Muscle, Ageing and Temperature Influence the Changes in Texture, Cooking Loss and Shrinkage of Cooked Beef

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Muscle Procurement, Allocation to Ageing Treatment, Measurement of pH and Sampling
2.2. Characterization of the Muscles
2.2.1. Sarcomere Length Measurement
2.2.2. Collagen Content Determination
2.3. Physical Changes during Cooking
2.3.1. Method of Cooking, Measurement of Dimensions and Weight
Subsampling
2.3.2. WBSF
2.3.3. Calculation of Cooking Loss, Longitudinal and Transverse Shrinkage
2.3.4. Diameter of Fibre Fragments Obtained by Homogenization of Cooked Meat
2.4. Data Analysis
3. Results
3.1. Muscle Characterization
3.2. Physical Changes of Bovine Muscles with Ageing and Cooking
3.2.1. WBSF
3.2.2. Cooking Loss
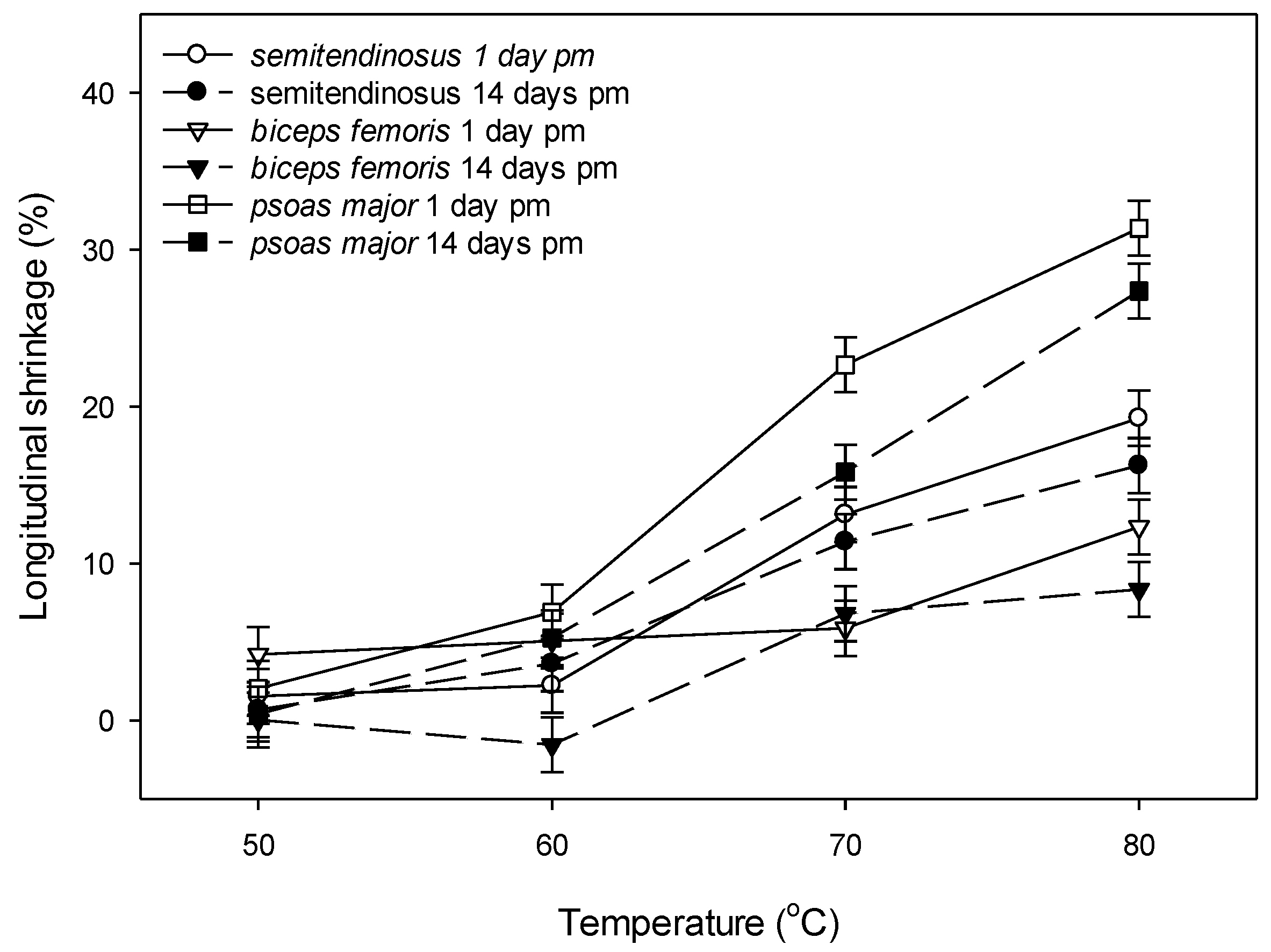
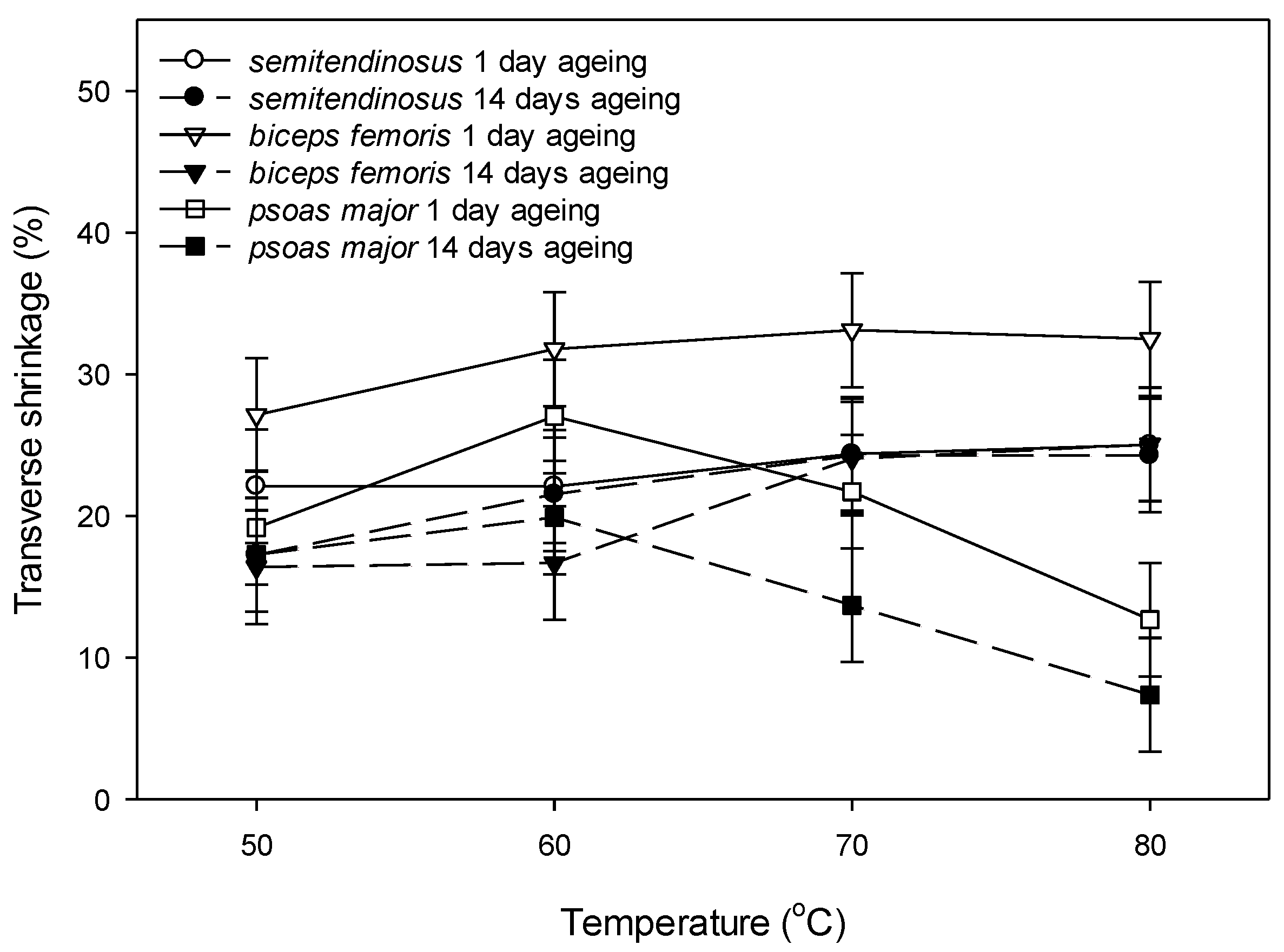
3.2.3. Shrinkage
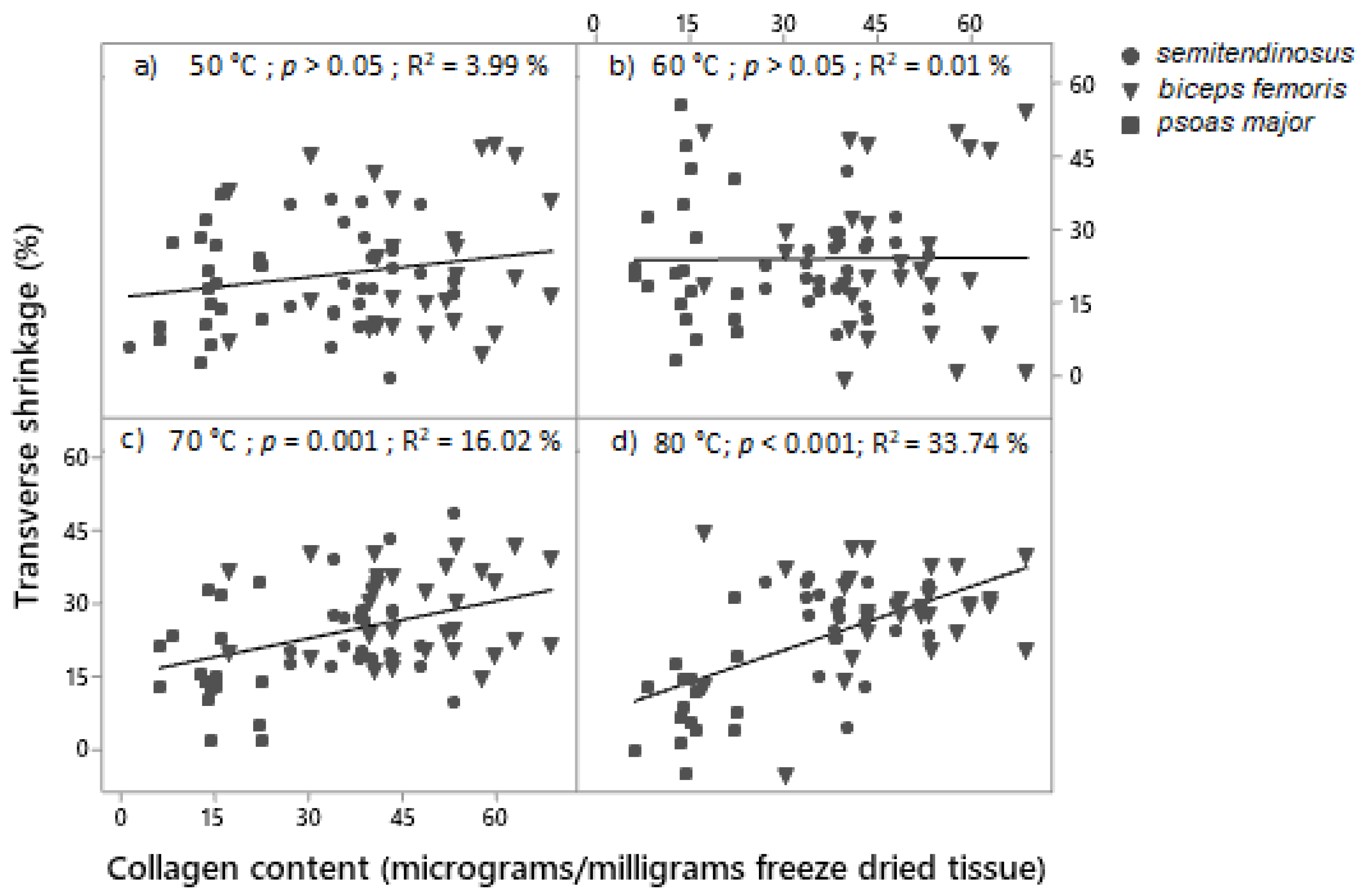
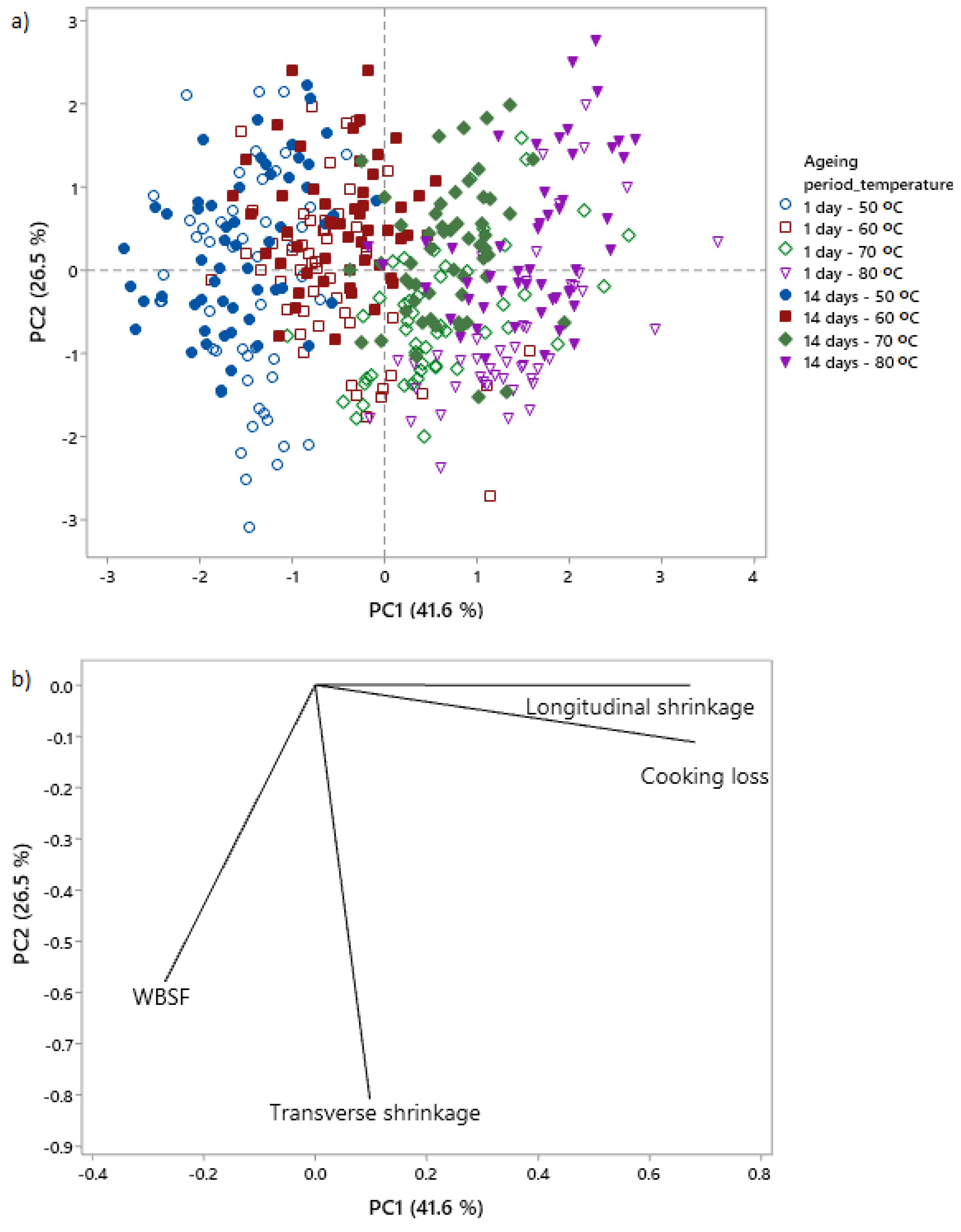
3.3. Multivariate Analysis of the Relationships between Shrinkage and WBSF/Cooking Loss: Ageing and Cooking Temperature
4. Discussion
4.1. Changes in WBSF with Ageing and Cooking
4.2. Changes in Cooking Loss with Ageing and Cooking
4.3. Changes in Shrinkage with Ageing and Cooking
4.4. Relationships between WBSF, Cooking Loss and Shrinkage as Affected by Ageing and Cooking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davey, C.L.; Gilbert, K.V. Temperature-dependent cooking toughness in beef. J. Sci. Food Agric. 1974, 25, 931–938. [Google Scholar] [CrossRef]
- Tornberg, E. Effects of heat on meat proteins—Implications on structure and quality of meat products. Meat Sci. 2005, 70, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Bendall, J.R.; Restall, D.J. The cooking of single myofibres, small myofibre bundles and muscle strips from beef M. psoas and M. sternomandibularis muscles at varying heating rates and temperatures. Meat Sci. 1983, 8, 93–117. [Google Scholar] [CrossRef]
- Hostetler, R.L.; Landmann, W.A. Photomicrographic studies of dynamic changes in muscle fiber fragments. 1. Effect of various heat treatments on length, width and birefringence. J. Food Sci. 1968, 33, 468–470. [Google Scholar] [CrossRef]
- Hearne, L.E.; Penfield, M.P.; Goertz, G.E. Heating effects on bovine semitendinosus: Shear, muscle fiber measurements, and cooking losses. J. Food Sci. 1978, 43, 10–12. [Google Scholar] [CrossRef]
- Palka, K.; Daun, H. Changes in texture, cooking losses, and myofibrillar structure of bovine M. semitendinosus during heating. Meat Sci. 1999, 51, 237–243. [Google Scholar] [CrossRef]
- Purslow, P.P.; Oiseth, S.; Hughes, J.; Warner, R.D. The structural basis of cooking loss in beef: Variations with temperature and ageing. Food Res. Int. 2016, 89, 739–748. [Google Scholar] [CrossRef]
- Offer, G.; Trinick, J. On the mechanism of water holding in meat: The swelling and shrinking of myofibrils. Meat Sci. 1983, 8, 245–281. [Google Scholar] [CrossRef]
- Astruc, T.; Peyrin, F.; Vénien, A.; Labas, R.; Abrantes, M.; Dumas, P.; Jamme, F. In situ thermal denaturation of myofibre sub-type proteins studied by immunohistofluorescence and synchrotron radiation FT-IR microspectroscopy. Food Chem. 2012, 134, 1044–1051. [Google Scholar] [CrossRef]
- Bouton, P.E.; Harris, P.V.; Shorthose, W.R. Dimensional changes in meat during cooking. J. Texture Stud. 1976, 7, 179–192. [Google Scholar] [CrossRef]
- Platter, W.J.; Tatum, J.D.; Belk, K.E.; Chapman, P.L.; Scanga, J.A.; Smith, G.C. Relationships of consumer sensory ratings, marbling score, and shear force value to consumer acceptance of beef strip loin steaks. J. Anim. Sci. 2003, 81, 2741–2750. [Google Scholar] [CrossRef] [Green Version]
- Koohmaraie, M. Biochemical factors regulating the toughening and tenderization processes of meat. Meat Sci. 1996, 43, 193–201. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Zhang, W.; Lonergan, S.M. Biochemistry of postmortem muscle—Lessons on mechanisms of meat tenderization. Meat Sci. 2010, 86, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Mendez, C.H.; Becila, S.; Boudjellal, A.; Ouali, A. Meat ageing: Reconsideration of the current concept. Trends Food Sci. Technol. 2006, 17, 394–405. [Google Scholar] [CrossRef]
- Sentandreu, M.; Coulis, G.; Ouali, A. Role of muscle endopeptidases and their inhibitors in meat tenderness. Trends Food Sci. Technol. 2002, 13, 400–421. [Google Scholar] [CrossRef]
- Davey, C.L.; Niederer, A.F.; Graafhuis, A.E. Effects of ageing and cooking on the tenderness of beef muscle. J. Sci. Food Agric. 1976, 27, 251–256. [Google Scholar] [CrossRef]
- Bouton, P.E.; Harris, P.V.; Shorthose, W.R.; Ratcliff, D. Changes in the mechanical properties of veal muscles produced by myofibrillar contraction state, cooking temperature and cooking time. J. Food Sci. 1974, 39, 869–875. [Google Scholar] [CrossRef]
- Barbera, S.; Tassone, S. Meat cooking shrinkage: Measurement of a new meat quality parameter. Meat Sci. 2006, 73, 467–474. [Google Scholar] [CrossRef]
- Cho, S.; Kang, S.M.; Seong, P.; Kang, G.; Kim, Y.; Kim, J.; Lee, S.; Kim, S. Effect of aging time on physicochemical meat quality and sensory property of hanwoo bull beef. Korean J. Food Sci. Anim. Resour. 2016, 36, 68–76. [Google Scholar] [CrossRef]
- Chinzorig, O.; Hwang, I. Mechanical texture profile of Hanwoo muscles as a function of heating temperatures. J. Anim. Sci. Technol. 2018, 60, 22. [Google Scholar] [CrossRef] [Green Version]
- Rhee, M.S.; Wheeler, T.L.; Shackelford, S.D.; Koohmaraie, M. Variation in palatability and biochemical traits within and among eleven beef muscles. J. Anim. Sci. 2004, 82, 534–550. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, S.D.; Wheeler, T.L.; Koohmaraie, M. Relationship between shear force and trained sensory panel tenderness ratings of 10 major muscles from Bos indicus and Bos taurus cattle. J. Anim. Sci. 1995, 73, 3333–3340. [Google Scholar] [CrossRef] [PubMed]
- Keith, F.K.; Miles, R.S.; Bechtel, P.J.; Carr, T.R. Chemical and sensory properties of thirteen major beef muscles. J. Food Sci. 1985, 50, 869–872. [Google Scholar] [CrossRef]
- Sullivan, G.A.; Calkins, C.R. Ranking beef muscles for Warner-Bratzler shear force and trained sensory panel ratings from published literature. J. Food Qual. 2011, 34, 195–203. [Google Scholar] [CrossRef]
- Kirchofer, K.; Calkins, C.R.; Gwartney, B. Fiber-type composition of muscles of the beef chuck and round 1. J. Anim. Sci. 2002, 80, 2872–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, M.; Hedrick, H. Profile of fiber types and related properties of five bovine muscles. J. Food Sci. 1977, 42, 513–517. [Google Scholar] [CrossRef]
- Bendall, J.R. The elastin content of various muscles of beef animals. J. Sci. Food Agric. 1967, 18, 553–558. [Google Scholar] [CrossRef]
- Burson, D.E.; Hunt, M.C. Proportion of collagen types I and III in four bovine muscles differing in tenderness. J. Food Sci. 1986, 51, 51–53. [Google Scholar] [CrossRef]
- Dashdorj, D.; Uddin, M.; Aguayo, D.; Ochirbat, C.; Lee, J.; Hwang, I. Collagen types of hanwoo beef in relation to texture properties of individual muscles. In Proceedings of the 63th International Congress of Meat Science and Technology: Nurturing Locally, Growing Globally, Cork, Ireland, 13–18 August 2017. [Google Scholar]
- Light, N.; Champion, A.E.; Voyle, C.; Bailey, A.J. The rôle of epimysial, perimysial and endomysial collagen in determining texture in six bovine muscles. Meat Sci. 1985, 13, 137–149. [Google Scholar] [CrossRef]
- Warner, R.; Dunshea, F.; Gutzke, D.; Lau, J.; Kearney, G. Factors influencing the incidence of high rigor temperature in beef carcasses in Australia. Anim. Prod. Sci. 2014, 54, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Colle, M.J.; Richard, R.P.; Killinger, K.M.; Bohlscheid, J.C.; Gray, A.R.; Loucks, W.I.; Day, R.N.; Cochran, A.S.; Nasados, J.A.; Doumit, M.E. Influence of extended aging on beef quality characteristics and sensory perception of steaks from the biceps femoris and semimembranosus. Meat Sci. 2016, 119, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Starkey, C.P.; Geesink, G.H.; Oddy, V.H.; Hopkins, D.L. Explaining the variation in lamb longissimus shear force across and within ageing periods using protein degradation, sarcomere length and collagen characteristics. Meat Sci. 2015, 105, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.P. Intramuscular connective tissue and its role in meat quality. Meat Sci. 2005, 70, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.P. Contribution of collagen and connective tissue to cooked meat toughness; some paradigms reviewed. Meat Sci. 2018. [Google Scholar] [CrossRef]
- Latorre, M.E.; Velázquez, D.E.; Purslow, P.P. The thermal shrinkage force in perimysium from different beef muscles is not affected by post-mortem ageing. Meat Sci. 2018, 135, 109–114. [Google Scholar] [CrossRef]
- Honikel, K. How to measure the water-holding capacity of meat? Recommendation of standardized methods. In Evaluation and Control of Meat Quality in Pigs; Springer: Berlin/Heidelberg, Germany, 1987; pp. 129–142. [Google Scholar]
- Locker, R.; Daines, G.J. Cooking loss in Beef. The effect of cold shortening, searing and rate of heating; time course and histology of changes during cooking. J. Sci. Food Agric. 1974, 25, 1411–1418. [Google Scholar] [CrossRef]
- Watson, R.; Polkinghorne, R.; Thompson, J.M. Development of the Meat Standards Australia (MSA) prediction model for beef palatability. Aust. J. Exp. Agric. 2008, 48, 1368–1379. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Kim, Y.H.B. Proteolytic changes of myofibrillar and small heat shock proteins in different bovine muscles during aging: Their relevance to tenderness and water-holding capacity. Meat Sci. 2020, 163, 108090. [Google Scholar] [CrossRef]
- Boakye, K.; Mittal, G.S. Changes in pH and water holding properties of Longissimus dorsi muscle during beef ageing. Meat Sci. 1993, 34, 335–349. [Google Scholar] [CrossRef]
- Quali, A. Proteolytic and physicocohemical mechanisms involved in meat texture development. Biochimie 1992, 74, 251–265. [Google Scholar] [CrossRef]
- Lewis, G.J.; Purslow, P.P.; Rice, A.E. The effect of conditioning on the strength of perimysial connective tissue dissected from cooked meat. Meat Sci. 1991, 30, 1–12. [Google Scholar] [CrossRef]
- Stabursvik, E.; Martens, H. Thermal denaturation of proteins in post rigor muscle tissue as studied by differential scanning calorimetry. J. Sci. Food Agric. 1980, 31, 1034–1042. [Google Scholar] [CrossRef]
- Findlay, C.J.; Stanley, D.W. Differential scanning calorimetry of beef muscle: Influence of sarcomere length. J. Food Sci. 1984, 49, 1529–1531. [Google Scholar] [CrossRef]
- Pospiech, E.; Greaser, M.L.; Mikolajczak, B.; Chiang, W.; Krzywdzińska, M. Thermal properties of titin from porcine and bovine muscles. Meat Sci. 2002, 62, 187–192. [Google Scholar] [CrossRef]
- Lusby, M.L.; Ridpath, J.F.; Parrish, F.C.; Robson, R.M. Effect of postmortem storage on degradation of the myofibrillar protein titin in bovine longissimus muscle. J. Food Sci. 1983, 48, 1787–1790. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Parrish, F.C.; Robson, R.M. Effects of postmortem aging time, animal age, and sex on degradation of titin and nebulin in bovine longissimus muscle. J. Anim. Sci. 1995, 73, 1064–1073. [Google Scholar] [CrossRef] [Green Version]
- Bouton, P.E.; Harris, P.V.; Shorthose, W.R. Changes in shear parameters of meat associated with structural changes produced by ageing, cooking and myofibrillar contraction. J. Food Sci. 1975, 40, 1122–1126. [Google Scholar] [CrossRef]
- Møller, A.J. Analysis of Warner-Bratzler shear pattern with regard to myofibrillar and connective tissue components of tenderness. Meat Sci. 1981, 5, 247–260. [Google Scholar] [CrossRef]
- Hamm, R. Heating of muscle systems. In The Physiology and Biochemistry of Muscle as Food; Briskey, E.J., Cassens, R.G., Trautman, J.C., Eds.; The University of Wisconsin Press: Madison, WI, USA, 1966; pp. 363–385. [Google Scholar]
- Martens, H.; Stabursvik, E.; Martens, M. Texture and colour changes in meat during cooking related to thermal denaturation of muscle proteins. J. Texture Stud. 1982, 13, 291–309. [Google Scholar] [CrossRef]
- Ishiwatari, N.; Fukuoka, M.; Sakai, N. Effect of protein denaturation degree on texture and water state of cooked meat. J. Food Eng. 2013, 117, 361–369. [Google Scholar] [CrossRef]
- Dominguez-Hernandez, E.; Salaseviciene, A.; Ertbjerg, P. Low-temperature long-time cooking of meat: Eating quality and underlying mechanisms. Meat Sci. 2018, 143, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, M.; Purslow, P.P.; Larsen, L.M. The effect of cooking temperature on mechanical properties of whole meat, single muscle fibres and perimysial connective tissue. Meat Sci. 2000, 55, 301–307. [Google Scholar] [CrossRef]
- Aaslyng, M.; Bejerholm, C.; Ertbjerg, P.; Bertram, H.C.; Andersen, H.J. Cooking loss and juiciness of pork in relation to raw meat quality and cooking procedure. Food Qual. Prefer. 2003, 14, 277–288. [Google Scholar] [CrossRef]
- Kim, G.-D.; Jeong, J.-Y.; Jung, E.-Y.; Yang, H.-S.; Lim, H.-T.; Joo, S.-T. The influence of fiber size distribution of type IIB on carcass traits and meat quality in pigs. Meat Sci. 2013, 94, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.N.; Canto, A.C.V.C.S.; Rentfrow, G.; Suman, S.P. Muscle-specific effect of aging on beef tenderness. LWT 2019, 100, 250–252. [Google Scholar] [CrossRef]
- Shanks, B.; Wulf, D.; Maddock, R. The effect of freezing on Warner-Bratzler shear force values of beef longissimus steaks across several postmortem aging periods 1. J. Anim. Sci. 2002, 80, 2122–2125. [Google Scholar] [CrossRef] [Green Version]
- Lagerstedt, Å.; Enfält, L.; Johansson, L.; Lundström, K. Effect of freezing on sensory quality, shear force and water loss in beef m. longissimus dorsi. Meat Sci. 2008, 80, 457–461. [Google Scholar] [CrossRef]
- Kristensen, L.; Purslow, P.P. The effect of ageing on the water-holding capacity of pork: Role of cytoskeletal proteins. Meat Sci. 2001, 58, 17–23. [Google Scholar] [CrossRef]
- Dube, G.; Bramblett, V.D.; Judge, M.D.; Harrington, R.B. Physical properties and sulfhydryl content of bovine muscles. J. Food Sci. 1972, 37, 23–26. [Google Scholar] [CrossRef]
- Lepetit, J.; Grajales, A.; Favier, R. Modelling the effect of sarcomere length on collagen thermal shortening in cooked meat: Consequence on meat toughness. Meat Sci. 2000, 54, 239–250. [Google Scholar] [CrossRef]
- Vaskoska, R.; Ha, M.; Ong, L.; Kearney, G.; White, J.; Gras, S.; Warner, R. Ageing and cathepsin inhibition affect the shrinkage of fibre fragments of bovine semitendinosus, biceps femoris and psoas major during heating. 2020. under review. [Google Scholar]
- Locker, R.H.; Daines, G.J. Transverse anisotropy in beef muscle. J. Sci. Food Agric. 1976, 27, 186–192. [Google Scholar] [CrossRef]
- Mohr, V.; Bendall, J.R. Constitution and physical chemical properties of intramuscular connective tissue. Nature 1969, 223, 404–405. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.P.; Trotter, J.A. The morphology and mechanical properties of endomysium in series-fibred muscles: Variations with muscle length. J. Muscle Res. Cell Motil. 1994, 15, 299–308. [Google Scholar] [CrossRef]
- Purslow, P.P. Strain-induced reorientation of an intramuscular connective tissue network: Implications for passive muscle elasticity. J. Biomech. 1989, 22, 21–31. [Google Scholar] [CrossRef]
- Hiner, E.L.; Hankins, O.G.; Sloane, H.S.; Fellers, C.R.; Anderson, E.E. Fiber diameter in relation to tenderness of beef muscle. J. Food Sci. 1953, 18, 364–376. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Muscle | Ageing | SED (Ageing) | p-Values | |||
|---|---|---|---|---|---|---|
| 1 Day | 14 Days | Muscle | Ageing | Muscle × Ageing | ||
| semitendinosus | 5.41 | 5.49 | 0.026 | >0.05 | <0.01 | >0.05 |
| biceps femoris | 5.36 | 5.50 | ||||
| psoas major | 5.48 | 5.54 | ||||
| Characteristic | Muscle | Mean | SED (Muscle) | p-Value |
|---|---|---|---|---|
| Sarcomere length (µm) | semitendinosus | 2.04 | 0.059 | <0.001 |
| biceps femoris | 1.89 | |||
| psoas major | 3.46 | |||
| Total collagen content (µg/mg freeze dried tissue) | semitendinosus | 39.25 | 3.969 | <0.001 |
| biceps femoris | 47.40 | |||
| psoas major | 14.29 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaskoska, R.; Ha, M.; Naqvi, Z.B.; White, J.D.; Warner, R.D. Muscle, Ageing and Temperature Influence the Changes in Texture, Cooking Loss and Shrinkage of Cooked Beef. Foods 2020, 9, 1289. https://doi.org/10.3390/foods9091289
Vaskoska R, Ha M, Naqvi ZB, White JD, Warner RD. Muscle, Ageing and Temperature Influence the Changes in Texture, Cooking Loss and Shrinkage of Cooked Beef. Foods. 2020; 9(9):1289. https://doi.org/10.3390/foods9091289
Chicago/Turabian StyleVaskoska, Rozita, Minh Ha, Zahra Batool Naqvi, Jason David White, and Robyn Dorothy Warner. 2020. "Muscle, Ageing and Temperature Influence the Changes in Texture, Cooking Loss and Shrinkage of Cooked Beef" Foods 9, no. 9: 1289. https://doi.org/10.3390/foods9091289
APA StyleVaskoska, R., Ha, M., Naqvi, Z. B., White, J. D., & Warner, R. D. (2020). Muscle, Ageing and Temperature Influence the Changes in Texture, Cooking Loss and Shrinkage of Cooked Beef. Foods, 9(9), 1289. https://doi.org/10.3390/foods9091289