Modelling of the Behaviour of Salmonella enterica serovar Reading on Commercial Fresh-Cut Iceberg Lettuce Stored at Different Temperatures

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Microbiological Analysis
2.1.1. Preparation of Inoculum
2.1.2. Inoculation of Lettuce Bags
2.1.3. Bacterial Analyses
2.2. Microbial Modelling
2.2.1. Growth Model
2.2.2. Survival Kinetic Model
2.2.3. Goodness of Fit of the Growth and Survival Models
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jideani, A.I.O.; Anyasi, T.A.; Mchau, G.R.A.; Udoro, E.O.; Onipe, O.O. Processing and preservation of fresh-cut fruit and vegetable products. In Postharvest Handling; IntechOpen: London, UK, 2017; pp. 47–73. [Google Scholar] [CrossRef] [Green Version]
- Baselice, A.; Colantuoni, F.; Lass, D.A.; Nardone, G.; Stasi, A. Trends in EU consumers’ attitude towards fresh-cut fruit and vegetables. Food Qual. Prefer. 2017, 59, 87–96. [Google Scholar] [CrossRef]
- Steele, M.; Odumeru, J. Irrigation water as source of foodborne pathogens on fruit and vegetables. J. Food Prot. 2004, 67, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- De Corato, U. Improving the shelf-life and quality of fresh and minimally-processed fruits and vegetables for a modern food industry: A comprehensive critical review from the traditional technologies into the most promising advancements. Crit. Rev. Food Sci. Nutr. 2020, 60, 940–975. [Google Scholar] [CrossRef]
- Oms-Oliu, G.; Rojas-Graü, M.A.; González, L.A.; Varela, P.; Soliva-Fortuny, R.; Hernando, M.I.H.; Munuera, I.P.; Fiszman, S.; Martín-Belloso, O. Recent approaches using chemical treatments to preserve quality of fresh-cut fruit: A review. Postharvest Biol. Technol. 2010, 57, 139–148. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, M.; Bhandari, B.; Gao, Z. Recent developments in novel shelf life extension technologies of fresh-cut fruits and vegetables. Trends Food Sci. Technol. 2017, 64, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Murray, K.; Wu, F.; Shi, J.; Jun Xue, S.; Warriner, K. Challenges in the microbiological food safety of fresh produce: Limitations of post-harvest washing and the need for alternative interventions. Food Qual. Saf. 2017, 1, 289–301. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union one health 2018 zoonoses report. EFSA J. 2019, 17, e05926. [Google Scholar] [CrossRef] [Green Version]
- Callejón, R.M.; Rodríguez-Naranjo, M.I.; Ubeda, C.; Hornedo-Ortega, R.; Garcia-Parrilla, M.C.; Troncoso, A.M. Reported foodborne outbreaks due to fresh produce in the united states and European Union: Trends and causes. Foodborne Pathog. Dis. 2015, 12, 32–38. [Google Scholar] [CrossRef]
- Da Silva Felício, M.T.; Hald, T.; Liebana, E.; Allende, A.; Hugas, M.; Nguyen-The, C.; Johannessen, G.S.; Niskanen, T.; Uyttendaele, M.; McLauchlin, J. Risk ranking of pathogens in ready-to-eat unprocessed foods of non-animal origin (FoNAO) in the EU: Initial evaluation using outbreak data (2007–2011). Int. J. Food Microbiol. 2015, 195, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Herman, K.M.; Hall, A.J.; Gould, L.H. Outbreaks attributed to fresh leafy vegetables, United States, 1973–2012. Epidemiol. Infect. 2015, 143, 3011–3021. [Google Scholar] [CrossRef] [Green Version]
- Tanguay, F.; Vrbova, L.; Anderson, M.; Whitfield, Y.; Macdonald, L.; Tschetter, L.; Hexemer, A. Outbreak of Salmonella Reading in persons of Eastern Mediterranean origin in Canada, 2014–2015. Can. Commun. Dis. Rep. 2017, 43, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Lienemann, T.; Niskanen, T.; Guedes, S.; Siitonen, A.; Kuusi, M.; Rimhanen-Finne, R. Iceberg lettuce as suggested source of a nationwide outbreak caused by two Salmonella serotypes, Newport and Reading, in Finland in 2008. J. Food Prot. 2011, 74, 1035–1040. [Google Scholar] [CrossRef]
- Robertson, L.J.; Johannessen, G.S.; Gjerde, B.K.; Loncarevic, S. Microbiological analysis of seed sprouts in Norway. Int. J. Food Microbiol. 2002, 75, 119–126. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Salmonella: Outbreak of Multidrug-Resistant Salmonella Infections Linked to Raw Turkey Products; Final Update; US Department of Health and Human Services: Atlanta, GA, USA, 2019. Available online: https://www.cdc.gov/salmonella/reading-07-18/index.html (accessed on 17 December 2019).
- Koseki, S.; Isobe, S. Prediction of pathogen growth on iceberg lettuce under real temperature history during distribution from farm to table. Int. J. Food Microbiol. 2005, 104, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Guo, M.; Buchanan, R.L.; Schaffner, D.W.; Pradhan, A.K. Development of growth and survival models for Salmonella and Listeria monocytogenes during non-isothermal time-temperature profiles in leafy greens. Food Control 2017, 71, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Puerta-Gomez, A.F.; Moreira, R.G.; Kim, J.; Castell-Perez, E. Modeling the growth rates of Escherichia coli spp. and Salmonella Typhimurium LT2 in baby spinach leaves under slow cooling. Food Control 2013, 29, 11–17. [Google Scholar] [CrossRef]
- Sant’Ana, A.S.; Franco, B.D.G.M.; Schaffner, D.W. Modeling the growth rate and lag time of different strains of Salmonella enterica and Listeria monocytogenes in ready-to-eat lettuce. Food Microbiol. 2012, 30, 267–273. [Google Scholar] [CrossRef]
- Veys, O.; de Oliveira Elias, S.; Sampers, I.; Tondo, E.C. Modelling the growth of Salmonella spp. and Escherichia coli O157 on lettuce. Procedia Food Sci. 2016, 7, 168–172. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Bae, Y.M.; Jung, S.Y.; Cha, M.H.; Ryu, K.; Park, K.H.; Lee, S.Y. Predictive modeling for the growth of Listeria monocytogenes and Salmonella Typhimurium on fresh-cut cabbage at various temperatures. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 631–638. [Google Scholar] [CrossRef]
- Carrasco, E.; Pérez-Rodríguez, F.; Valero, A.; García-Gimeno, R.M.; Zurera, G. Growth of Listeria monocytogenes on shredded, ready-to-eat iceberg lettuce. Food Control 2008, 19, 487–494. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]
- Dabadé, D.S.; Azokpota, P.; Nout, M.J.R.; Hounhouigan, D.J.; Zwietering, M.H.; den Besten, H.M.W. Prediction of spoilage of tropical shrimp (Penaeus notialis) under dynamic temperature regimes. Int. J. Food Microbiol. 2015, 210, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, H.M.W.; Mataragas, M.; Moezelaar, R.; Abee, T.; Zwietering, M.H. Quantification of the effects of salt stress and physiological state on thermotolerance of Bacillus cereus ATCC 10987 and ATCC 14579. Appl. Environ. Microbiol. 2006, 72, 5884–5894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafart, P.; Couvert, O.; Gaillard, S.; Leguerinel, I. On calculating sterility in thermal preservation methods: Application of the Weibull frequency distribution model. Int. J. Food Microbiol. 2002, 72, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Ioannidis, A.G.; Kerckhof, F.M.; Riahi Drif, Y.; Vanderroost, M.; Boon, N.; Ragaert, P.; De Meulenaer, B.; Devlieghere, F. Characterization of spoilage markers in modified atmosphere packaged iceberg lettuce. Int. J. Food Microbiol. 2018, 279, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kakiomenou, K.; Tassou, C.; Nychas, G.J. Survival of Salmonella enteritidis and Listeria monocytogenes on salad vegetables. World J. Microbiol. Biotechnol. 1998, 14, 383–387. [Google Scholar] [CrossRef]
- Paillart, M.J.M.; van der Vossen, J.M.B.M.; Levin, E.; Lommen, E.; Otma, E.C.; Snels, J.C.M.A.; Woltering, E.J. Bacterial population dynamics and sensorial quality loss in modified atmosphere packed fresh-cut iceberg lettuce. Postharvest Biol. Technol. 2017, 124, 91–99. [Google Scholar] [CrossRef]
- De Oliveira Elias, S.; Noronha, T.B.; Tondo, E.C. Assessment of Salmonella spp. and Escherichia coli O157:H7 growth on lettuce exposed to isothermal and non-isothermal conditions. Food Microbiol. 2018, 72, 206–213. [Google Scholar] [CrossRef]
- Chang, J.M.; Fang, T.J. Survival of Escherichia coli O157:H7 and Salmonella enterica serovars Typhimurium in iceberg lettuce and the antimicrobial effect of rice vinegar against E. coli O157:H7. Food Microbiol. 2007, 24, 745–751. [Google Scholar] [CrossRef]
- Delbeke, S.; Ceuppens, S.; Jacxsens, L.; Uyttendaele, M. Survival of Salmonella and Escherichia coli O157:H7 on strawberries, basil, and other leafy greens during storage. J. Food Prot. 2015, 78, 652–660. [Google Scholar] [CrossRef]


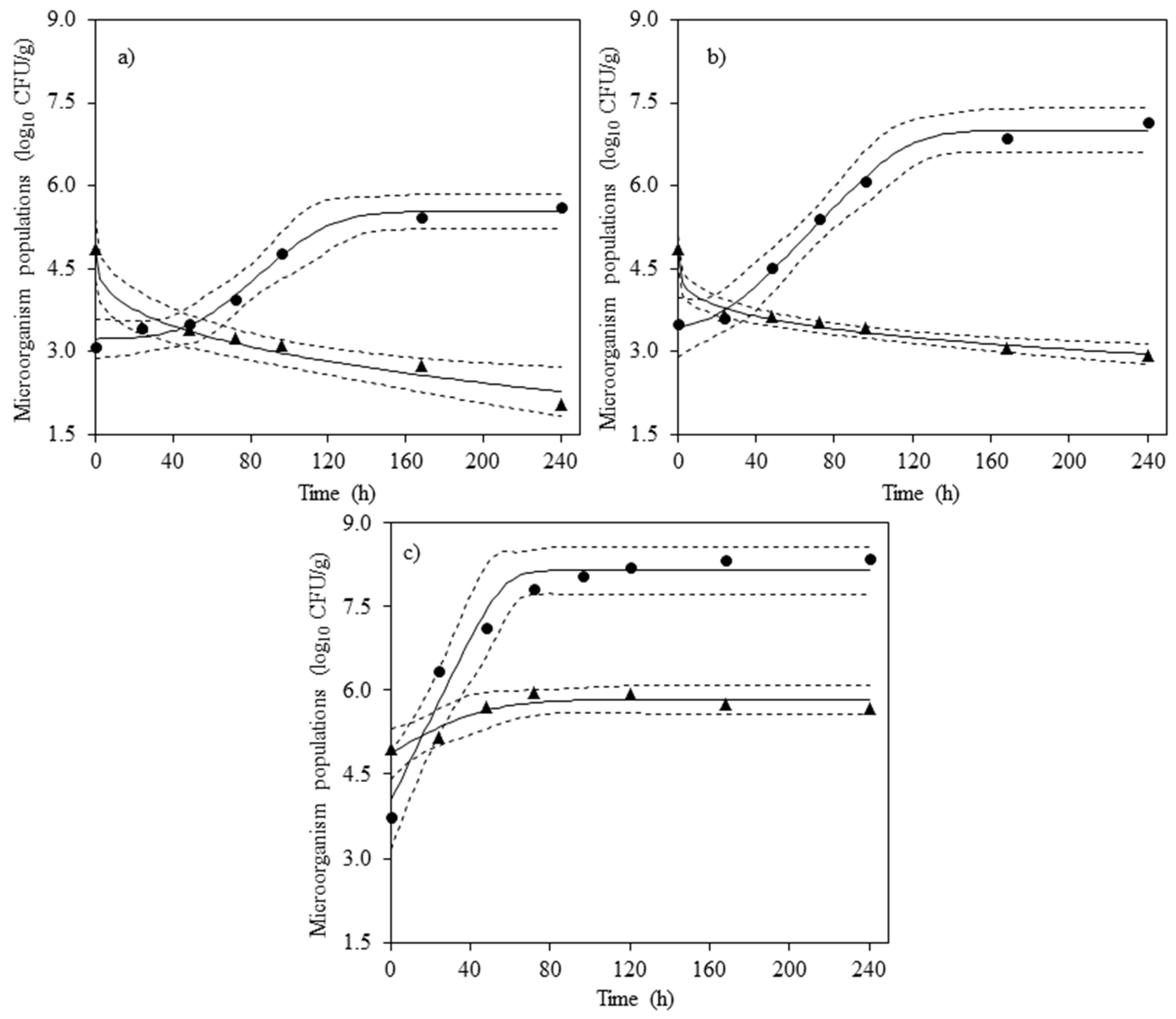{kind=link}
| Lactic Acid Bacteria (log10 CFU/g) a | Salmonella Reading (log10 CFU/g) a | ||||||
|---|---|---|---|---|---|---|---|
| Storage Temperature (°C) | 4 | 8 | 15 | 4 | 8 | 15 | |
| Storage Time (day) | 0 | 3.10 ± 0.06 | 3.51 ± 0.07 | 3.75 ± 0.35 | 4.87 ± 0.04 | 4.86 ± 0.05 | 4.95 ± 0.11 |
| 1 | 3.42 ± 0.18 | 3.61 ± 0.08 | 6.34 ± 0.52 | 3.46 ± 0.20 | 3.67 ± 0.11 | 5.16 ± 0.06 | |
| 2 | 3.49 ± 0.19 | 4.53 ± 0.18 | 7.13 ± 0.69 | 3.39 ± 0.21 | 3.62 ± 0.15 | 5.70 ± 0.13 | |
| 3 | 3.96 ± 0.50 | 5.42 ± 0.18 | 7.82 ± 0.05 | 3.25 ± 0.16 | 3.53 ± 0.14 | 5.95 ± 0.40 | |
| 4 | 4.78 ± 0.64 | 6.08 ± 0.01 | 8.04 ± 0.05 | 3.13 ± 012 | 3.42 ± 0.13 | - | |
| 7 | 5.44 ± 0.73 | 6.85 ± 0.62 | 8.19 ± 0.13 | 2.74 ± 035 | 3.07 ± 0.30 | 5.74 ± 0.22 | |
| 10 | 5.62 ± 0.17 | 7.16 ± 0.34 | 8.36 ± 0.13 | 2.06 ± 0.08 | 2.93 ± 0.25 | 5.68 ± 0.29 | |
| Bacteria | Temp. (°C) | Parameters a | MSE b | R2 c | |||
|---|---|---|---|---|---|---|---|
| N0 (log10 CFU/g) | Nmax (log10 CFU/g) | μmax (h−1) | λ (h) | ||||
| LAB | 4 | 3.23 ± 0.110 | 5.54 ± 0.102 | 0.080 ± 0.018 | 51.1 ± 9.5 | 0.020 | 0.981 |
| 8 | 3.45 ± 0.168 | 7.00 ± 0.125 | 0.087 ± 0.012 | 21.9 ± 9.4 | 0.031 | 0.986 | |
| 15 | 4.04 ± 0.335 | 8.14 ± 0.166 | 0.168 ± 0.028 | - | 0.132 | 0.946 | |
| S. Reading | 15 | 4.87 ± 0.162 | 5.83 ± 0.94 | 0.056 ± 0.005 | - | 0.029 | 0.806 |
| Bacteria | Temp. (°C) | Parameters a | MSE b | R2 c | ||
|---|---|---|---|---|---|---|
| N0 (log10 CFU/g) | delta | P | ||||
| Salmonella Reading | 4 | 4.84 ± 0.201 | 16.03 ± 10.5 | 0.348 ± 0.075 | 0.041 | 0.944 |
| 8 | 4.85 ± 0.087 | 18.81 ± 7.81 | 0.250 ± 0.036 | 0.007 | 0.981 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarlak, F.; Johannessen, G.; Bascón Villegas, I.; Bolívar, A.; Posada-Izquierdo, G.D.; Pérez-Rodríguez, F. Modelling of the Behaviour of Salmonella enterica serovar Reading on Commercial Fresh-Cut Iceberg Lettuce Stored at Different Temperatures. Foods 2020, 9, 946. https://doi.org/10.3390/foods9070946
Tarlak F, Johannessen G, Bascón Villegas I, Bolívar A, Posada-Izquierdo GD, Pérez-Rodríguez F. Modelling of the Behaviour of Salmonella enterica serovar Reading on Commercial Fresh-Cut Iceberg Lettuce Stored at Different Temperatures. Foods. 2020; 9(7):946. https://doi.org/10.3390/foods9070946
Chicago/Turabian StyleTarlak, Fatih, Gro Johannessen, Isabel Bascón Villegas, Araceli Bolívar, Guiomar Denisse Posada-Izquierdo, and Fernando Pérez-Rodríguez. 2020. "Modelling of the Behaviour of Salmonella enterica serovar Reading on Commercial Fresh-Cut Iceberg Lettuce Stored at Different Temperatures" Foods 9, no. 7: 946. https://doi.org/10.3390/foods9070946
APA StyleTarlak, F., Johannessen, G., Bascón Villegas, I., Bolívar, A., Posada-Izquierdo, G. D., & Pérez-Rodríguez, F. (2020). Modelling of the Behaviour of Salmonella enterica serovar Reading on Commercial Fresh-Cut Iceberg Lettuce Stored at Different Temperatures. Foods, 9(7), 946. https://doi.org/10.3390/foods9070946