Bioprospecting Cultivated Tropical Green Algae, Caulerpa racemosa (Forsskal) J. Agardh: A Perspective on Nutritional Properties, Antioxidative Capacity and Anti-Diabetic Potential

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection, Identification and Processing of Algal Material
2.2. Proximate Analysis
2.3. Fatty Acid Determination
2.4. Amino Acid Analysis
2.5. Mineral and Heavy Metal Analysis
2.6. Antioxidative Properties
2.6.1. Total Phenolic Content (Tpc) and Flavonoid Content (Tfc)
2.6.2. Estimation of Total Antioxidant Activity
2.6.3. Estimation of Ferric Reducing Antioxidant Power (Frap)
2.6.4. Estimation of Hydrogen Peroxide (H 2O 2)
2.7. In Vivo Anti-Diabetic Study
2.7.1. Experimental Animals
2.7.2. Experimental Design
2.7.3. Oral Glucose Tolerance Test (OGTT)
2.8. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition of C. racemosa
3.2. Fatty Acids Composition
3.3. Amino Acid Composition
3.4. Mineral Contents
3.5. Anti-Oxidative Properties
3.5.1. Total Phenol Content (TPC) and Total Flavonoid Content (TFC).
3.5.2. Antioxidant Capacity
3.6. In Vivo Anti-Diabetic Activity
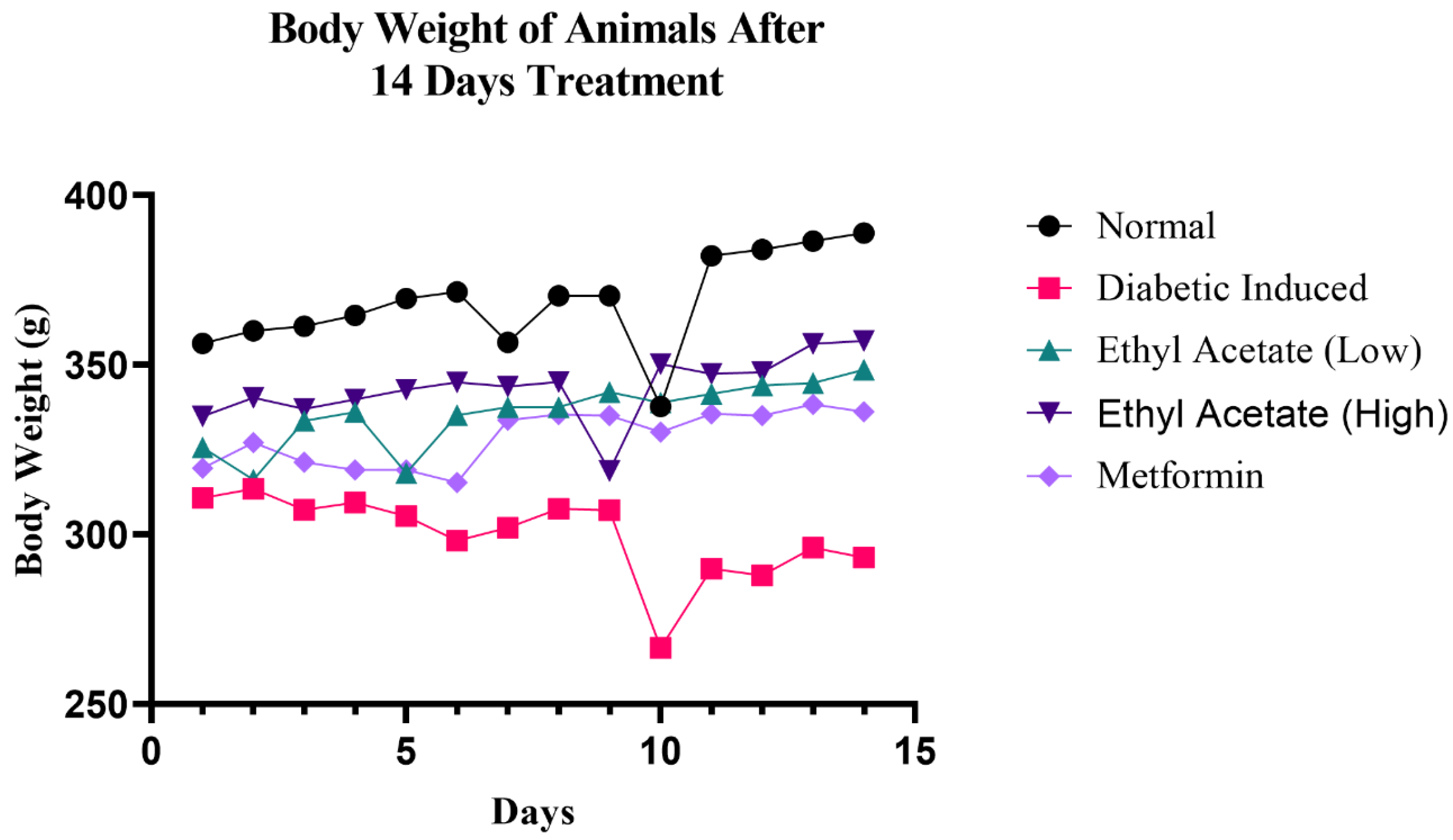
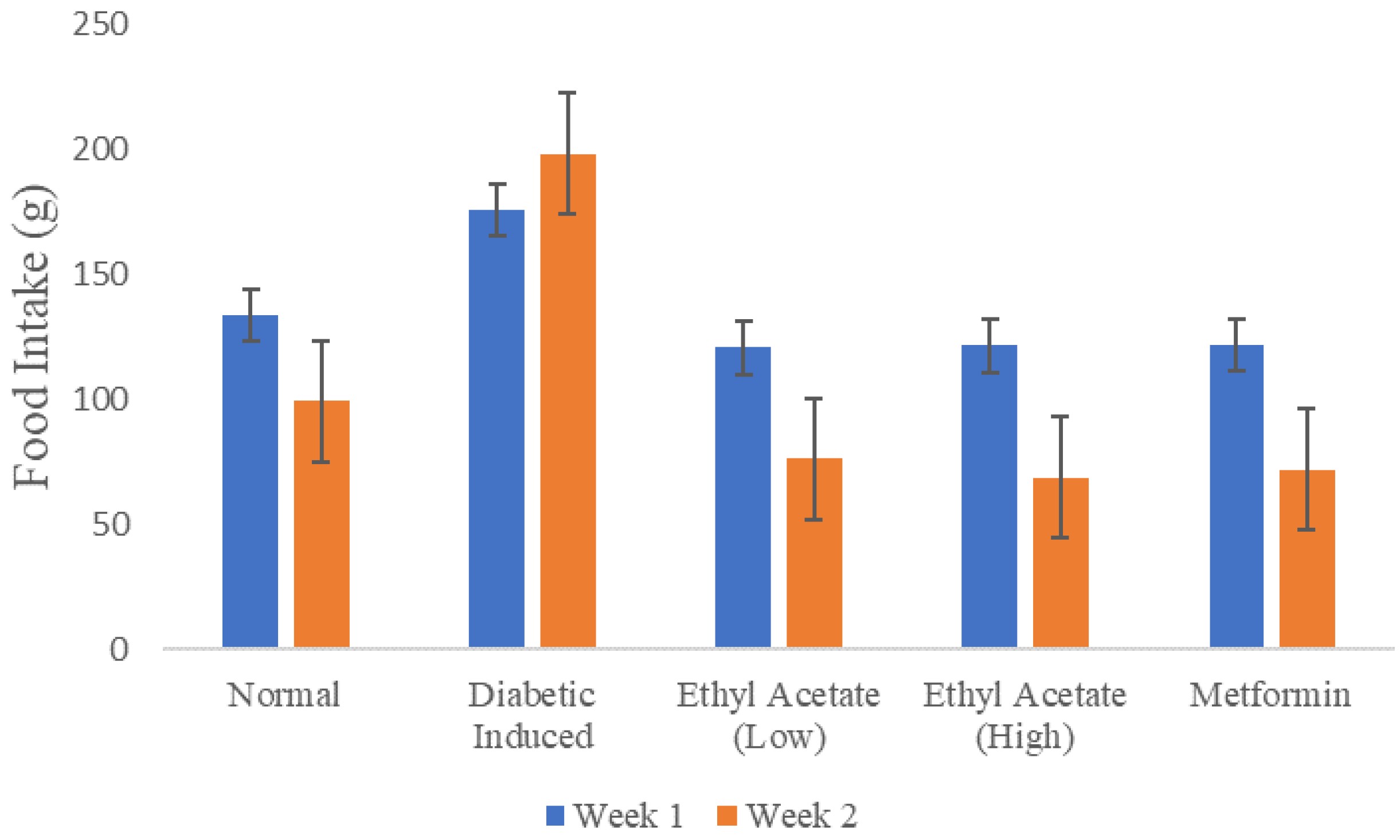
3.6.1. Effect of C. racemosa Etoac Extract on the Body Weight and Food Intake
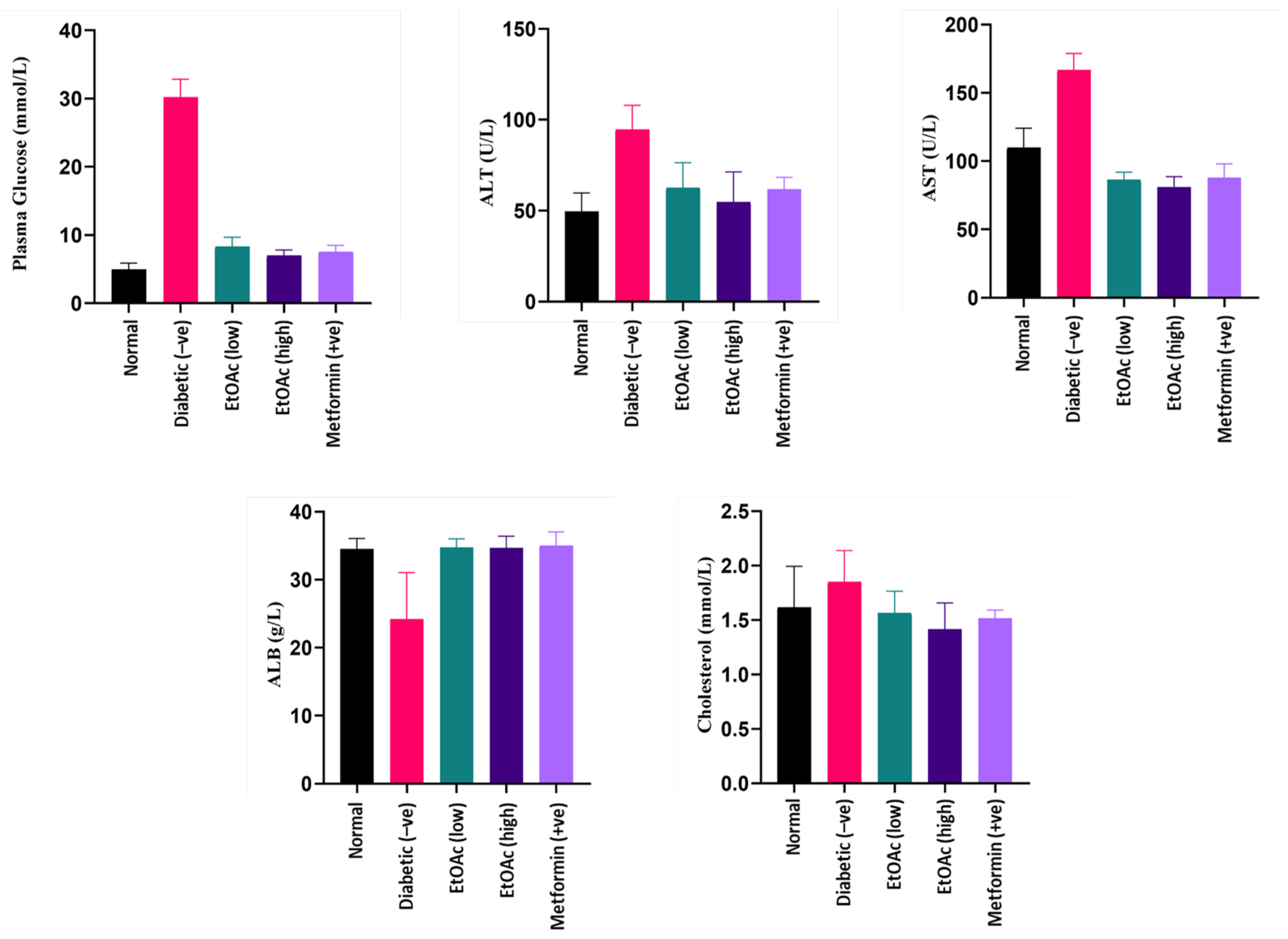
3.6.2. Effects of C. racemosa Etoac Extract on Plasma Glucose Levels
3.6.3. Effect of C. racemosa Ethyl Acetate Extract on Plasma ALT and AST Levels
3.6.4. Effects of C. racemosa Etoac Extract on Albumin Levels
3.6.5. Effects of C. racemosa Etoac Extract on Cholesterol Levels
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhuyar, P.; Rahim, M.H.; Sundararaju, S.; Maniam, G.P.; Govindan, N. Antioxidant and antibacterial activity of red seaweed Kappaphycus alvarezii against pathogenic bacteria. Glob. J. Environ. Sci. Manag. 2020, 6, 47–58. [Google Scholar]
- Okuda, K. Coastal environment and seaweed-bed ecology in Japan. Kuroshio Sci. 2008, 2, 15–20. [Google Scholar]
- Nicoletti, R.; Trincone, A. Bioactive compounds produced by strains of Penicillium and Talaromyces of marine origin. Mar. Drugs 2016, 14, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, K.; Fitzgerald, G.; Stanton, C.; Ross, R. Looking beyond the terrestrial: The potential of seaweed derived bioactives to treat non-communicable diseases. Mar. Drugs 2016, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.T.; Ueng, J.P.; Tsai, G.J. Proximate composition, total phenolic content, and antioxidant activity of seagrape (Caulerpa lentillifera). J. Food Sci. 2011, 76, C950–C958. [Google Scholar] [CrossRef]
- Nagaraj, S.R.; Osborne, J.W. Bioactive compounds from Caulerpa racemosa as a potent larvicidal and antibacterial agent. Front. Biol. 2014, 9, 300–305. [Google Scholar] [CrossRef]
- Tanna, B.; Mishra, A. Metabolites unravel nutraceutical potential of edible seaweeds: An emerging source of functional food. Compr. Rev. Food Sci. Food Saf. 2018, 17, 161–1624. [Google Scholar] [CrossRef] [Green Version]
- Largo, D.B.; Diola, A.G.; Marababol, M.S. Development of an integrated multi-trophic aquaculture (IMTA) system for tropical marine species in southern cebu, Central Philippines. Aquacult. Rep. 2016, 3, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Phang, S.M.; Critchley, A.T.; Ang, P.O. Advances in seaweed cultivation and utilisation in Asia. In Proceedings from Asian Fisheries Forum 2004: Penang, Malaysia; Phang, S.M., Critchley, A.T., Ang, P.O., Jr., Eds.; University of Malaya Press: Kuala Lumpur, Malaysia, 2006; p. 177. [Google Scholar]
- Novaczek, I. A Guide to the Common Edible and Medicinal Sea Plants of the Pacific Islands; Community Fisheries Training Pacific Series; University of the South Pacific: Suva, Fiji, 2001; p. 3. [Google Scholar]
- Kumar, M.; Kumari, P.; Trivedi, N.; Shukla, M.K.; Gupta, V.; Reddy, C.; Jha, B. Minerals, PUFAs and antioxidant properties of some tropical seaweeds from Saurashtra coast of India. J. Appl. Phycol. 2011, 23, 797–810. [Google Scholar] [CrossRef]
- Nagappan, T.; Vairappan, C.S. Nutritional and bioactive properties of three edible species of green algae, genus Caulerpa (Caulerpaceae). J. Appl. Phycol. 2014, 26, 1019–1027. [Google Scholar] [CrossRef]
- Kumar, M.; Gupta, V.; Kumari, P.; Reddy, C.R.K.; Jha, B. Assessment of nutrient composition and antioxidant potential of Caulerpaceae seaweeds. J. Food Compos. Anal. 2011, 24, 270–278. [Google Scholar] [CrossRef]
- Tanna, B.; Choudhary, B.; Mishra, A. Metabolite profiling, antioxidant, scavenging and anti-proliferative activities of selected tropical green seaweeds reveal the nutraceutical potential of Caulerpa spp. Algal Res. 2018, 36, 96–105. [Google Scholar] [CrossRef]
- Yap, W.-F.; Tay, V.; Tan, S.-H.; Yow, Y.-Y.; Chew, J. Decoding Antioxidant and Antibacterial Potentials of Malaysian Green Seaweeds: Caulerpa racemosa and Caulerpa lentillifera. Antibiotics 2019, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Paul, N.A.; Neveux, N.; Magnusson, M.; De Nys, R. Comparative production and nutritional value of “sea grapes”—The tropical green seaweeds Caulerpa lentillifera and Caulerpa racemosa. J. Appl. Phycol. 2014, 26, 1833–1844. [Google Scholar] [CrossRef]
- Smyth, K.; Elliott, M. Effects of changing salinity on the ecology of the marine environment. In Stressors in the Marine Environment: Physiological and Ecological Responses; Societal Implications; Solan, M., Whiteley, N., Eds.; Oxford University Press: Selangor, Malaysia, 2016; pp. 161–174. [Google Scholar]
- Verlaque, M.; Boudouresque, C.F.; Meinesz, A.; Gravez, V. The Caulerpa racemosa complex (Caulerpales, Ulvophyceae) in the Mediterranean sea. Bot. Mar. 2000, 43, 49–68. [Google Scholar] [CrossRef]
- Belleza, D.F.C.; Liao, L.M. Taxonomic inventory of the marine green algal genus Caulerpa (Chlorophyta, Bryopsidales) at the University of San Carlos (Cebu) herbarium. Philipp. Sci. 2007, 44, 71–104. [Google Scholar] [CrossRef]
- Zubia, M.; Draisma, S.G.; Morrissey, K.L.; Varela-Álvarez, E.; De Clerck, O. Concise review of the genus Caulerpa JV Lamouroux. J. Appl. Phycol. 2019, 32, 23–39. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Nutritional quality of some wild and cultivated seaweeds: Nutrient composition, total phenolic content and in vitro digestibility. J. Appl. Phycol. 2016, 28, 3575–3585. [Google Scholar] [CrossRef]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- McDermid, K.J.; Stuercke, B. Nutritional composition of edible Hawaiian seaweeds. J. Appl. Phycol. 2003, 15, 513–524. [Google Scholar] [CrossRef]
- Aroyehun, A.Q.; Palaniveloo, K.; Ghazali, F.; Rizman-Idid, M.; Razak, S.A. Effects of Seasonal Variability on the Physicochemical, Biochemical, and Nutritional Composition of Western Peninsular Malaysia Gracilaria manilaensis. Molecules 2019, 24, 3298. [Google Scholar] [CrossRef] [PubMed]
- Concha-Herrera, V.; Lerma-García, M.J.; Herrero-Martinez, J.M.; Simo-Alfonso, E.F. Classification of vegetable oils according to their botanical origin using amino acid profiles established by high performance liquid chromatography with UV–vis detection: A first approach. Food Chem. 2010, 120, 1149–1154. [Google Scholar] [CrossRef]
- Pan, Y.; Wernberg, T.; de Bettignies, T.; Holmer, M.; Li, K.; Wu, J.; Lin, F.; Yu, Y.; Xu, J.; Zhou, C. Screening of seaweeds in the East China Sea as potential bio-monitors of heavy metals. Environ. Sci. Pollut. Res. 2018, 25, 16640–16651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, K.; Joseph, D.; Praveen, N.K. Antioxidant activities and phenolic contents of three red seaweeds (Division: Rhodophyta) harvested from the Gulf of Mannar of Peninsular India. J. Food Sci. Technol. 2015, 52, 1924–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akter, M.; Afrin, S.; Sakib, M.; Biswas, R.; Billah, M.M.; Zohora, U.S. Investigation of Antibacterial, Cytotoxic and antioxidant properties of the mangrove plant Xylocarpus mekongensis. Adv. Biosci. Biotechnol. 2016, 7, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Saeed, N.; Khan, M.R.; Shabbir, M. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts Torilis leptophylla L. BMC Complement. Altern. Med. 2012, 12, 221. [Google Scholar] [CrossRef] [Green Version]
- Suthagar, E.; Soudamani, S.; Yuvaraj, S.; Ismail Khan, A.; Aruldhas, M.M.; Balasubramanian, K. Effects of streptozotocin (STZ)-induced diabetes and insulin replacement on rat ventral prostate. Biomed. Pharmacother. 2009, 63, 43–50. [Google Scholar] [CrossRef]
- Owen, M.R.; Doran, E.; Halestrap, A.P. Evidence that metformin exerts its anti-diabetic effects through inhibition of complex 1 of the mitochondrial respiratory chain. Biochem. J. 2000, 348, 607–614. [Google Scholar] [CrossRef]
- Eddouks, M.; Chattopadhyay, D.; Zeggwagh, N.A. Animal models as tools to investigate antidiabetic and anti-inflammatory plants. Evid.-Based Complement. Altern. Med. 2012, 2012, 142087. [Google Scholar] [CrossRef] [PubMed]
- Al-Awadi, F.M.; Khattar, M.A.; Gumaa, K.A. On the mechanism of the hypoglycaemic effect of a plant extract. Diabetologia 1985, 28, 432–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, H.; Fu, M.; Yan, R.; He, B.; Li, M.; Liu, Q.; Cai, Y.; Zhang, X.; Huang, R. Chemical composition and immunostimulatory properties of green alga Caulerpa racemosa var peltata. Food Agric. Immunol. 2019, 30, 937–954. [Google Scholar] [CrossRef] [Green Version]
- Renaud, S.M.; Luong-Van, J.T. Seasonal variation in the chemical composition of tropical Australian marine macroalgae. In Proceedings from Eighteenth International Seaweed Symposium; Critchley, A.T., Brodie, J.A., Onsoyen, E., Anderson, R.J., Eds.; Springer Nature: Cham, Switzerland, 2006; pp. 155–161. [Google Scholar]
- De Gaillande, C.; Payri, C.; Remoissenet, G.; Zubia, M. Caulerpa consumption, nutritional value and farming in the Indo-Pacific region. J. Appl. Phycol. 2017, 29, 2249–2266. [Google Scholar] [CrossRef]
- Bocanegra, A.; Bastida, S.; Benedi, J.; Rodenas, S.; Sanchez-Muniz, F.J. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J. Med. Food. 2009, 12, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Setthamongkol, P.; Tunkijjanukij, S.; Satapornvanit, K.; Salaenoi, J. Growth and nutrients analysis in marine macroalgae. Agric. Nat. Resour. 2015, 49, 211–218. [Google Scholar]
- Bhuiyan, M.; Qureshi, S.; Mustafa Kamal, A.; AftabUddin, S.; Siddique, A. Proximate chemical composition of sea grapes Caulerpa racemosa (J. Agardh, 1873) collected from a sub-tropical coast. Virol. Mycol. 2016, 5, 2161–0517. [Google Scholar]
- Da Silva, V.M.; Silva, L.A.; de Andrade, J.B.; Veloso, M.C.; Santos, G.V. Determination of moisture content and water activity in algae and fish by thermoanalytical techniques. Quim. Nova 2008, 31, 901–905. [Google Scholar] [CrossRef] [Green Version]
- Rameshkumar, S.; Ramakritinan, C.; Yokeshbabu, M. Proximate composition of some selected seaweeds from Palk bay and Gulf of Mannar, Tamilnadu, India. Asian J. Biomed. Pharm. Sci. 2013, 3, 1. [Google Scholar]
- Khairy, H.M.; El-Shafay, S.M. Seasonal variations in the biochemical composition of some common seaweed species from the coast of Abu Qir Bay, Alexandria, Egypt. Oceanologia 2013, 55, 435–452. [Google Scholar] [CrossRef] [Green Version]
- Salleh, A.; Wakid, S.A. Nutritional Composition of Macroalgae in Tanjung Tuan, Port Dickson, Malaysia. Malays. J. Sci. 2008, 27, 19–26. [Google Scholar]
- Nomura, M.; Kamogawa, H.; Susanto, E.; Kawagoe, C.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Seasonal variations of total lipids, fatty acid composition, and fucoxanthin contents of Sargassum horneri (Turner) and Cystoseira hakodatensis (Yendo) from the northern seashore of Japan. J. Appl. Phycol. 2013, 25, 1159–1169. [Google Scholar] [CrossRef]
- Fariman, G.A.; Shastan, S.J.; Zahedi, M.M. Seasonal variation of total lipid, fatty acids, fucoxanthin content, and antioxidant properties of two tropical brown algae (Nizamuddinia zanardinii and Cystoseira indica) from Iran. J. Appl. Phycol. 2016, 28, 1323–1331. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Resek, E.; Engelen, A.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, P.; Bijo, A.; Mantri, V.A.; Reddy, C.; Jha, B. Fatty acid profiling of tropical marine macroalgae: An analysis from chemotaxonomic and nutritional perspectives. Phytochemistry 2013, 86, 44–56. [Google Scholar] [CrossRef]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K. Nutrient content of tropical edible seaweeds, Eucheuma cottonii, Caulerpa lentillifera and Sargassum polycystum. J. Appl. Phycol. 2009, 21, 75–80. [Google Scholar] [CrossRef]
- Astorga-Espana, M.S.; Rodriguez-Galdon, B.; Rodriguez-Rodriguez, E.M.; Diaz-Romero, C. Amino acid content in seaweeds from the Magellan Straits (Chile). J. Food Compos. Anal. 2016, 53, 77–84. [Google Scholar] [CrossRef]
- Teuling, E.; Wierenga, P.A.; Schrama, J.W.; Gruppen, H. Comparison of protein extracts from various unicellular green sources. J. Agric. Food Chem. 2017, 65, 7989–8002. [Google Scholar] [CrossRef] [Green Version]
- Martinez–Hernandez, G.B.; Castillejo, N.; Carrion–Monteagudo, M.d.M.; Artes, F.; Artes-Hernandez, F. Nutritional and bioactive compounds of commercialized algae powders used as food supplements. Food Sci. Technol. Int. 2018, 24, 172–182. [Google Scholar] [CrossRef]
- Cofrades, S.; Lopez-Lopez, I.; Bravo, L.; Ruiz-Capillas, C.; Bastida, S.; Larrea, M.T.; Jimenez-Colmenero, F. Nutritional and antioxidant properties of different brown and red Spanish edible seaweeds. Food Sci. Technol. Int. 2010, 16, 361–370. [Google Scholar] [CrossRef]
- Misurcova, L.; Bunka, F.; Ambrozova, J.V.; Machu, L.; Samek, D.; Kracmar, S. Amino acid composition of algal products and its contribution to RDI. Food Chem. 2014, 151, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, S.O.; Barbarino, E.; De-Paula, J.C.; Pereira, L.O.d.S.; Marquez, U.M.L. Amino acid composition, protein content and calculation of nitrogen-to-protein conversion factors for 19 tropical seaweeds. Phycol. Res. 2002, 50, 233–241. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Patarra, R.F.; Neto, A.I.; Baptista, J. Edible Azorean macroalgae as source of rich nutrients with impact on human health. Food Chem. 2014, 164, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Rodrıguez, I.; Huerta-Diaz, M.; Choumiline, E.; Holguın-Quinones, O.; Zertuche-Gonzalez, J. Elemental concentrations in different species of seaweeds from Loreto Bay, Baja California Sur, Mexico: Implications for the geochemical control of metals in algal tissue. Environ. Pollut. 2001, 114, 145–160. [Google Scholar] [CrossRef]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.; Vasconcelos, M.W.; Roriz, M.; Rodriguez-Alcala, L.M.; Gomes, A.M.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Stipanuk, M.H.; Caudill, M.A. Biochemical, Physiological, and Molecular Aspects of Human Nutrition; Stipanuk, M.H., Caudill, M.A., Eds.; Elsevier–Health Sciences: St. Louis, MO, USA, 2013; p. 968. [Google Scholar]
- Peng, Y.; Xie, E.; Zheng, K.; Fredimoses, M.; Yang, X.; Zhou, X.; Wang, Y.; Yang, B.; Lin, X.; Liu, J.; et al. Nutritional and chemical composition and antiviral activity of cultivated seaweed Sargassum naozhouense Tseng et Lu. Mar. Drugs 2013, 11, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Arulkumar, A.; Rosemary, T.; Paramasivam, S.; Rajendran, R.B. Phytochemical composition, in vitro antioxidant, antibacterial potential and GC-MS analysis of red seaweeds (Gracilaria corticata and Gracilaria edulis) from Palk Bay, India. Biocatal Agric. Biotechnol. 2018, 15, 63–71. [Google Scholar] [CrossRef]
- Mekinic, I.G.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, M.; Perkovic, Z.P. Phenolic content of brown algae (Pheophyceae) species: Extraction, identification, and quantification. Biomolecules 2019, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Farvin, K.S.; Jacobsen, C. Phenolic compounds and antioxidant activities of selected species of seaweeds from Danish coast. Food Chem. 2013, 138, 1670–1681. [Google Scholar] [CrossRef]
- Vinayak, R.C.; Sudha, S.A.; Chatterji, A. Bio-screening of a few green seaweeds from India for their cytotoxic and antioxidant potential. J. Sci. Food Agric. 2011, 91, 2471–2476. [Google Scholar] [CrossRef]
- Marinho, G.S.; Sørensen, A.-D.M.; Safafar, H.; Pedersen, A.H.; Holdt, S.L. Antioxidant content and activity of the seaweed Saccharina latissima: A seasonal perspective. J. Appl. Phycol. 2018, 1, 1343–1354. [Google Scholar] [CrossRef] [Green Version]
- Kuda, T.; Tsunekawa, M.; Hishi, T.; Araki, Y. Antioxidant properties of driedkayamo-nori’, a brown alga Scytosiphon lomentaria (Scytosiphonales, Phaeophyceae). Food Chem. 2005, 89, 617–622. [Google Scholar] [CrossRef]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K.; Ming, C.H. Antioxidant activities and phenolics content of eight species of seaweeds from north Borneo. J. Appl. Phycol. 2008, 20, 367. [Google Scholar] [CrossRef]
- Benzie, I.F.; Szeto, Y. Total antioxidant capacity of teas by the ferric reducing/antioxidant power assay. J. Agric. Food Chem. 1999, 47, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Chew, Y.; Lim, Y.; Omar, M.; Khoo, K. Antioxidant activity of three edible seaweeds from two areas in southeast Asia. LWT-Food Sci. Technol. 2008, 41, 1067–1072. [Google Scholar] [CrossRef]
- Caccetta, R.; Al Salami, H. Screening for Antidiabetic Activities. In Metabolomics Tools for Natural Product Discovery; Roessner, U.T.E., Dias, D.A., Eds.; Springer Nature: Cham, Switzerland, 2013; pp. 207–218. [Google Scholar]
- Kavishankar, G.B.; Lakshmidevi, N. Anti-diabetic effect of a novel N-Trisaccharide isolated from Cucumis prophetarum on streptozotocin-nicotinamide induced type 2 diabetic rats. Phytomedicine 2014, 21, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Bugianesi, E.; McCullough, A.J.; Marchesini, G. Insulin resistance: A metabolic pathway to chronic liver disease. Hepatology 2005, 42, 987–1000. [Google Scholar] [CrossRef]
- Manna, P.; Das, J.; Ghosh, J.; Sil, P.C. Contribution of type 1 diabetes to rat liver dysfunction and cellular damage via activation of NOS, PARP, IκBα/NF-κB, MAPKs, and mitochondria-dependent pathways: Prophylactic role of arjunolic acid. Free Radic. Biol. Med. 2010, 48, 1465–1484. [Google Scholar] [CrossRef]
- Palsamy, P.; Sivakumar, S.; Subramanian, S. Resveratrol attenuates hyperglycemia-mediated oxidative stress, proinflammatory cytokines and protects hepatocytes ultrastructure in streptozotocin–nicotinamide-induced experimental diabetic rats. Chem. Biol. Interact. 2010, 186, 200–210. [Google Scholar] [CrossRef]
- Harris, E.H. Elevated liver function tests in type 2 diabetes. Clin. Diabetes 2005, 23, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Vergès, B. Pathophysiology of diabetic dyslipidaemia: Where are we? Diabetologia 2015, 58, 886–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Grouping | Characteristics |
|---|---|
| Group I | Normal control group |
| Group II | Diabetes-induced group |
| Group III | C. racemosa EtOAc extract treated group (100 mg kg−1 body weight) |
| Group IV | C. racemosa EtOAc extract treated group (200 mg kg−1 body weight) |
| Group V | Standard Metformin treated group |
| Component | Pond 1 | Pond 2 | Pond 3 | Average |
|---|---|---|---|---|
| Proximate Composition | ||||
| Moisture | 13.807 | 14.807 | 12.94 | 13.85 ± 0.93 |
| Ash | 28.558 | 28.097 | 28.097 | 28.25 ± 0.27 |
| Crude lipid | 4.092 | 4.562 | 3.949 | 4.20 ± 0.32 |
| Protein | 20.15 | 20.43 | 20.24 | 20.27 ± 0.14 |
| Carbohydrate | 33.393 | 32.104 | 34.774 | 33.42 ± 1.34 |
| Caloric Value (cal g−1) | 2547.45 | 2549.9 | 2536.66 | 2544.67 ± 7.04 |
| Vitamin B2 (mg kg−1) | 2.00 | 2.00 | 2.10 | 2.03 ± 0.06 |
| Fatty Acids Profile | Pond 1 | Pond 2 | Pond 3 | Average |
|---|---|---|---|---|
| SFA | ||||
| Lauric acids (C12:0) | 2.50 | 2.40 | 2.50 | 2.47 ± 0.06 |
| Myristic acid (C14:0) | 4.40 | 4.30 | 4.30 | 4.33 ± 0.06 |
| Palmitic acid (C16:0) | 63.50 | 63.10 | 63.20 | 63.27 ± 0.21 |
| Stearic acid (C18: 0) | 4.70 | 4.80 | 4.70 | 4.73 ± 0.06 |
| Behenic acid (C22:0) | 1.40 | 1.40 | 1.40 | 1.40 ± 0.00 |
| Lignoseric acid DHA (C24:0) | 4.30 | 4.50 | 4.30 | 4.37 ± 0.12 |
| Total | 80.57 ± 0.12 | |||
| MUFA | ||||
| Palmitoleic acid (C16:1) | 3.90 | 3.80 | 3.90 | 3.87 ± 0.06 |
| Oleic acid(C18:1) | 5.70 | 5.80 | 5.90 | 5.80 ± 0.10 |
| Erucic acid (C22:1) | 1.10 | 1.20 | 1.10 | 1.13 ± 0.06 |
| Nervonic acid (C24:1) | 1.40 | 1.50 | 1.40 | 1.43 ± 0.06 |
| Total | 12.23 ± 0.07 | |||
| PUFA | ||||
| -Linolenic acid (C18:3) | 1.80 | 1.80 | 1.90 | 1.83 ± 0.06 |
| Linoleic acid (C18:2) | 5.30 | 5.30 | 5.40 | 5.33 ± 0.06 |
| Total | 7.16 ± 0.07 | |||
| 6/3 | 2.91 ± 0.03 |
| Amino Acids | Pond 1 | Pond 2 | Pond 3 | Average |
|---|---|---|---|---|
| Essential Amino acids | ||||
| Threonine (Thr) | 1.49 | 1.39 | 1.52 | 1.42 ± 0.60 |
| Valine (Val) | 1.45 | 1.37 | 1.45 | 1.42 ± 0.49 |
| Phe + Tyr (TAA) | 1.21 | 1.29 | 1.22 | 1.24 ± 0. 43 |
| Isoleucine (Ile) | 0.83 | 0.82 | 0.81 | 0.82 ± 0.11 |
| Leucine (Leu) | 2.21 | 2.43 | 2.39 | 2.34 ± 1.16 |
| Lysine (Lys) | 0.40 | 0.43 | 0.44 | 0.42 ± 0.21 |
| Met + Cys (SAA) | ND | ND | ND | ND |
| Histidine | ND | ND | ND | ND |
| Tryptophan (Try) | ND | ND | ND | ND |
| ∑ EAAs | 7.59 | 7.73 | 7.67 | 7.68 ± 0.81 |
| Non-essential acids | ||||
| Aspartic acid (Asp) | 115.73 | 115.88 | 114.97 | 115.53 ± 4.85 |
| Glutamic acid (Glu) | 15.94 | 15.31 | 15.23 | 15.49 ± 3.87 |
| Serine (Ser) | 1.52 | 1.54 | 1.58 | 1.55 ± 0.27 |
| Glycine (Gly) | 1.07 | 1.13 | 1.21 | 1.14 ± 0.69 |
| Arginine (Arg) | 3.06 | 3.01 | 3.08 | 3.05 ± 0.38 |
| Alanine (Ala) | 0.78 | 0.79 | 0.77 | 0.78 ± 0.08 |
| Tyrosine (Tyr) | 6.51 | 6.53 | 6.51 | 6.52 ± 0.10 |
| Cysteine (Cys) | 2.54 | 2.74 | 2.57 | 2.62 ± 1.06 |
| Proline (Pro) | 5.47 | 5.34 | 5.23 | 5.35 ± 1.20 |
| Asparagine (Asn) | ND | ND | ND | ND |
| Glutamine (Gln) | ND | ND | ND | ND |
| ∑ NEAA | 152.62 | 152.27 | 151.15 | 152.02 ± 8.22 |
| ∑ AA | 160.21 | 160.00 | 158.82 | 159.69 ± 7.41 |
| ∑ EAAs/∑ NEAAs (ratio) | 0.05 |
| Minerals | Pond 1 | Pond 2 | Pond 3 | Average |
|---|---|---|---|---|
| Macro Mineral (ppm) | ||||
| Calcium (Ca) | 26.30 | 26.12 | 27.41 | 26.61 ± 0.70 |
| Magnesium (Mg) | 0.040 | 0.036 | 0.020 | 0.032 ± 0.01 |
| Potassium (K) | 0.055 | 0.054 | 0.056 | 0.055 ± 0.001 |
| Sodium (Na) | 0.078 | 0.089 | 0.076 | 0.081 ± 0.007 |
| Trace Mineral (ppm) | ||||
| Copper (Cu) | 0.008 | 0.012 | 0.010 | 0.010 ± 0.002 |
| Iron (Fe) | 22.98 | 24.38 | 23.86 | 23.74 ± 0.71 |
| Manganese (Mn) | 7.48 | 7.26 | 7.25 | 7.33 ± 0.13 |
| Selenium (Se) | 0.004 | 0.005 | 0.003 | 0.004 ± 0.001 |
| Zinc (Zn) | 0.048 | 0.024 | 0.024 | 0.032 ± 0.014 |
| Chromium (Cr) | 0.002 | 0.004 | 0.003 | 0.003 ± 0.001 |
| Cobalt (Co) | 0.002 | 0.002 | 0.002 | 0.002 ± 0.00 |
| Heavy metal (ppb ) | ||||
| Total Arsenic (As) | 3.00 | 3.00 | 3.00 | 3.00 ± 0.00 |
| Cadmium (Cd) | - | - | - | <MDL |
| Lead (Pb) | 4.52 | 4.84 | 4.65 | 4.67 ± 0.16 |
| Aluminium (Al) | 1798.33 | 1890.67 | 1967.95 | 1885.65 ± 84.92 |
| C. racemosa | Total Phenolic Content | Total Flavonoid | Extraction Yield |
|---|---|---|---|
| (mg GAE g−1) | (mg QE g−1) | (%) | |
| Pond 1 | 17.23 ± 0.55 | 58.90 ± 3.22 | 31.8 |
| Pond 2 | 17.52 ± 1.78 | 59.52 ± 1.58 | 25.7 |
| Pond 3 | 18.89 ± 0.82 | 59.87 ± 2.55 | 22.3 |
| Average | 17.88 ± 0.78 | 59.43 ± 2.45 | 26.6 |
| Correlation (R2) | |||
| Radical Scavenging Value | Phenols | Flavonoids | |
| TAC | |||
| Pond 1 | 0.26 ± 1.45 | ||
| Pond 2 | 0.98 ± 2.13 | 0.8169 | 0.3797 |
| Pond 3 | 0.48 ± 1.94 | ||
| Average | 0.31 ± 1.84 | ||
| FRAP | |||
| Pond 1 | 6.05 ± 1.98 | ||
| Pond 2 | 5.79 ± 2.55 | 0.4372 | 0.991 |
| Pond 3 | 6.88 ± 4.56 | ||
| Average | 6.24 ± 3.03 | ||
| H2O2 | |||
| Pond 1 | 80.24 ± 0.23 | ||
| Pond 2 | 80.63 ± 0.84 | 0.1425 | 0.2131 |
| Pond 3 | 80.78 ± 0.58 | ||
| Average | 80.55 ± 0.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qudus B Aroyehun, A.; Abdul Razak, S.; Palaniveloo, K.; Nagappan, T.; Suraiza Nabila Rahmah, N.; Wee Jin, G.; Chellappan, D.K.; Chellian, J.; Kunnath, A.P. Bioprospecting Cultivated Tropical Green Algae, Caulerpa racemosa (Forsskal) J. Agardh: A Perspective on Nutritional Properties, Antioxidative Capacity and Anti-Diabetic Potential. Foods 2020, 9, 1313. https://doi.org/10.3390/foods9091313
Qudus B Aroyehun A, Abdul Razak S, Palaniveloo K, Nagappan T, Suraiza Nabila Rahmah N, Wee Jin G, Chellappan DK, Chellian J, Kunnath AP. Bioprospecting Cultivated Tropical Green Algae, Caulerpa racemosa (Forsskal) J. Agardh: A Perspective on Nutritional Properties, Antioxidative Capacity and Anti-Diabetic Potential. Foods. 2020; 9(9):1313. https://doi.org/10.3390/foods9091313
Chicago/Turabian StyleQudus B Aroyehun, Abdul, Shariza Abdul Razak, Kishneth Palaniveloo, Thilahgavani Nagappan, Nur Suraiza Nabila Rahmah, Gan Wee Jin, Dinesh Kumar Chellappan, Jestin Chellian, and Anil Philip Kunnath. 2020. "Bioprospecting Cultivated Tropical Green Algae, Caulerpa racemosa (Forsskal) J. Agardh: A Perspective on Nutritional Properties, Antioxidative Capacity and Anti-Diabetic Potential" Foods 9, no. 9: 1313. https://doi.org/10.3390/foods9091313
APA StyleQudus B Aroyehun, A., Abdul Razak, S., Palaniveloo, K., Nagappan, T., Suraiza Nabila Rahmah, N., Wee Jin, G., Chellappan, D. K., Chellian, J., & Kunnath, A. P. (2020). Bioprospecting Cultivated Tropical Green Algae, Caulerpa racemosa (Forsskal) J. Agardh: A Perspective on Nutritional Properties, Antioxidative Capacity and Anti-Diabetic Potential. Foods, 9(9), 1313. https://doi.org/10.3390/foods9091313