Effect of Moringa oleifera L. Leaf Powder Addition on the Phenolic Bioaccessibility and on In Vitro Starch Digestibility of Durum Wheat Fresh Pasta

 ,
,  ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Fresh Pasta Sample Preparation
2.2. Moisture Content, Water Activity and Pasta Cooking Properties
2.3. Cooking Process and Experimental Details
2.3.1. In Vitro Static Digestion of Cooked Samples for the Evaluation of the Fate of Polyphenols
2.3.2. Nutritional Starch Fractions Determination
2.3.3. Starch Hydrolysis Index
2.4. Extraction and Characterization of Untargeted Phenolic Profile by UHPLC-ESI/QTOF Mass Spectrometry
2.5. Statistical Analysis
3. Results and Discussion
3.1. Moisture Content, Water Activity and Optimal Cooking Time of Samples
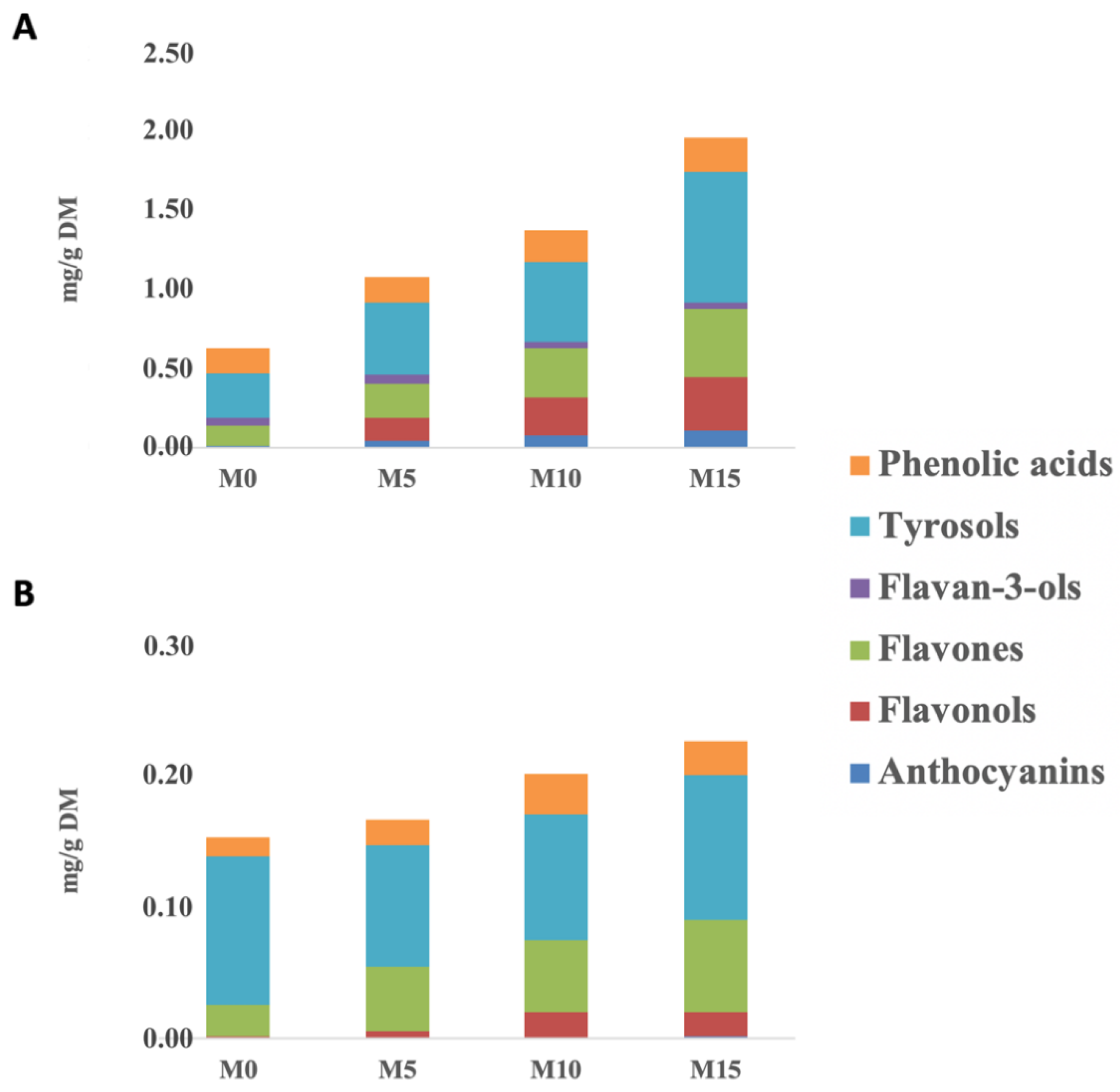
3.2. Free and Bound Phenolic Profiles of Cooked-To-Optimum Samples
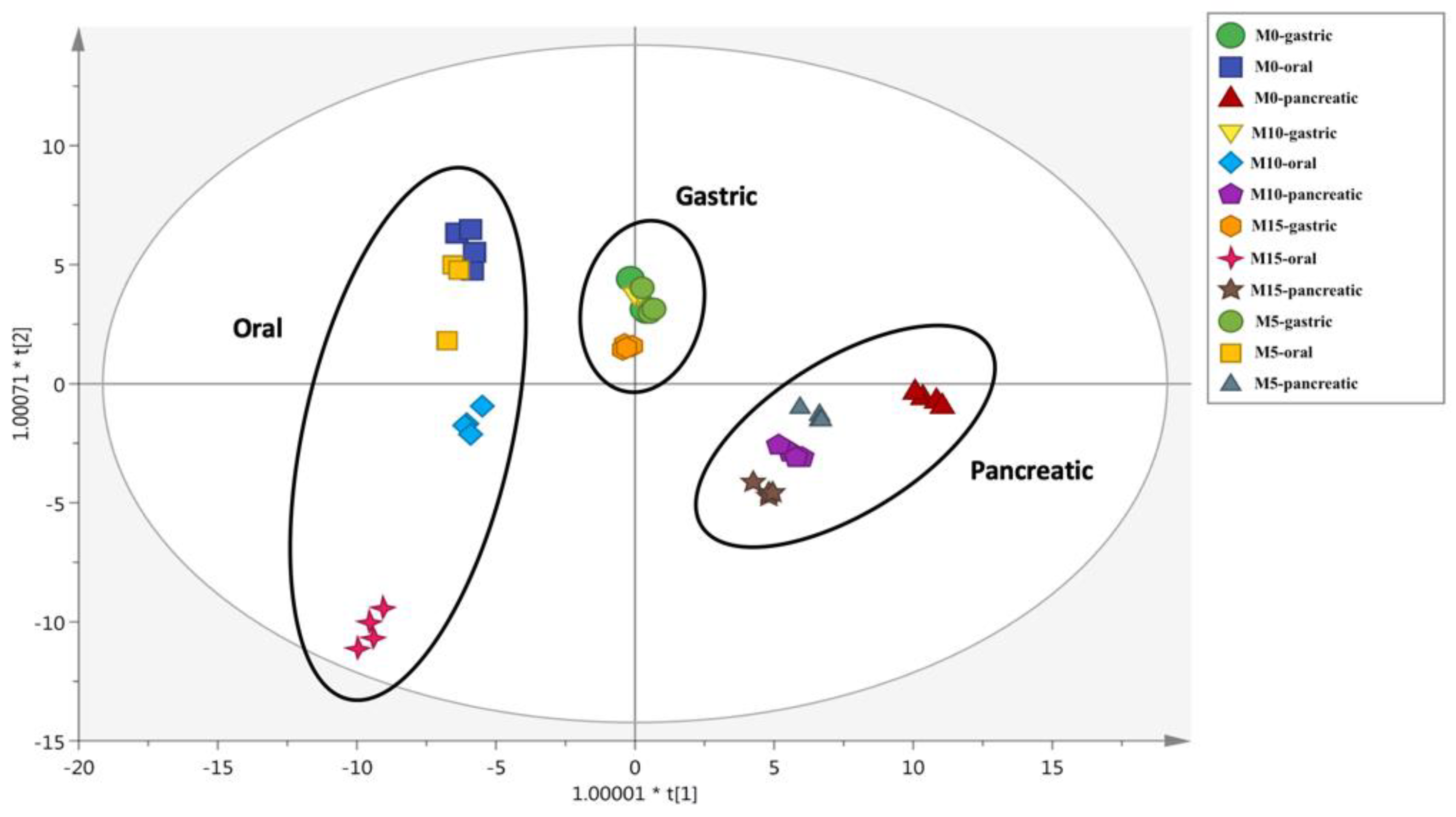
3.3. Changes of Phenolic Profiles during In Vitro Static Digestion
3.4. In Vitro Starch Digestion of Cooked Samples
4. Conclusions
Supplementary Material
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sparvoli, F.; Laureati, M.; Pilu, R.; Pagliarini, E.; Toschi, I.; Giuberti, G.; Fortunati, P.; Daminati, M.G.; Cominelli, E.; Bollini, R. Exploitation of common bean flours with low antinutrient content for making nutritionally enhanced biscuits. Front. Plant Sci. 2016, 7, 00928. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, L.M.; Friesen, V.M. Chapter 32—Impact Evaluation of Food Fortification Programs: Review of Methodological Approaches Used and Opportunities to Strengthen Them. In Food Fortification in a Globalized World; Academic Press: Cambridge, MA, USA, 2018; pp. 305–315. [Google Scholar]
- Spinelli, S.; Padalino, L.; Costa, C.; Del Nobile, M.A.; Conte, A. Food by-products to fortified pasta: A new approach for optimization. J. Clean. Prod. 2019, 215, 985–991. [Google Scholar] [CrossRef]
- Mercier, S.; Moresoli, C.; Mondor, M.; Villeneuve, S.; Marcos, B. A meta-analysis of enriched pasta: What are the effects of enrichment and process specifications on the quality attributes of pasta? Compr. Rev. Food Sci. F 2016, 15, 685–704. [Google Scholar] [CrossRef] [Green Version]
- Simonato, B.; Trevisan, S.; Tolve, R.; Favati, F.; Pasini, G. Pasta fortification with olive pomace: Effects on the technological characteristics and nutritional properties. LWT 2019, 114, 108368. [Google Scholar] [CrossRef]
- Oyeyinka, A.T.; Oyeyinka, S.A. Moringa oleifera as a food fortificant: Recent trends and prospects. J. Saudi Soc. Agric. Sci. 2018, 17, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Rocchetti, G.; Blasi, F.; Montesano, D.; Gisoni, S.; Marcotullio, M.C.; Sabatini, S.; Cossignani, L.; Lucini, L. Impact of conventional/non-conventional extraction methods on the untargeted phenolic profile of Moringa oleifera leaves. Food Res. Int. 2019, 115, 319–327. [Google Scholar] [CrossRef]
- Kardum, N.; Glibetic, M. Chapter Three—Polyphenols and their interactions with other dietary compounds: Implications for human health. Adv. Food Nutr. Res. 2018, 84, 103–144. [Google Scholar]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Rocchetti, G.; Giuberti, G.; Lucini, L. Gluten-free cereal-based food products: The potential of metabolomics to investigate changes in phenolics profile and their in vitro bioaccessibility. Curr. Opin. Food Sci. 2018, 22, 1–8. [Google Scholar] [CrossRef]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food Sci. 2019, 23, 1–15. [Google Scholar] [CrossRef]
- Caicedo-Lopez, L.H.; Luzardo-Ocampo, I.; Cuellar-Nunez, M.L.; Campos-Vega, R.; Mendoza, S.; Loarca-Pina, G. Effect of the in vitro gastrointestinal digestion on free-phenolic compounds and mono/oligosaccharides from Moringa oleifera leaves: Bioaccessibility, intestinal permeability and antioxidant capacity. Food Res. Int. 2019, 120, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists. Official Methods of Analysis, 17th ed.; AOAC, Inc.: Arlington, VA, USA, 2000. [Google Scholar]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food - an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46, 33–50. [Google Scholar]
- Granfeldt, Y.; Björck, I.; Drews, A.; Tovar, J. An in vitro procedure based on chewing to predict metabolic response to starch in cereal and legume products. Eur. J. Clin. Nutr. 1992, 46, 649–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocchetti, G.; Lucini, L.; Giuberti, G.; Bhumireddy, S.R.; Mandal, R.; Trevisan, M.; Wishart, D. Transformation of polyphenols found in pigmented gluten-free flours during in vitro large intestinal fermentation. Food Chem. 2019, 298, 125068. [Google Scholar] [CrossRef] [PubMed]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. Gigascience 2013, 2, 13. [Google Scholar] [CrossRef]
- Rocchetti, G.; Chiodelli, G.; Giuberti, G.; Lucini, L. Bioaccessibility of phenolic compounds following in vitro large intestine fermentation of nuts for human consumption. Food Chem. 2018, 245, 633–640. [Google Scholar] [CrossRef]
- Rakhesh, N.; Fellows, C.M.; Sissons, M. Evaluation of the technological and sensory properties of durum wheat spaghetti enriched with different dietary fibres. J. Sci. Food Agric. 2015, 95, 2–11. [Google Scholar] [CrossRef]
- Shahidi, F.; Peng, H. Bioaccessibility and bioavailability of phenolic compounds. J. Food Bioact. 2018, 4, 11–68. [Google Scholar] [CrossRef] [Green Version]
- Fares, C.; Platani, C.; Baiano, A.; Menga, V. Effect of processing and cooking on phenolic acid profile and antioxidant capacity of durum wheat pasta enriched with debranning fractions of wheat. Food Chem. 2010, 119, 1023–1029. [Google Scholar] [CrossRef]
- Žilić, S. Phenolic compounds of wheat: Their content, antioxidant capacity and bioaccessibility. MOJ Food Process. Technol. 2016, 2, 85–89. [Google Scholar] [CrossRef]
- Sun-Waterhouse, D.; Jin, D.; Waterhouse, G.I.N. Effect of adding elderberry juice concentrate on the quality attributes, polyphenol contents and antioxidant activity of three fibre-enriched pastas. Food Res. Int. 2013, 54, 781–789. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Chiodelli, G.; Giuberti, G.; Gallo, A.; Masoero, F.; Trevisan, M. Phenolic profile and fermentation patterns of different commercial gluten-free pasta during in vitro large intestine fermentation. Food Res. Int. 2017, 197, 78–86. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Goncalves, S.; Heranz, D.; Heredia, F.J.; Romano, A. Effects of in vitro gastrointestinal digestion on phenolic compounds and antioxidant activity of different white winemaking byproducts extracts. Food Res. Int. 2018, 109, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Ti, H.; Zhang, R.; Li, Q.; Wei, Z.; Zhang, M. Effects of cooking and in vitro digestion of rice on phenolic profiles and antioxidant activity. Food Res. Int. 2015, 76, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Aboul-Enein, B.; Bernstein, J.; Marchlewicz, M. Dietary alkylresorcinols and cancer prevention: A systematic review. Eur. Food Res. Technol. 2017, 243, 1693–1710. [Google Scholar] [CrossRef] [Green Version]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant starch—A review. Compr. Rev. Food Sci. F 2006, 5, 1–17. [Google Scholar] [CrossRef]
- Jia, M.; Yu, Q.; Chen, J.; He, Z.; Chen, Y.; Xie, J.; Nie, S.; Xie, M. Physical quality and in vitro starch digestibility of biscuits as affected by addition of soluble dietary fiber from defatted rice bran. Food Hydrocoll. 2020, 99, 105349. [Google Scholar] [CrossRef]
- Fuentes-Zaragoza, E.; Sánchez-Zapata, E.; Sendra, E.; Sayas, E.; Navarro, C.; Fernández-López, J.; Pérez-Alvarez, J.A. Resistant starch as prebiotic: A review. Starch/Stärke 2011, 63, 406–415. [Google Scholar] [CrossRef]
- Espinosa-Solis, W.; Zamudio-Flores, P.B.; Tirado-Gallegos, J.M.; Ramírez-Mancinas, S.; Olivas-Orozco, G.I.; Espino-Díaz, M.; Hernández-González, M.; García-Cano, V.G.; Sánchez-Ortíz, O.; Buenrostro-Figueroa, J.J.; et al. Evaluation of cooking quality, nutritional and texture characteristics of pasta added with oat bran and apple flour. Foods 2019, 8, 299. [Google Scholar] [CrossRef] [Green Version]
- Xou, W.; Sissons, M.; Gidley, M.J.; Gilbert, R.G.; Warren, F.J. Combined techniques for characterising pasta structure reveals how the gluten network slows enzymic digestion rate. Food Chem. 2015, 188, 559–568. [Google Scholar]
- Yue, P.; Rayas-Duarte, P.; Elias, E. Effect of drying temperature on physicochemical properties of starch isolated from pasta. Cereal. Chem. 1999, 76, 541–547. [Google Scholar] [CrossRef]
- Singh, J.; Dartois, A.; Kaur, L. Starch digestibility in food matrix: A review. Trends Food Sci. Technol. 2010, 21, 168–180. [Google Scholar] [CrossRef]
- Dachana, K.B.; Rajiv, J.; Indrani, D.; Prakash, J. Effect of dried Moringa (Moringa Oleifera Lam) leaves on rheological, microstructural, nutritional, textural and organoleptic characteristics of cookies. J. Food Qual. 2010, 33, 660–677. [Google Scholar] [CrossRef]
- Giuberti, G.; Rocchetti, G.; Sigolo, S.; Fortunati, P.; Lucini, L.; Gallo, A. Exploitation of alfalfa seed (Medicago sativa L.) flour into gluten-free rice cookies: Nutritional, antioxidant and quality characteristics. Food Chem. 2018, 239, 679–687. [Google Scholar] [CrossRef]
- Zhu, F. Interactions between starch and phenolic compound. Trends Food Sci. Technol. 2015, 43, 129–143. [Google Scholar] [CrossRef]
- Rocchetti, G.; Giuberti, G.; Gallo, A.; Bernardi, J.; Marocco, A.; Lucini, L. Effect of dietary polyphenols on the in vitro starch digestibility of pigmented maize varieties under cooking conditions. Food Res. Int. 2018, 108, 183–191. [Google Scholar] [CrossRef]
- Khan, W.; Parveen, R.; Chester, K.; Parveen, S.; Ahmad, S. Hypoglycemic potential of aqueous extract of Moringa oleifera leaf and in vivo GC-MS metabolomics. Front. Pharmacol. 2017, 8, 577. [Google Scholar] [CrossRef] [Green Version]
- Leone, A.; Bertoli, S.; Di Lello, S.; Bassoli, A.; Ravasenghi, S.; Borgonovo, G.; Forlani, F.; Battezzati, A. Effect of Moringa oleifera leaf powder on postprandial blood glucose response: In vivo study on Saharawi people living in refugee camps. Nutrients 2018, 10, 1494. [Google Scholar] [CrossRef] [Green Version]
- Ademiluyi, A.O.; Aladeselu, O.H.; Oboh, G.; Boligon, A.A. Drying alters the phenolic constituents, antioxidant properties, α-amylase, and α-glucosidase inhibitory properties of Moringa (Moringa oleifera) leaf. Food Sci. Nutr. 2018, 6, 2123–2133. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Phenolic Subclasses | Pasta Samples | TPC Cooked Samples (mg Eq./100 g) | % Bioaccessibility | ||
|---|---|---|---|---|---|
| Oral | Gastric | Pancreatic | |||
| Anthocyanins | M0 | 0.87 ± 0.04 a | 0.31 | 0.45 | nd |
| M5 | 4.47 ± 0.22 b | 0.14 | 0.13 | 0.07 | |
| M10 | 7.43 ± 0.36 c | 0.17 | 0.19 | 0.37 | |
| M15 | 10.41 ± 0.50 d | 0.22 | 0.20 | 0.27 | |
| Flavonols | M0 | 0.19 ± 0.01 a | nd | nd | nd |
| M5 | 14.74 ± 0.72 b | 0.04 | 0.22 | 1.01 | |
| M10 | 26.06 ± 1.20 c | 0.09 | 0.26 | 1.13 | |
| M15 | 35.73 ± 1.80 d | 0.16 | 0.44 | 1.14 | |
| Flavones | M0 | 15.19 ± 0.71 a | 13.08 | 9.31 | 5.53 |
| M5 | 26.59 ± 1.33 b | 7.48 | 4.69 | 28.26 | |
| M10 | 36.70 ± 1.82 c | 5.01 | 3.10 | 17.27 | |
| M15 | 50.28 ± 2.41 d | 3.23 | 2.13 | 15.16 | |
| Flavan-3-ols | M0 | 4.36 ± 0.18 | 14.45 | nd | nd |
| M5 | 5.20 ± 0.21 | 7.05 | nd | nd | |
| M10 | 3.91 ± 0.12 | 5.52 | nd | nd | |
| M15 | 4.01 ± 0.13 | 8.50 | nd | nd | |
| Tyrosols | M0 | 39.25 ± 1.90 a | 1.92 | 1.89 | 29.21 |
| M5 | 55.11 ± 2.71 b | 1.41 | 1.93 | 13.80 | |
| M10 | 60.11 ± 2.98 b | 1.30 | 1.73 | 8.32 | |
| M15 | 93.53 ± 4.65 c | 0.87 | 1.17 | 3.01 | |
| Phenolic acids | M0 | 17.96 ± 0.86 a | 3.30 | 3.10 | 12.45 |
| M5 | 17.95 ± 0.89 a | 2.42 | 4.10 | 12.36 | |
| M10 | 23.50 ± 1.18 b | 1.81 | 3.09 | 8.97 | |
| M15 | 24.86 ± 1.25 b | 1.86 | 3.22 | 8.28 | |
| Substitution with MOLP | ||||
|---|---|---|---|---|
| M0 | M5 | M10 | M15 | |
| Rapidly digestible starch | 44.3 ± 0.31 a | 43.8 ± 0.79 a | 38.1 ± 1.76 b | 34.1 ± 3.49 b |
| Slowly digestible starch | 16.8 ± 0.70 a | 16.8 ± 0.67 a | 18.1 ± 0.20 b | 20.8 ± 0.67 c |
| Resistant starch | 2.1 ± 0.26 a | 1.4 ± 0.04 b | 1.3 ± 0.01 c | 1.1 ± 0.04 d |
| Total starch | 63.1 ± 1.33 b | 62.1 ± 1.77 b | 57.7 ± 1.21 a | 55.9 ± 1.44 a |
| HI 1 | 47.4 ± 1.05 a | 45.4 ± 1.32 ab | 43.9 ± 1.21 ab | 41.8 ± 0.81 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocchetti, G.; Rizzi, C.; Pasini, G.; Lucini, L.; Giuberti, G.; Simonato, B. Effect of Moringa oleifera L. Leaf Powder Addition on the Phenolic Bioaccessibility and on In Vitro Starch Digestibility of Durum Wheat Fresh Pasta. Foods 2020, 9, 628. https://doi.org/10.3390/foods9050628
Rocchetti G, Rizzi C, Pasini G, Lucini L, Giuberti G, Simonato B. Effect of Moringa oleifera L. Leaf Powder Addition on the Phenolic Bioaccessibility and on In Vitro Starch Digestibility of Durum Wheat Fresh Pasta. Foods. 2020; 9(5):628. https://doi.org/10.3390/foods9050628
Chicago/Turabian StyleRocchetti, Gabriele, Corrado Rizzi, Gabriella Pasini, Luigi Lucini, Gianluca Giuberti, and Barbara Simonato. 2020. "Effect of Moringa oleifera L. Leaf Powder Addition on the Phenolic Bioaccessibility and on In Vitro Starch Digestibility of Durum Wheat Fresh Pasta" Foods 9, no. 5: 628. https://doi.org/10.3390/foods9050628
APA StyleRocchetti, G., Rizzi, C., Pasini, G., Lucini, L., Giuberti, G., & Simonato, B. (2020). Effect of Moringa oleifera L. Leaf Powder Addition on the Phenolic Bioaccessibility and on In Vitro Starch Digestibility of Durum Wheat Fresh Pasta. Foods, 9(5), 628. https://doi.org/10.3390/foods9050628