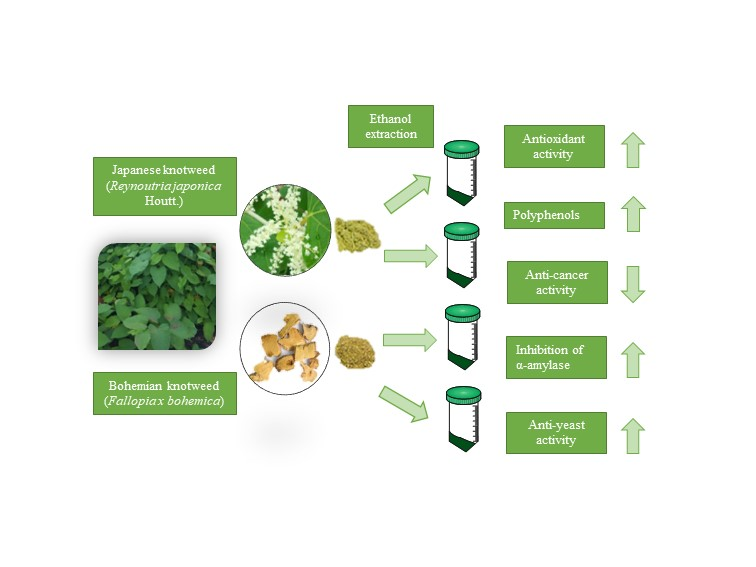
In Vitro Comparison of the Bioactivities of Japanese and Bohemian Knotweed Ethanol Extracts

 ,
, 
 ,
, 

Abstract
:
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Plant Material and Extraction Procedure
2.3. Identification and Quantification of Phenolic Compounds with HPLC/DAD
2.4. Antioxidant Capacity by DPPH
2.5. Cellular Antioxidant Activity Assay
2.6. Cytotoxicity Assay
2.7. Antidiabetic Activity Assay
2.8. Antimicrobial Activity
2.9. Data Procesing and Statistical Analysis
3. Results and Discussion
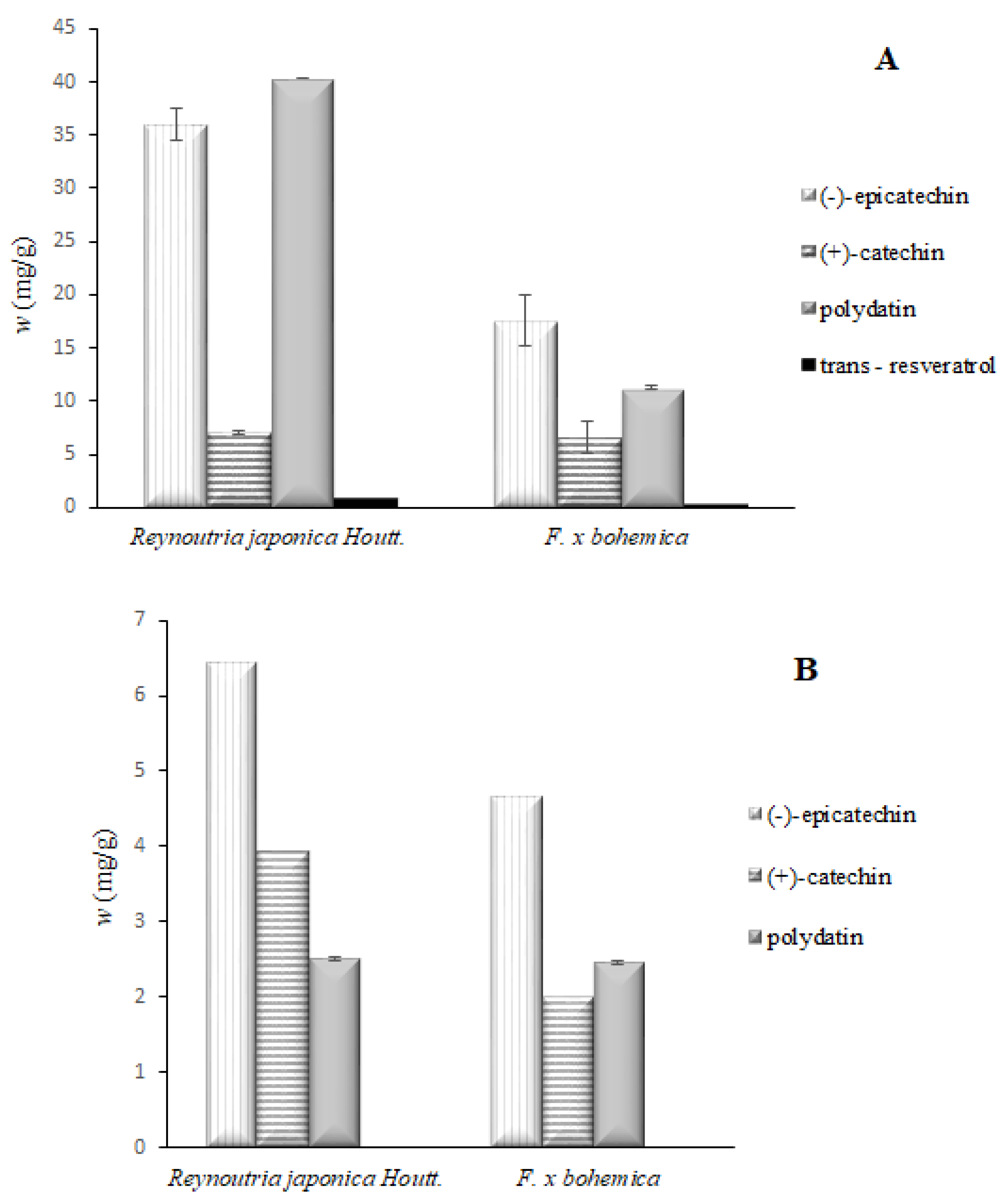
3.1. Identification and Quantification of Phenolic Compounds
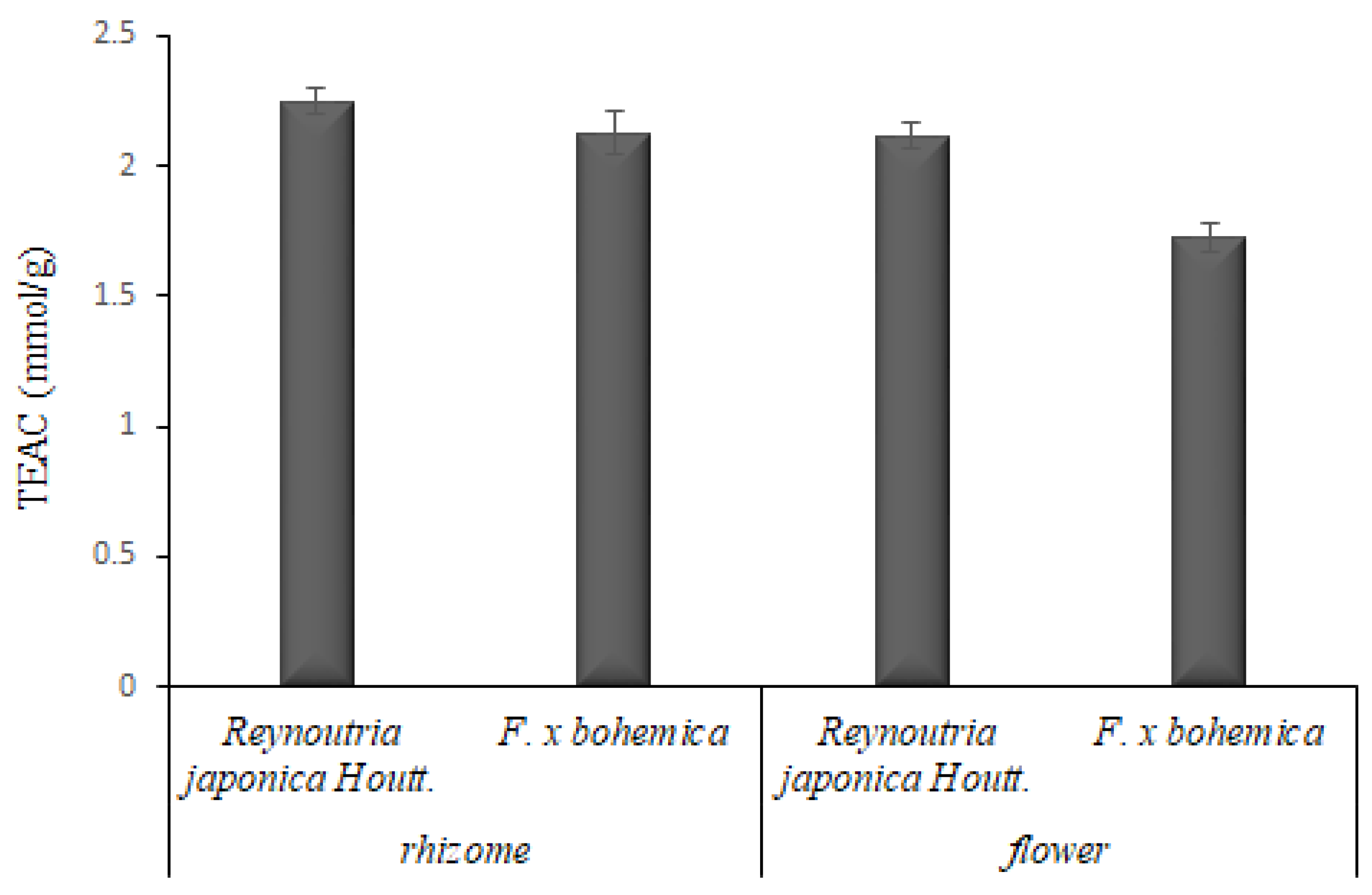
3.2. Antioxidant Capacity
3.2.1. DPPH Scavenging Capacity Assay
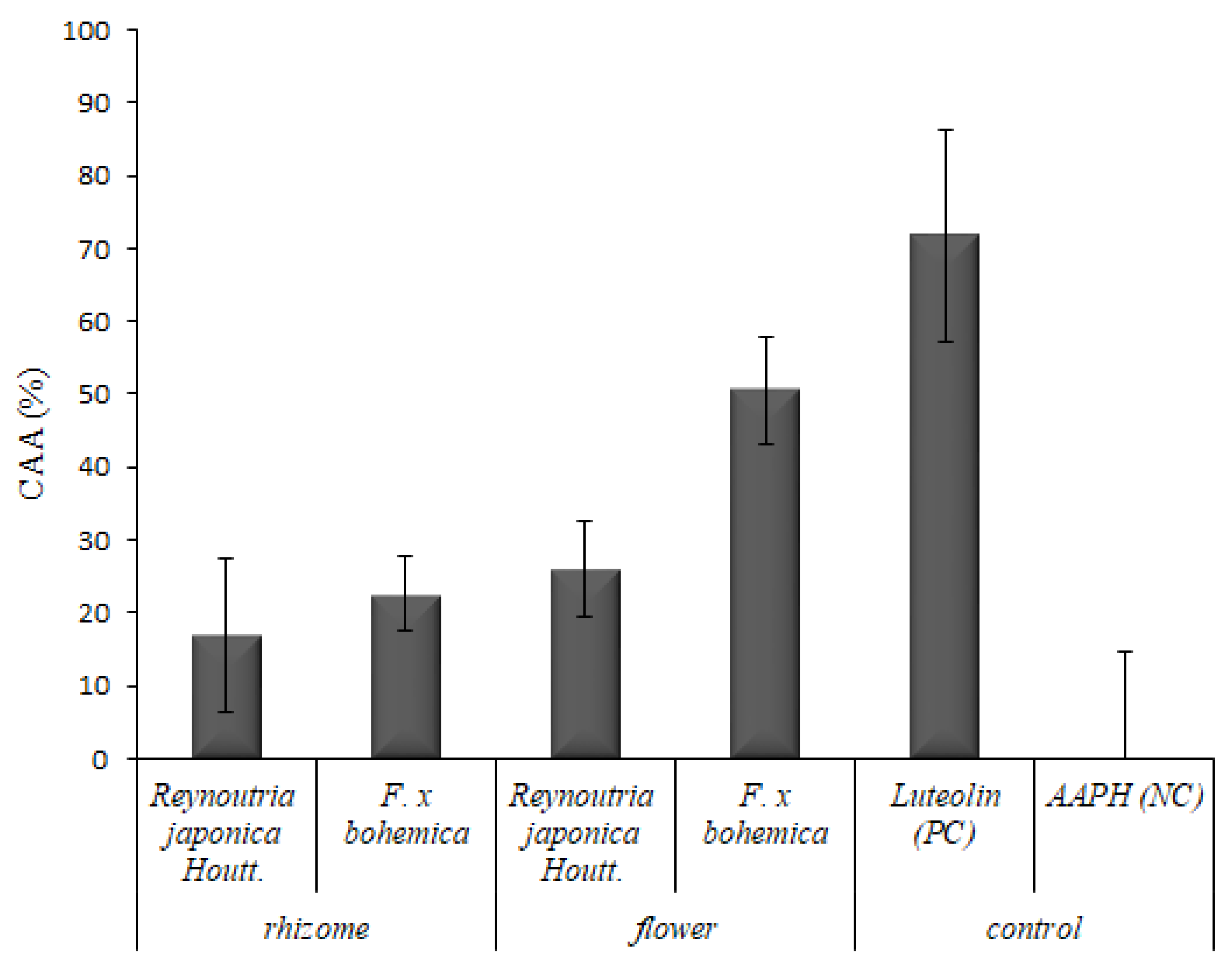
3.2.2. Cellular Antioxidant Activity (CAA) Assay
3.3. Cytotoxicity Activity
3.4. Antidiabetic Activity
3.5. Antimicrobial Activity
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bailey, J.P.; Conolly, A. Prize-winners to pariahs—A history of Japanese knotweed s.l. (Polygonaceae) in the British Isles. Watsonia 2000, 23, 93–110. [Google Scholar]
- Murrell, C.; Gerber, E.; Krebs, C.; Parepa, M.; Schaffner, U.; Bossdorf, O. Invasive Knotweed Affects Native Plants through Allelopathy. Am. J. Bot. 2011, 98, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Strgulc Krajšek, S.; Jogan, N. The genus Fallopia Adans. in Slovenia. Hladnikia 2011, 28, 17–40. [Google Scholar]
- Bailey, J.P.; Bimova, K.; Mandak, B. Asexual spread versus sexual reproduction and evolution in Japanese Knotweed s.l. sets the stage for the “Battle of the Clones”. Biol. Invasions 2009, 11, 1189–1203. [Google Scholar] [CrossRef]
- Lachowicz, S.; Oszmiański, J. Profile of Bioactive Compounds in the Morphological Parts of Wild Fallopia japonica (Houtt) and Fallopia sachalinensis (F. Schmidt) and Their Antioxidative Activity. Molecules 2019, 24, 1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, T.; Zhang, H.; Cai, Z. Analysis of rhizoma polygoni cuspidati by HPLC and HPLC-ESI/MS. Phytochem. Anal. 2007, 18, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.G.; Liu, W.Y.; Chen, G.T. A simple method for the isolation and purification of resveratrol from Polygonum cuspidatum. J. Pharm. Anal. 2013, 3, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.G.; Tuck, T.; Ji, X.H.; Zhou, X.; Kelly, G.; Cuerrier, A.; Zhang, J.Z. Quality Assessment of Japanese Knotweed (Fallopia japonica) Grown on Prince Edward Island as a Source of Resveratrol. J. Agric. Food Chem. 2013, 61, 6383–6392. [Google Scholar] [CrossRef]
- Kovarova, M.; Bartunkova, K.; Frantik, T.; Koblihova, H.; Prchlova, K.; Vosatka, M. Factors influencing the production of stilbenes by the knotweed, Reynoutria × bohemica. BMC Plant Biol. 2010, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Vrchotova, N.; Sera, B.; Dadakova, E. HPLC and CE analysis of catechins, stilbens and quercetin in flowers and stems of Polygonum Cuspidatum, P. sachalinense and P. x bohemicum. J. Indian Chem. Soc. 2010, 87, 1267–1272. [Google Scholar]
- Metlicar, V.; Vovk, I.; Albreht, A. Japanese and Bohemian Knotweeds as Sustainable Sources of Carotenoids. Plants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.; Okuda, H. Resveratrol isolated from Polygonum cuspidatum root prevents tumor growth and metastasis to lung and tumor-induced neovascularization in Lewis lung carcinoma-bearing mice. J. Nutr. 2001, 131, 1844–1849. [Google Scholar] [CrossRef]
- Lee, C.C.; Chen, Y.T.; Chiu, C.C.; Liao, W.T.; Liu, Y.C.; Wang, H.M.D. Polygonum cuspidatum extracts as bioactive antioxidaion, anti-tyrosinase, immune stimulation and anticancer agents. J. Biosci. Bioeng. 2015, 119, 464–469. [Google Scholar] [CrossRef]
- Shan, B.; Cai, Y.Z.; Brooks, J.D.; Corke, H. Antibacterial properties of Polygonum cuspidatum roots and their major bioactive constituents. Food Chem. 2008, 109, 530–537. [Google Scholar] [CrossRef]
- Kumagai, H.; Kawai, Y.; Sawano, R.; Kurihara, H.; Yamazaki, K.; Inoue, N. Antimicrobial substances from rhizomes of the giant knotweed Polygonum sachalinense against the fish pathogen Photobacterium damselae subsp piscicida. Z. Naturforsch. C 2005, 60, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.Y.; Wu, C.H.; Qiu, S.H.; Yuan, X.L.; Li, L. Effects of resveratrol on glucose control and insulin sensitivity in subjects with type 2 diabetes: Systematic review and meta-analysis. Nutr. Metab. 2017, 14, 60. [Google Scholar] [CrossRef]
- Grzesik, M.; Naparlo, K.; Bartosz, G.; Sadowska-Bartosz, I. Antioxidant properties of catechins: Comparison with other antioxidants. Food Chem. 2018, 241, 480–492. [Google Scholar] [CrossRef]
- Rogelj, A. Antioxidants and their Efficiency in Different Tissues of Invasive Alien Knotweed Species. Master’s Thesis, University of Ljubljana, Ljubljana, Slovenia, 2015. [Google Scholar]
- Pogacnik, L.; Rogelj, A.; Ulrih, N.P. Chemiluminescence Method for Evaluation of Antioxidant Capacities of Different Invasive Knotweed Species. Anal. Lett. 2016, 49, 350–363. [Google Scholar] [CrossRef]
- Pogačnik, L. Bioactive substances from invasive knotweed species. J. EcoAgriTourism 2020, 16, 21–25. [Google Scholar]
- Viktorova, J.; Stranska-Zachariasova, M.; Fenclova, M.; Vitek, L.; Hajslova, J.; Kren, V.; Ruml, T. Complex Evaluation of Antioxidant Capacity of Milk Thistle Dietary Supplements. 2019, 8, 317. [Google Scholar] [CrossRef] [Green Version]
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Fenclova, M.; Stranska-Zachariasova, M.; Vitek, L.; Hajslova, J.; et al. In Silico and In Vitro Studies of Mycotoxins and Their Cocktails; Their Toxicity and Its Mitigation by Silibinin Pre-Treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.; Novakova, A.; Spurna, K.; Hricko, J.; Phung, H.; Viktorova, J.; Stranska, M.; Hajslova, J.; Ruml, T. Antidiabetic Compounds in Stem Juice from Banana. Czech J. Food Sci. 2017, 35, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Benova, B.; Adam, M.; Onderkova, K.; Kralovsky, J.; Krajicek, M. Analysis of selected stilbenes in Polygonum cuspidatum by HPLC coupled with CoulArray detection. J. Sep. Sci. 2008, 31, 2404–2409. [Google Scholar] [CrossRef] [PubMed]
- Vrchotova, N.; Sera, B.; Triska, J. The stilbene and catechin content of the spring sprouts of Reynoutria species. Acta Chromatogr. 2007, 19, 21–28. [Google Scholar]
- Ravagnan, G.; De Filippis, A.; Carteni, M.; De Maria, S.; Cozza, V.; Petrazzuolo, M.; Tufano, M.A.; Donnarumma, G. Polydatin, A Natural Precursor of Resveratrol, Induces beta-Defensin Production and Reduces Inflammatory Response. Inflammation 2013, 36, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Romero-Perez, A.I.; Ibern-Gomez, M.; Lamuela-Raventos, R.M.; de la Torre-Boronat, M.C. Piceid, the major resveratrol derivative in grape juices. J. Agric. Food Chem. 1999, 47, 1533–1536. [Google Scholar] [CrossRef]
- Frantik, T.; Kovarova, M.; Koblihova, H.; Bartunkova, K.; Nyvltova, Z.; Vosatka, M. Production of medically valuable stilbenes and emodin in knotweed. Ind. Crops Prod. 2013, 50, 237–243. [Google Scholar] [CrossRef]
- Tabart, J.; Franck, T.; Kevers, C.; Pincemail, J.; Serteyn, D.; Defraigne, J.O.; Dornmes, J. Antioxidant and anti-inflammatory activities of Ribes nigrum extracts. Food Chem. 2012, 131, 1116–1122. [Google Scholar] [CrossRef]
- Xing, J.L.; Wang, G.; Zhang, Q.X.; Liu, X.M.; Gu, Z.N.; Zhang, H.; Chen, Y.Q.; Chen, W. Determining Antioxidant Activities of Lactobacilli Cell-Free Supernatants by Cellular Antioxidant Assay: A Comparison with Traditional Methods. PLoS ONE 2015, 10, e0119058. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Bishayee, A. Cancer Prevention and Treatment with Resveratrol: From Rodent Studies to Clinical Trials. Cancer Prev. Res. 2009, 2, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Ravipati, A.S.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Bartlett, J.; Smith, P.T.; de la Cruz, M.; Monteiro, M.C.; Melguizo, A.; et al. Anti-fungal and anti-bacterial activities of ethanol extracts of selected traditional Chinese medicinal herbs. Asian Pac. J. Trop. Med. 2013, 6, 673–681. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rhizome Extract | Flower Extract | |||
|---|---|---|---|---|
| Reynoutria Japonica Houtt. | F. x Bohemica | Reynoutria Japonica Houtt. | F. x Bohemica | |
| HEK 293T | 36.7 ± 4.9 ab | 24.9 ± 7.6 a | 92.4 ± 8.5 c | 54.7 ± 8.2 b |
| PaTu | 182.2 ± 48.5 a | 228.0 ± 37.3 a | 430.6 ± 135.8 b | 311.7 ± 58.9 ab |
| HeLa | 36.9 ± 4.2 a | 45.8 ± 7.0 a | 90.2 ± 13.9 b | 59.0 ± 6.1 a |
| HepG2 | 63.0 ± 9.0 a | 63.7 ± 7.6 a | 277.3 ± 41.9 c | 174.7 ± 26.9 b |
| Plant Tissue | Species | IC50 [µg/mL] |
|---|---|---|
| Rhizome | Reynoutria japonica Houtt. | 0.17 ± 0.02 a |
| F. x bohemica | 6.5 ± 2.5 b | |
| Flower | Reynoutria japonica Houtt. | 8.2 ± 0.4 bc |
| F. x bohemica | 25.1 ± 13.3 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pogačnik, L.; Bergant, T.; Skrt, M.; Poklar Ulrih, N.; Viktorová, J.; Ruml, T. In Vitro Comparison of the Bioactivities of Japanese and Bohemian Knotweed Ethanol Extracts. Foods 2020, 9, 544. https://doi.org/10.3390/foods9050544
Pogačnik L, Bergant T, Skrt M, Poklar Ulrih N, Viktorová J, Ruml T. In Vitro Comparison of the Bioactivities of Japanese and Bohemian Knotweed Ethanol Extracts. Foods. 2020; 9(5):544. https://doi.org/10.3390/foods9050544
Chicago/Turabian StylePogačnik, Lea, Tina Bergant, Mihaela Skrt, Nataša Poklar Ulrih, Jitka Viktorová, and Tomáš Ruml. 2020. "In Vitro Comparison of the Bioactivities of Japanese and Bohemian Knotweed Ethanol Extracts" Foods 9, no. 5: 544. https://doi.org/10.3390/foods9050544
APA StylePogačnik, L., Bergant, T., Skrt, M., Poklar Ulrih, N., Viktorová, J., & Ruml, T. (2020). In Vitro Comparison of the Bioactivities of Japanese and Bohemian Knotweed Ethanol Extracts. Foods, 9(5), 544. https://doi.org/10.3390/foods9050544