen, A.K.; Knutsen, S.H. Variations in water absorption capacity and baking performance of barley varieties with different polysaccharide content and composition. LWT Food Sci. Technol. 2008, 41, 2085–2091. [Google Scholar] [CrossRef]
- Vicentini, A.; Liberatore, L.; Mastrocola, D. Functional foods: Trends and development of the global market. Ital. J. Food. Sci. 2016, 28, 338–351. [Google Scholar]
- Cáceres, P.J.; Martínez-Villaluenga, C.; Amigo, L.; Frias, J. Maximising the phytochemical content and antioxidant activity of Ecuadorian brown rice sprouts through optimal germination conditions. Food Chem. 2014, 152, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Cáceres, P.J.; Peñas, E.; Martinez-Villaluenga, C.; Amigo, L.; Frias, J. Enhancement of biologically active compounds in germinated brown rice and the effect of sun-drying. J. Cereal Sci. 2017, 73, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Frias, J.; Peñas, E.; Vidal-Valverde, C. Changes of vitamin content of powder enteral formulas as a consequence of storage. Food Chem. 2009, 115, 1411–1416. [Google Scholar] [CrossRef]
- Martin-Diana, A.B.; Izquierdo, N.; Albertos, I.; Sanchez, M.S.; Herrero, A.; Sanz, M.A.; Rico, D. Valorization of Carob’s germ and seed peel as natural antioxidant ingredients in gluten-free crackers. J. Food Process. Preserv. 2017, 41, e12770. [Google Scholar] [CrossRef]
- Mattila, P.; Pihlava, J.M.; Hellström, J. Contents of phenolic acids, alkyl- and alkenylresorcinols, and avenanthramides in commercial grain products. J. Agric. Food Chem. 2005, 53, 8290–8295. [Google Scholar] [CrossRef]
- Birsan, R.; Wilde, P.; Waldron, K.; Rai, D. Recovery of polyphenols from brewer’s spent grains. Antioxidants 2019, 8, 380. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Vijayalakshmi, M.; Ruckmani, K. Ferric reducing anti-oxidant power assay in plant extract. Bangladesh J. Pharmacol. 2016, 11, 570–572. [Google Scholar] [CrossRef] [Green Version]
- Gularte, M.; Rosell, C. Physicochemical properties and enzymatic hydrolysis of different starches in the presence of hydrocolloids. Carbohyd. Polym. 2011, 85, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Granfeldt, Y. Foods Factors Affecting Metabolic Responses to Cereal Products. Ph.D. Thesis, University of Lund, Lund, Sweden, 1994. [Google Scholar]
- Onimawo, I.A.; Asugo, S. Effects of germination on the nutrient content and functional properties of pigeon pea flour. J. Food Sci. Technol. 2004, 41, 170–174. [Google Scholar]
- Gómez-Favela, M.A.; Gutiérrez-Dorado, R.; Cuevas-Rodríguez, E.O.; Canizalez-Román, V.A.; del Rosario, C.L.-S.; Milán-Carrillo, J. Improvement of chia seeds with antioxidant activity, GABA, essential amino acids, and dietary fiber by controlled germination bioprocess. Plant Food Hum. Nutr. 2017, 72, 345–352. [Google Scholar] [CrossRef]
- Chavarín-Martínez, C.D.; Gutiérrez-Dorado, R.; Perales-Sánchez, J.X.K.; Cuevas-Rodríguez, E.O.; Milán-Carrillo, J.; Reyes-Moreno, C. Germination in optimal conditions as effective strategy to improve nutritional and nutraceutical value of underutilized Mexican blue maize seeds. Plant Food Hum. Nutr. 2019, 74, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Krist, S.; Stuebiger, G.; Unterweger, H.; Bandion, F.; Buchbauer, G. Analysis of volatile compounds and triglycerides of seed oils extracted from different poppy varieties (Papaver somniferum L.). J. Agric. Food Chem. 2005, 53, 8310–8316. [Google Scholar] [CrossRef] [PubMed]
- Copeland, L.; McDonald, M.B. Seed germination. In Principle of Seed Science and Technology, 4th ed.; Copeland, L., Ed.; Springer: Cham, Switzerland, 2001; pp. 72–124. [Google Scholar]
- Bibi, N.; Aurang, Z.; Amal, B.K.; Mohammad, S.K. Effect of germination time and type of illumination on proximate composition of chickpea seed (Cicer arietinum L.). Am. J. Food Technol. 2008, 3, 24–32. [Google Scholar] [CrossRef]
- Arora, S.; Jood, S.; Khetarpaul, N. Effect of germination and probiotic fermentation on nutrient composition of barley based food mixtures. Food Chem. 2010, 119, 779–784. [Google Scholar] [CrossRef]
- Marconi, O.; Tomasi, I.; Dionisio, L.; Perretti, G.; Fantozzi, P. Effects of malting on molecular weight distribution and content of water-extractable β-glucans in barley. Food Res. Int. 2014, 64, 677–682. [Google Scholar] [CrossRef]
- Betts, N.S.; Berkowitz, O.; Liu, R.; Collins, H.M.; Skadhauge, B.; Dockter, C.; Burton, R.A.; Whelan, J.; Fincher, G.B. Isolation of tissues and preservation of RNA from intact, germinated barley grain. Plant J. 2017, 91, 754–765. [Google Scholar] [CrossRef] [Green Version]
- Rimsten, L.; Haraldsson, A.-K.; Andersson, R.; Alminger, M.; Sandberg, A.-S.; Åman, P. Effects of malting on glucanase and phytase activity in barley grain. J. Sci. Food Agric. 2002, 82, 904–912. [Google Scholar] [CrossRef]
- Farzaneh, V.; Ghodsvali, A.; Bakhshabadi, H.; Zare, Z.; Carvalho, I.S. The impact of germination time on the some selected parameters through malting process. Int. J. Biol. Macromol. 2017, 94, 663–668. [Google Scholar] [CrossRef]
- Chu, C.; Yan, N.; Du, Y.; Liu, X.; Chu, M.; Shi, J.; Zhang, H.; Liu, Y.; Zhang, Z. iTRAQ-based proteomic analysis reveals the accumulation of bioactive compounds in Chinese wild rice (Zizania latifolia) during germination. Food Chem. 2019, 289, 635–644. [Google Scholar] [CrossRef]
- Yang, R.; Chen, H.; Gu, Z. Factors influencing diamine oxidase activity and γ-aminobutyric acid content of fava bean (Vicia faba L.) during germination. J. Agric. Food Chem. 2011, 59, 11616–11620. [Google Scholar] [CrossRef]
- Chung, H.J.; Jang, S.H.; Cho, H.Y.; Lim, S.T. Effects of steeping and anaerobic treatment on GABA (γ-aminobutyric acid) content in germinated waxy hull-less barley. LWT Food Sci. Technol. 2009, 42, 1712–1716. [Google Scholar] [CrossRef]
- Zhao, H.; Fan, W.; Dong, J.; Lu, J.; Chen, J.; Shan, L.; Lin, Y.; Kong, W. Evaluation of antioxidant activities and total phenolic contents of typical malting barley varieties. Food Chem. 2008, 107, 296–304. [Google Scholar] [CrossRef]
- Leitao, C.; Marchioni, E.; Bergaentzlé, M.; Zhao, M.; Didierjean, L.; Miesch, L.; Holder, E.; Miesch, M.; Ennahar, S. Fate of polyphenols and antioxidant activity of barley throughout malting and brewing. J. Cereal Sci. 2012, 55, 318–322. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Gonçalves, L.M.; Guido, L.F. Overall antioxidant properties of malt and how they are influenced by the individual constituents of barley and the malting process. Compr. Rev. Food Sci. Food Saf. 2016, 15, 927–943. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Curto, A.F.; Guido, L.F. Determination of phenolic content in different barley varieties and corresponding malts by liquid chromatography-diode array detection-electrospray ionization tandem mass spectrometry. Antioxidants 2015, 4, 563–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangopadhyay, N.; Rai, D.K.; Brunton, N.P.; Gallagher, E.; Hossain, M.B. Antioxidant-guided isolation and mass spectrometric identification of the major polyphenols in barley (hordeum vulgare) grain. Food Chem. 2016, 210, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Zilic, S.; Basic, Z.; Sucakovic, V.H.T.; Maksimovic, V.; Jancovic, M.; Filipovic, M. Can the sprouting process applied to wheat improve the contents of vitamins and phenolic compounds and antioxidant capacity of the flour? Int. J. Food Sci. Technol. 2014, 49, 1040–1047. [Google Scholar] [CrossRef]
- Yang, F.; Basu, T.K.; Ooraikul, B. Studies on germination conditions and antioxidant contents of wheat grain. Int. J. Food Sci. Nutr. 2001, 52, 319–330. [Google Scholar] [CrossRef]
- Kim, M.J.; Kwak, H.C.S.; Kim, S.S. Effects of germination on protein, γ-aminobutyric acid, phenolic acids, and antioxidant capacity in wheat. Molecules 2018, 23, 2244. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Orozco, R.; Frias, J.; Zielinski, H.; Piskula, M.K.; Kozlowska, H.; Vidal-Valverde, C. Kinetic study of the antioxidant compounds and antioxidant capacity during germination of Vigna radiata cv. emmerald, Glycine max cv. jutro and Glycine max cv. merit. Food Chem. 2008, 111, 622–630. [Google Scholar] [CrossRef]
- Gordillo-Bastidas, E.; Díaz-Rizzolo, D.A.; Roura, E.; Massanés, T.; Gomis, R. Quinoa (Chenopodium quinoa wild), from nutritional value to potential health benefits: An integrative review. J. Nutr. Food Sci. 2016, 6, 1–10. [Google Scholar]
- Charoenthaikij, P.; Jangchud, K.; Jangchud, A.; Prinyawiwatkul, W.; No, H. Composite wheat–germinated brown rice flours: Selected physicochemical properties and bread application. Int. J. Food Sci. Technol. 2012, 47, 75–82. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor A Temperature (°C) | Factor B Time (Days) | Experiments (Codes) |
|---|---|---|
| 12.1 | 3.58 | 1 |
| 13.2 | 1.60 | 2 |
| 13.2 | 5.52 | 3 |
| 16.0 | 0.81 | 4 |
| 16.0 | 3.48 | 5* |
| 16.0 | 6.19 | 6 |
| 18.8 | 1.60 | 7 |
| 18.8 | 5.52 | 8 |
| 19.9 | 3.60 | 9 |
| Response Variables | Model Equations | Optimum Temperature (Tp) | Optimum Time (Tm) |
|---|---|---|---|
| Fat | 4.984 – 0.214 × TP + 0.022 × TM + 0.006 × TP2 – 0.008 × TM × TP – 0.009 × TP2 | 12.1 | 0.81 |
| Protein | 19.066 – 0.551 × TM + 0.167 × TP + 0.019 × TM2 + 0.010 × TM × TP – 0.026 × TP2 | 19.9 | 6.19 |
| Fiber | 35.541 – 2.362 × TP – 1.176 × TM + 0.080 × TP2 + 0.00004 × TM × TP + 0.098 × TP2 | 19.9 | 0.81 |
| Carbohydrates | 65.948 – 0.075 × TM + 2.804 × TP + 0.025 × TM2 – 0.098 × TM × TP – 0.121 × TP2 | 12.1 | 0.81 |
| Saturated fatty acids | 24.444 – 0.587 × TM – 2.426 × TP + 0.019 × TM2 + 0.179 × TP × TM + 0.010 × TP2 | 19.9 | 6.19 |
| Unsaturated fatty acids | 75.556 + 0.587 × TM + 2.426 × TP – 0.019 × TM2 – 0.179 × TM × TP – 0.100 × TP2 | 12.1 | 0.81 |
| Monounsat. fatty acids | 17.847 + 0.398 × TM + 0.288 × TP – 0.010 × TP2 – 0.123 × TM × TP + 0.036 × TP2 | 14.3 | 0.81 |
| Polyunsat. fatty acids | 57.747 + 0.184 × TM + 2.141 × TP – 0.008 × TP2 – 0.056 × TM × TP – 0.136 × TP2 | 12.1 | 5.4 |
| Vitamin C | 61.315 – 6.418 × TP + 0.101 × TM + 0.110 × TP2 + 0.060 × TM × TP – 0.020 × TP2 | 19.9 | 6.19 |
| Vitamin B1 | 1003.83 – 30.743×TM – 37.626×TP + 0.867 × TM2 + 0.059 × TP × TM + 4.207 × TM2 | 12.1 | 0.81 |
| Vitamin B2 | 4.880 – 0.329 × TP + 39.175 × TM + 0.290 × TP2 – 0.460 × TM × TP – 1.271 × TP2 | 19.9 | 6.19 |
| β-glucan | 6.886 + 0.052 × TP – 1.68 × TM – 0.009 × TP2 + 0.082 × TP × TM + 0.014 × TP2 | 12.1 | 0.81 |
| GABA | 311.39 – 28.559 × TM – 4.590 × TP + 0.759 × TM2 + 2.101 × TP × TM – 1.924 × TM2 | 19.9 | 6.19 |
| TPC | 367.582 – 31.560 × TP – 8.571 × TM + 0.874 × TM2 + 1.297×TP × TM – 0.929 × TP2 | 19.9 | 6.19 |
| Bound ferulic acid | 423.975 – 36.487 × TP – 47.023 × TM + 0.784 × TP2 + 2.769 × TM × TP + 0.212 × TP2 | 12.1 | 0.81 |
| Free procyanidin | 53.593 – 5.589 × TP – 2.911 × TM + 0.147 × TP2 + 0.181 × TM × TP – 0.025 × TP2 | 12.1 | 0.81 |
| FRAP | 0.407 – 0.027 × TP – 0.017 × TM + 0.0007 × TP2 + 0.0009 × TM × TP + 0.0005 × TP2 | 19.9 | 6.19 |
| ORAC | 205.967 – 23.180 × TM – 0.708 × TP + 0.709 × TM2 + 0.923 × TM × TP – 1.361 × TP2 | 19.9 | 6.19 |
| DPPH | 4274.09 – 291.532 × TP – 284.032 × TM + 5.985 × TP2 + 18.461 × TM × TP – 0.231 × TP2 | 12.1 | 0.81 |
| TEAC | 15296.3 – 1782.21 × TM + 1758.61 × TP + 59.311 × TM2 – 21.386 × TP × TM − 119.929 × TP2 | 19.9 | 6.19 |
| DPPH-Q * | 5252.4 – 507.735 × TP – 136.6 × TM + 14.287 × TP2 + 11.670 × TM × TP – 0.503 × TM2 | 19.9 | 6.19 |
| L* | 100.793 – 1.859 × TM + 0.478 × TP + 0.054 × TM2 – 0.003 × TM × TP – 0.069 × TP2 | 12.1 | 3.2 |
| a* | 0.455 + 0.088 × TM + 0.182 × TP − 0.002 × TM2 – 0.013 × TM × TP – 0.0004 × TP2 | 19.9 | 0.81 |
| b* | 16.906 – 0.874 × TM – 0.564 × TP + 0.025 × TM2 + 0.057 × TP × TM – 0.014 × TP2 | 12.1 | 0.81 |
| Glycemic index | 105.067 – 4.918 × TM – 3.373 × TP + 0.180 × TM2 + 0.093 × TP × TM + 0.471 × TP2 | 19.9 | 6.19 |
| Color | (−) 0.322 + 0.0469 × TM + 0.075 × TP | 12.1 | 0.81 |
| Texture | 0.268 + 0.039 × TM − 0.011 × TP | 12.1 | 0.81 |
| Flavour | (−) 0.365 + 0.047 × TM + 0.085 × TP | 19.9 | 6.19 |
| Fraction | Germination Experiments | Catechin | Protocatechuic Acid | Diferulic Acid | 4-hydroxybenzoic Acid | Phloridzin | Ferulic Acid | Caffeic Acid | 4-coumaric Acid | Procyanidin B |
|---|---|---|---|---|---|---|---|---|---|---|
| Free | BS | 0.99 ± 0.16 cB | 0.01 ± 0.02 aA | 2.22 ± 0.31 dC | ND aA | 0.01 ± 0.00 aA | ND aA | ND aA | ND aA | 12.44 ± 0.80 cD |
| 1 | 0.20 ± 0.04 bB | 0.02 ± 0.00 bA | 0.76 ± 0.32 cC | ND aA | 0.02 ± 0.01 aA | ND aA | ND aA | ND aA | 1.94 ± 0.56 abB | |
| 2 | 0.02 ± 0.00 aA | 0.02 ± 0.00 bA | 0.36 ± 0.19 bB | ND aA | 0.02 ± 0.01 aA | ND aA | ND aA | ND aA | 3.26 ± 1.41 bC | |
| 3 | 0.53 ± 0.40 bcB | 0.01 ± 0.02 aA | 1.19 ± 0.02 eB | ND aA | 0.03 ± 0.01 aA | ND aA | ND aA | ND aA | 1.40 ± 0.57 abB | |
| 4 | 0.12 ± 0.12 aA | 0.03 ± 0.00 bA | 1.06 ± 0.07 deB | ND aA | 0.01 ± 0.00 aA | ND aA | ND aA | ND aA | 1.06 ± 0.01 aB | |
| 5 | 0.18 ± 0.02 bB | 0.02 ± 0.00 bA | 1.61 ± 0.84 defC | ND aA | 0.02 ± 0.01 aA | ND aA | ND aA | ND aA | 1.68 ± 1.11 abC | |
| 6 | 0.67 ± 0.71 bcAB | 0.03 ± 0.00 bA | 1.90 ± 0.82 efB | ND aA | 0.01 ± 0.00 aA | ND aA | ND aA | ND aA | 2.79 ± 2.23 aB | |
| 7 | 0.08 ± 0.08 aA | 0.03 ± 0.00 bA | 0.65 ± 0.00 cB | ND aA | 0.01 ± 0.00 aA | ND aA | ND aA | ND aA | 0.49 ± 0.70 aAB | |
| 8 | 0.87 ± 0.75 bcB | 0.02 ± 0.00 bA | 1.03 ± 0.10 cdB | ND aA | 0.01 ± 0.00 aA | ND aA | ND aA | ND aA | 2.68 ± 2.05 abB | |
| 9 | 0.57 ± 0.36 bcB | 0.03 ± 0.00 bA | 0.99 ± 0.02 dB | ND aA | 0.02 ± 0.00 aA | ND aA | ND aA | ND aA | 6.72 ± 2.87 bC | |
| Bound | BS | ND aA | 0.07 ± 0.05 bcB | 0.26 ± 0.05 bD | 0.45 ± 0.01 dE | 0.02 ± 0.01 aA | 45.06 ± 3.09 dH | 0.16 ± 0.01 bC | 2.34 ± 0.40 cdG | 1.07 ± 0.14 aF |
| 1 | ND aA | 0.10 ± 0.00 cB | 0.09 ± 0.00 aB | 0.31 ± 0.02 cC | 0.01 ± 0.00 aA | 36.45 ± 2.44 cF | 0.70 ± 0.09 dC | 2.46 ± 0.30 cdE | 1.22 ± 0.18 aD | |
| 2 | 0.02 ± 0.03 aA | 0.03 ± 0.01 abA | 0.09 ± 0.00 aB | 0.21 ± 0.04 bC | 0.01 ± 0.00 aA | 50.27 ± 9.18 deF | 0.13 ± 0.01 bB | 2.32 ± 0.43 cdE | 1.07 ± 0.33 aD | |
| 3 | ND aA | 0.08 ± 0.06 bcB | 0.19 ± 0.15 abBC | 0.22 ± 0.02 bC | 0.02 ± 0.01 aA | 31.15 ± 1.89 cE | 0.33 ± 0.19 bcC | 1.56 ± 0.27 bcD | 1.23 ± 0.29 aD | |
| 4 | ND aA | 0.05 ± 0.00 bB | 0.19 ± 0.02 bC | 0.27 ± 0.09 bcC | 0.02 ± 0.01 aA | 31.25 ± 5.16 cF | 0.22 ± 0.01 cC | 1.92 ± 0.19 cd.E | 0.90 ± 0.03 aD | |
| 5 | ND aA | 0.06 ± 0.01 bB | 1.18 ± 0.1 eD | 0.15 ± 0.08 bB | 0.01 ± 0.01 aA | 23.82 ± 1.02 bF | 0.67 ± 0.01 dC | 1.40 ± 0.03 bE | 0.66 ± 0.94 aBCD | |
| 6 | ND aA | 0.08 ± 0.02 bc.B | 1.52 ± 0.70 defCD | 0.22 ± 0.18 bcB | 0.01 ± 0.00 aA | 28.40 ± 17.43 bcdE | 0.68 ± 0.41 cdC | 2.01 ± 1.06 bcdD | 0.89 ± 0.06 aC | |
| 7 | ND aA | 0.04 ± 0.03 abA | 1.03 ± 0.11 dC | 0.12 ± 0.07 bB | 0.01 ± 0.01 aA | 31.27 ± 9.38 cdE | 0.26 ± 0.10 cB | 1.79 ± 0.57 bcD | 1.08 ± 0.14 aC | |
| 8 | ND aA | 0.15 ± 0.07 cB | 1.21 ± 1.12 bcdeCD | 0.41 ± 0.01 dC | 0.02 ± 0.01 aA | 72.95 ± 12.73 eF | 1.21 ± 0.21 eD | 2.60 ± 0.51 cdE | 0.99 ± 0.07 aD | |
| 9 | ND aA | 0.13 ± 0.03 c | 2.88 ± 0.42 fE | 0.32 ± 0.09 cC | 0.01 ± 0.01 aA | 43.89 ± 6.59 dF | 0.98 ± 0.05 eD | 3.28 ± 0.85 dE | 1.01 ± 0.07 aD |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rico, D.; Peñas, E.; García, M.d.C.; Martínez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martín-Diana, A.B. Sprouted Barley Flour as a Nutritious and Functional Ingredient. Foods 2020, 9, 296. https://doi.org/10.3390/foods9030296
Rico D, Peñas E, García MdC, Martínez-Villaluenga C, Rai DK, Birsan RI, Frias J, Martín-Diana AB. Sprouted Barley Flour as a Nutritious and Functional Ingredient. Foods. 2020; 9(3):296. https://doi.org/10.3390/foods9030296
Chicago/Turabian StyleRico, Daniel, Elena Peñas, María del Carmen García, Cristina Martínez-Villaluenga, Dilip K. Rai, Rares I. Birsan, Juana Frias, and Ana B. Martín-Diana. 2020. "Sprouted Barley Flour as a Nutritious and Functional Ingredient" Foods 9, no. 3: 296. https://doi.org/10.3390/foods9030296
APA StyleRico, D., Peñas, E., García, M. d. C., Martínez-Villaluenga, C., Rai, D. K., Birsan, R. I., Frias, J., & Martín-Diana, A. B. (2020). Sprouted Barley Flour as a Nutritious and Functional Ingredient. Foods, 9(3), 296. https://doi.org/10.3390/foods9030296