
α-Lipoic Acid Protects against Cyclosporine A-Induced Hepatic Toxicity in Rats: Effect on Oxidative Stress, Inflammation, and Apoptosis

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Experimental Design
2.4. Sampling
2.5. Biochemical Investigations
2.6. Real Time Polymerase Chain Reaction
2.7. Histopathological Examination
2.8. Immunohistochemical Studies
2.9. Statistical Analysis
3. Results
3.1. Effect of α-LA on CsA-Induced Changes in Liver Function
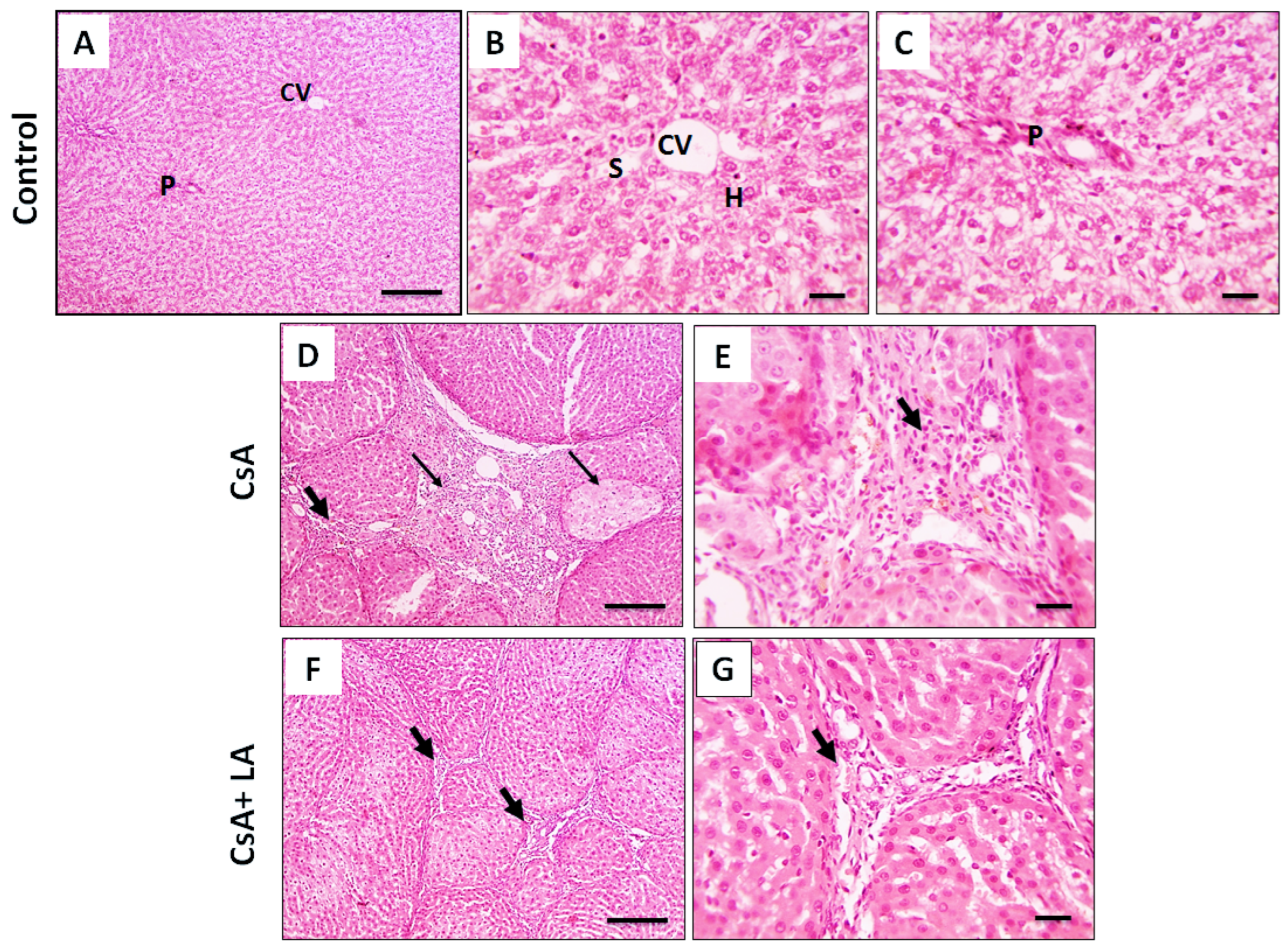
3.2. Effect of α-LA on CsA-Induced Hepatic Cellular Injury and Fibrosis
3.3. Effect of α-LA on CsA-Induced Oxidative Stress in Rat Serum and Liver Tissues
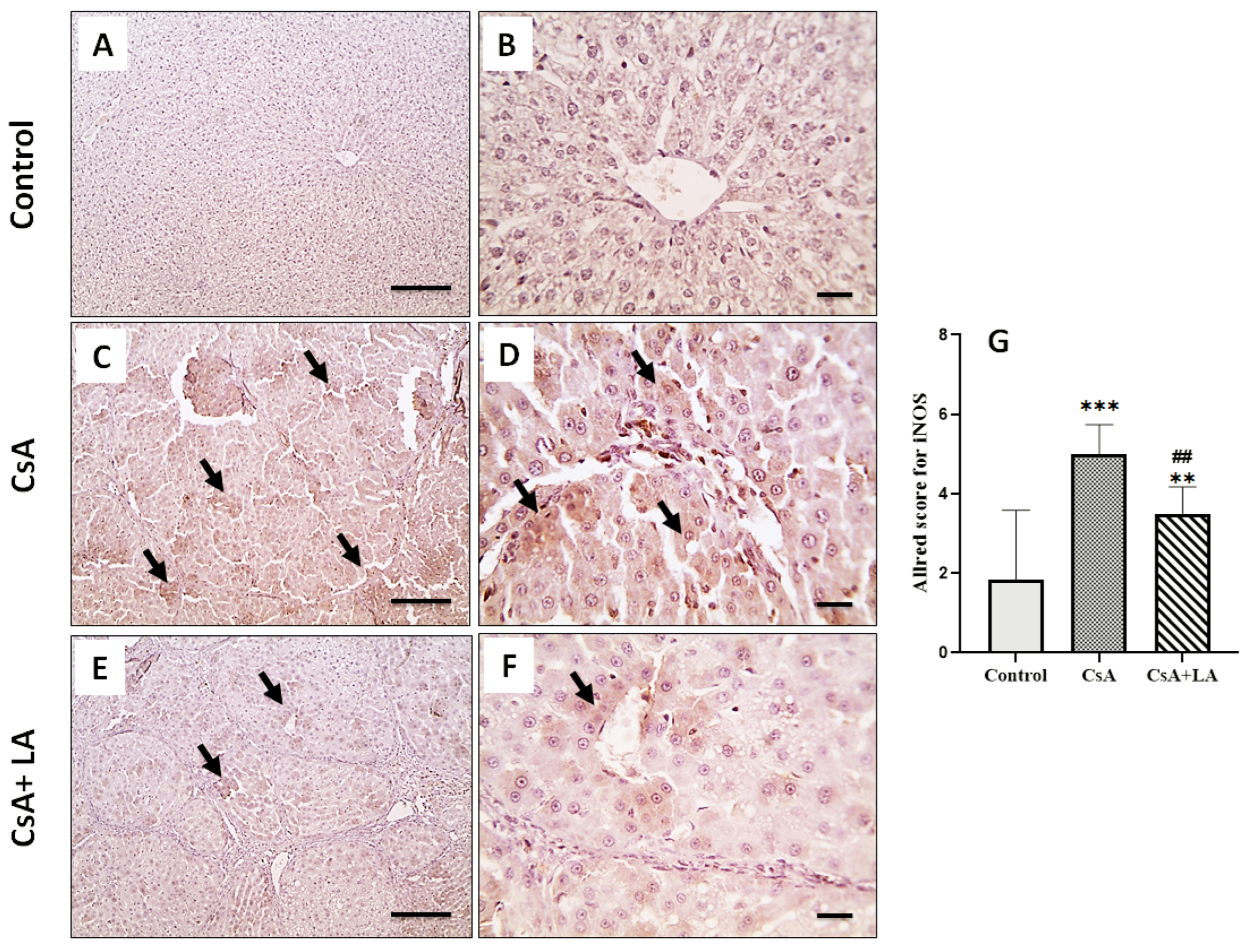
3.4. Effect of α-LA on CsA-Induced Inflammation in Rat Liver Tissues
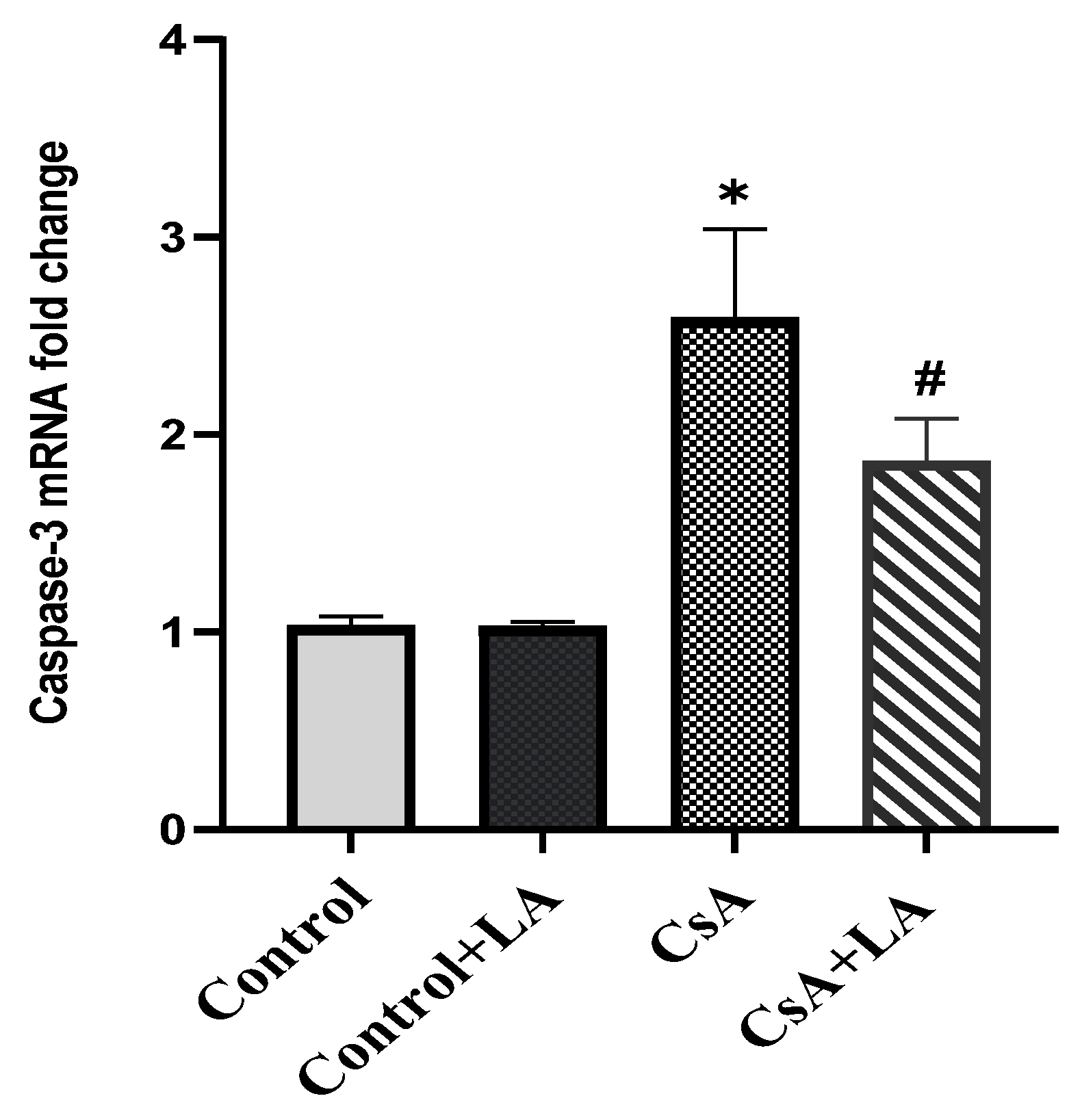
3.5. Effect of α-LA on CsA-Induced Apoptosis in Rat Liver Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vangaveti, S.; Das, P.; Kumar, V.L. Metformin and Silymarin Afford Protection in Cyclosporine A Induced Hepatorenal Toxicity in Rat by Modulating Redox Status and Inflammation. J. Biochem. Mol. Toxicol. 2021, 35, e22614. [Google Scholar] [CrossRef] [PubMed]
- Degasperi, G.R.; Castilho, R.F.; Vercesi, A.E. High Susceptibility of Activated Lymphocytes to Oxidative Stress-Induced Cell Death. Acad. Bras. Cienc. 2008, 80, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmoslemany, A.M.; El-Magd, M.A.; Ghamry, H.I.; Alshahrani, M.Y.; Zidan, N.S.; Zedan, A.M.G. Avocado Seeds Relieve Oxidative Stress-Dependent Nephrotoxicity but Enhance Immunosuppression Induced by Cyclosporine in Rats. Antioxidants 2021, 10, 1194. [Google Scholar] [CrossRef]
- Mahmoud, N.M.; Elshazly, S.M.; Rezq, S. Geraniol Protects against Cyclosporine A-Induced Renal Injury in Rats: Role of Wnt/β-Catenin and PPARγ Signaling Pathways. Life Sci. 2022, 291, 120259. [Google Scholar] [CrossRef]
- Pibiri, M.; Sulas, P.; Camboni, T.; Leoni, V.P.; Simbula, G. α-Lipoic Acid Induces Endoplasmic Reticulum Stress-Mediated Apoptosis in Hepatoma Cells. Sci. Rep. 2020, 10, 7139. [Google Scholar] [CrossRef] [PubMed]
- Hossain, K.F.B.; Akter, M.; Rahman, M.M.; Sikder, M.T.; Rahaman, M.S.; Yamasaki, S.; Kimura, G.; Tomihara, T.; Kurasaki, M.; Saito, T. Amelioration of Metal-Induced Cellular Stress by α-Lipoic Acid and Dihydrolipoic Acid through Antioxidative Effects in PC12 Cells and Caco-2 Cells. Int. J. Environ. Res. Public Health 2021, 18, 2126. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Ghibu, S.; Muresan, A.; Vergely, C. Alpha-Lipoic Acid: Molecular Mechanisms and Therapeutic Potential in Diabetes. Can. J. Physiol. Pharmacol. 2015, 93, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Joksimovic, S.L.; Lamborn, N.; Jevtovic-Todorovic, V.; Todorovic, S.M. Alpha Lipoic Acid Attenuates Evoked and Spontaneous Pain Following Surgical Skin Incision in Rats. Channels 2021, 15, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Deore, M.S.; Keerthana, S.; Naqvi, S.; Kumar, A.; Flora, S.J.S. Alpha-Lipoic Acid Protects Co-Exposure to Lead and Zinc Oxide Nanoparticles Induced Neuro, Immuno and Male Reproductive Toxicity in Rats. Front. Pharmacol. 2021, 12, 626238. [Google Scholar] [CrossRef]
- Yang, Y.; Li, W.; Liu, Y.; Sun, Y.; Li, Y.; Yao, Q.; Li, J.; Zhang, Q.; Gao, Y.; Gao, L.; et al. Alpha-Lipoic Acid Improves High-Fat Diet-Induced Hepatic Steatosis by Modulating the Transcription Factors SREBP-1, FoxO1 and Nrf2 via the SIRT1/LKB1/AMPK Pathway. J. Nutr. Biochem. 2014, 25, 1207–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, M.; Xie, Q.; Zou, Y.; He, R.; Zhang, Y.; Wang, J.; Bo, L.; Li, J.; Deng, X. Alpha-Lipoic Acid Protects Mice against Concanavalin A-Induced Hepatitis by Modulating Cytokine Secretion and Reducing Reactive Oxygen Species Generation. Int. Immunopharmacol. 2016, 35, 53–60. [Google Scholar] [CrossRef]
- Mahmoud, Y.I.; Mahmoud, A.A.; Nassar, G. Alpha-Lipoic Acid Treatment of Acetaminophen-Induced Rat Liver Damage. Biotech. Histochem. 2015, 90, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Kaibori, M.; Miki, H.; Nakatake, R.; Tokuhara, K.; Nishizawa, M.; Okumura, T.; Kwon, A.-H. Alpha-Lipoic Acid Exerts a Liver-Protective Effect in Acute Liver Injury Rats. J. Surg. Res. 2015, 193, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liu, J.; Pian, L.; Gui, S.; Lu, B. A-lipoic Acid Protects against Carbon Tetrachloride-induced Liver Cirrhosis through the Suppression of the TGF-β/Smad3 Pathway and Autophagy. Mol. Med. Rep. 2019, 19, 841–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lexis, L.A.; Fassett, R.G.; Coombes, J.S. Alpha-Tocopherol and Alpha-Lipoic Acid Enhance the Erythrocyte Antioxidant Defence in Cyclosporine A-Treated Rats. Basic Clin. Pharmacol. Toxicol. 2006, 98, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Faheem, S.A.; El-Sayed, N.M.; Moustafa, Y.M.; Saeed, N.M.; Hazem, R.M. Pyrvinium Pamoate Ameliorates Cyclosporin A—Induced Hepatotoxicity via the Modulation of Wnt/β-Catenin Signaling and Upregulation of PPAR-γ. Int. Immunopharmacol. 2022, 104, 108538. [Google Scholar] [CrossRef]
- Khalaf, A.A.; Zaki, A.R.; Galal, M.K.; Ogaly, H.A.; Ibrahim, M.A.; Hassan, A. The Potential Protective Effect of α-Lipoic Acid against Nanocopper Particle-Induced Hepatotoxicity in Male Rats. Hum. Exp. Toxicol. 2017, 36, 881–891. [Google Scholar] [CrossRef]
- Draper, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M.A. Comparative Evaluation of Thiobarbituric Acid Methods for the Determination of Malondialdehyde in Biological Materials. Free Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- Koracevic, D.; Koracevic, G.; Djordjevic, V.; Andrejevic, S.; Cosic, V. Method for the Measurement of Antioxidant Activity in Human Fluids. J. Clin. Pathol. 2001, 54, 356–361. [Google Scholar] [CrossRef] [Green Version]
- Shukry, M.; Alotaibi, S.S.; Albogami, S.M.; Fathallah, N.; Farrag, F.; Dawood, M.A.O.; Gewaily, M.S. Garlic Alleviates the Injurious Impact of Cyclosporine-A in Male Rats through Modulation of Fibrogenic and Steroidogenic Genes. Animals 2020, 11, 64. [Google Scholar] [CrossRef]
- He, X.; Sun, J.; Huang, X. Expression of Caspase-3, Bax and Bcl-2 in Hippocampus of Rats with Diabetes and Subarachnoid Hemorrhage. Exp. Ther. Med. 2018, 15, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Iheagwam, F.N.; Batiha, G.E.-S.; Ogunlana, O.O.; Chinedu, S.N. Terminalia Catappa Extract Palliates Redox Imbalance and Inflammation in Diabetic Rats by Upregulating Nrf-2 Gene. Int. J. Inflamm. 2021, 2021, 9778486. [Google Scholar] [CrossRef] [PubMed]
- Nouri, A.; Heidarian, E.; Nikoukar, M. Effects of N-Acetyl Cysteine on Oxidative Stress and TNF-α Gene Expression in Diclofenac-Induced Hepatotoxicity in Rats. Toxicol. Mech. Methods 2017, 27, 561–567. [Google Scholar] [CrossRef]
- Sherif, I.O.; Al-Shaalan, N.H. Vildagliptin Attenuates Hepatic Ischemia/Reperfusion Injury via the TLR4/NF-ΚB Signaling Pathway. Oxid. Med. Cell Longev. 2018, 2018, 3509091. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.E.; Elswefy, S.E.; Rashed, L.A.; Younis, N.N.; Shaheen, M.A.; Ghanim, A.M.H. Bone Marrow-Derived Mesenchymal Stem Cells Effectively Regenerate Fibrotic Liver in Bile Duct Ligation Rat Model. Exp. Biol. Med. 2016, 241, 581–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzahrani, S.; Zaitone, S.A.; Said, E.; El-Sherbiny, M.; Ajwah, S.; Alsharif, S.Y.; Elsherbiny, N.M. Protective Effect of Isoliquiritigenin on Experimental Diabetic Nephropathy in Rats: Impact on Sirt-1/NFκB Balance and NLRP3 Expression. Int. Immunopharmacol. 2020, 87, 106813. [Google Scholar] [CrossRef]
- Fedchenko, N.; Reifenrath, J. Different Approaches for Interpretation and Reporting of Immunohistochemistry Analysis Results in the Bone Tissue—A Review. Diagn. Pathol. 2014, 9, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open Source Software for Digital Pathology Image Analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [Green Version]
- Abboud, G.; Kaplowitz, N. Drug-Induced Liver Injury. Drug Saf. 2007, 30, 277–294. [Google Scholar] [CrossRef]
- Lorber, M.I.; Van Buren, C.T.; Flechner, S.M.; Williams, C.; Kahan, B.D. Hepatobiliary and Pancreatic Complications of Cyclosporine Therapy in 466 Renal Transplant Recipients. Transplantation 1987, 43, 35–40. [Google Scholar] [CrossRef]
- Korolczuk, A.; Caban, K.; Amarowicz, M.; Czechowska, G.; Irla-Miduch, J. Oxidative Stress and Liver Morphology in Experimental Cyclosporine A-Induced Hepatotoxicity. Biomed. Res. Int. 2016, 2016, 5823271. [Google Scholar] [CrossRef] [Green Version]
- Briguori, C.; Condorelli, G. Effect of Erdosteine on Cyclosporine Induced Chronic Nephrotoxicity. Nephrourol. Mon. 2012, 4, 487–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bingul, I.; Olgac, V.; Bekpinar, S.; Uysal, M. The Protective Effect of Resveratrol against Cyclosporine A-Induced Oxidative Stress and Hepatotoxicity. Arch. Physiol. Biochem. 2021, 127, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, X.; Nepovimova, E.; Wang, Y.; Yang, H.; Kuca, K. Mechanism of Cyclosporine A Nephrotoxicity: Oxidative Stress, Autophagy, and Signalings. Food Chem. Toxicol. 2018, 118, 889–907. [Google Scholar] [CrossRef] [PubMed]
- Elsherbiny, N.M.; Said, E.; Atef, H.; Zaitone, S.A. Renoprotective Effect of Calycosin in High Fat Diet-Fed/STZ Injected Rats: Effect on IL-33/ST2 Signaling, Oxidative Stress and Fibrosis Suppression. Chem. Biol. Interact. 2020, 315, 108897. [Google Scholar] [CrossRef]
- El-Sherbiny, M.; Fahmy, E.K.; Eisa, N.H.; Said, E.; Elkattawy, H.A.; Ebrahim, H.A.; Elsherbiny, N.M.; Ghoneim, F.M. Nanogold Particles Suppresses 5-Flurouracil-Induced Renal Injury: An Insight into the Modulation of Nrf-2 and Its Downstream Targets, HO-1 and γ-GCS. Molecules 2021, 26, 7684. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, B.; Liu, C.; Wang, L.; Tang, J.; Hong, L. Protective Role of Nrf2 against Mechanical-Stretch-Induced Apoptosis in Mouse Fibroblasts: A Potential Therapeutic Target of Mechanical-Trauma-Induced Stress Urinary Incontinence. Int. Urogynecol. J. 2018, 29, 1469–1477. [Google Scholar] [CrossRef]
- Di Paola, R.; Modafferi, S.; Siracusa, R.; Cordaro, M.; D’Amico, R.; Ontario, M.L.; Interdonato, L.; Salinaro, A.T.; Fusco, R.; Impellizzeri, D.; et al. S-Acetyl-Glutathione Attenuates Carbon Tetrachloride-Induced Liver Injury by Modulating Oxidative Imbalance and Inflammation. Int. J. Mol. Sci. 2022, 23, 4429. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-Y.; Lee, J.-H.; Yun, D.-H.; Lee, Y.-M.; Kim, D.-K. Inhibitory Effects of Nodakenin on Inflammation and Cell Death in Lipopolysaccharide-Induced Liver Injury Mice. Phytomedicine 2021, 81, 153411. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Luo, Z.; Zhou, K.; Wu, Q.; Xiao, W.; Yu, Y.; Li, T. Schisandrae Chinensis Fructus Extract Protects against Hepatorenal Toxicity and Changes Metabolic Ions in Cyclosporine A Rats. Nat. Prod. Res. 2021, 35, 2915–2920. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 Signaling Pathway: Pivotal Roles in Inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Akool, E.-S. Molecular Mechanisms of the Protective Role of Wheat Germ Oil against Cyclosporin A-Induced Hepatotoxicity in Rats. Pharm. Biol. 2015, 53, 1311–1317. [Google Scholar] [CrossRef] [Green Version]
- Iwakiri, Y.; Kim, M.Y. Nitric Oxide in Liver Diseases. Trends Pharmacol. Sci. 2015, 36, 524–536. [Google Scholar] [CrossRef] [Green Version]
- El-Gowelli, H.M.; Helmy, M.W.; Ali, R.M.; El-Mas, M.M. Celecoxib Offsets the Negative Renal Influences of Cyclosporine via Modulation of the TGF-Β1/IL-2/COX-2/Endothelin ET(B) Receptor Cascade. Toxicol. Appl. Pharmacol. 2014, 275, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.K.; Huang, H.; Ndebele, K.; Salahudeen, A.K. Vitamin E Inhibits Renal MRNA Expression of COX II, HO I, TGFbeta, and Osteopontin in the Rat Model of Cyclosporine Nephrotoxicity. Transplantation 2001, 71, 331–334. [Google Scholar] [CrossRef]
- Rao, S.R.; Balaji, T.M.; Prakash, P.S.G.; Lavu, V. Elevated Levels of Cyclooxygenase 1 and 2 in Human Cyclosporine Induced Gingival Overgrowth. Prostaglandins Other Lipid Mediat. 2014, 113–115, 69–74. [Google Scholar] [CrossRef]
- Eissa, L.A.; Kenawy, H.I.; El-Karef, A.; Elsherbiny, N.M.; El-Mihi, K.A. Antioxidant and Anti-Inflammatory Activities of Berberine Attenuate Hepatic Fibrosis Induced by Thioacetamide Injection in Rats. Chem. Biol. Interact. 2018, 294, 91–100. [Google Scholar] [CrossRef]
- Justo, P.; Lorz, C.; Sanz, A.; Egido, J.; Ortiz, A. Intracellular Mechanisms of Cyclosporin A-Induced Tubular Cell Apoptosis. J. Am. Soc. Nephrol. 2003, 14, 3072–3080. [Google Scholar] [CrossRef] [Green Version]
- Saad, K.M.; Abdelrahman, R.S.; Said, E. Mechanistic Perspective of Protective Effects of Nilotinib against Cisplatin-Induced Testicular Injury in Rats: Role of JNK/Caspase-3 Signaling Inhibition. Environ. Toxicol. Pharmacol. 2020, 76, 103334. [Google Scholar] [CrossRef] [PubMed]
- Deger, M.; Kaya, B.; Akdogan, N.; Kaplan, H.M.; Bagir, E.; Izol, V.; Aridogan, I.A. Protective Effect of Dapagliflozin against Cyclosporine A-Induced Nephrotoxicity. Drug Chem. Toxicol. 2021, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tastekin, N.; Aydogdu, N.; Dokmeci, D.; Usta, U.; Birtane, M.; Erbas, H.; Ture, M. Protective Effects of L-Carnitine and Alpha-Lipoic Acid in Rats with Adjuvant Arthritis. Pharmacol. Res. 2007, 56, 303–310. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, E.-S.M.; Mansour, A.M.; El-Sawy, W.S. Alpha Lipoic Acid Prevents Doxorubicin-Induced Nephrotoxicity by Mitigation of Oxidative Stress, Inflammation, and Apoptosis in Rats. J. Biochem. Mol. Toxicol. 2017, 31, 2411–2502. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, M.; Żebrowska, E.; Nesterowicz, M.; Żendzian-Piotrowska, M.; Zalewska, A. α-Lipoic Acid Strengthens the Antioxidant Barrier and Reduces Oxidative, Nitrosative, and Glycative Damage, as Well as Inhibits Inflammation and Apoptosis in the Hypothalamus but Not in the Cerebral Cortex of Insulin-Resistant Rats. Oxid. Med. Cell Longev. 2022, 2022, 7450514. [Google Scholar] [CrossRef]
- Deveci, H.A.; Akyuva, Y.; Nur, G.; Nazıroğlu, M. Alpha Lipoic Acid Attenuates Hypoxia-Induced Apoptosis, Inflammation and Mitochondrial Oxidative Stress via Inhibition of TRPA1 Channel in Human Glioblastoma Cell Line. Biomed. Pharm. 2019, 111, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Amudha, G.; Josephine, A.; Sudhahar, V.; Varalakshmi, P. Protective Effect of Lipoic Acid on Oxidative and Peroxidative Damage in Cyclosporine A-Induced Renal Toxicity. Int. Immunopharmacol. 2007, 7, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Abd-Ellah, H.F.; Abou-Zeid, N.R.A. Role of Alpha-Lipoic Acid in Ameliorating Cyclosporine A-Induced Pancreatic Injury in Albino Rats: A Structural, Ultrastructural, and Morphometric Study. Ultrastruct. Pathol. 2017, 41, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Kabel, A.M.; Salama, S.A.; Alghorabi, A.A.; Estfanous, R.S. Amelioration of Cyclosporine-Induced Testicular Toxicity by Carvedilol and/or Alpha-Lipoic Acid: Role of TGF-Β1, the Proinflammatory Cytokines, Nrf2/HO-1 Pathway and Apoptosis. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1169–1181. [Google Scholar] [CrossRef] [PubMed]
- Aliomrani, M.; Mesripour, A.; Mehrjardi, A.S. Creatine and Alpha-Lipoic Acid Antidepressant-Like Effect Following Cyclosporine A Administration. Turk. J. Pharm. Sci. 2022, 19, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Tibullo, D.; Li Volti, G.; Giallongo, C.; Grasso, S.; Tomassoni, D.; Anfuso, C.D.; Lupo, G.; Amenta, F.; Avola, R.; Bramanti, V. Biochemical and Clinical Relevance of Alpha Lipoic Acid: Antioxidant and Anti-Inflammatory Activity, Molecular Pathways and Therapeutic Potential. Inflamm. Res. 2017, 66, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, J.; Lodge, J.K.; Marcocci, L.; Tritschler, H.J.; Packer, L.; Rihn, B.H. Alpha-Lipoic Acid in Liver Metabolism and Disease. Free Radic. Biol. Med. 1998, 24, 1023–1039. [Google Scholar] [CrossRef]
- Silvestri, S.; Orlando, P.; Armeni, T.; Padella, L.; Brugè, F.; Seddaiu, G.; Littarru, G.P.; Tiano, L. Coenzyme Q10 and α-Lipoic Acid: Antioxidant and pro-Oxidant Effects in Plasma and Peripheral Blood Lymphocytes of Supplemented Subjects. J. Clin. Biochem. Nutr. 2015, 57, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigil, M.; Berkson, B.M.; Garcia, A.P. Adverse Effects of High Doses of Intravenous Alpha Lipoic Acid on Liver Mitochondria. Glob. Adv. Health Med. 2014, 3, 25–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Control + α-LA | CsA | CsA + α-LA |
|---|---|---|---|---|
| ALT(IU/L) | 41.7 ± 8.1 | 34.4 ± 9.5 | 54.1 ± 13.3 * | 43.1 ± 14.2 # |
| AST(IU/L) | 54.1 ± 9.5 | 47.2 ± 11.1 | 77.2 ± 21.8 * | 61.1 ± 2.4 # |
| GGT(IU/L) | 23.4 ± 7.2 | 22.3 ± 6.1 | 38.1 ± 1.7 * | 29.6 ± 1.2 # |
| ALP(IU/L) | 63.5 ± 12.4 | 59.5 ± 11.5 | 75.3 ± 12.5 * | 63.2 ± 12.2 # |
| Parameter | Control | Control + α-LA | CsA | CsA + α-LA |
|---|---|---|---|---|
| TBARS (μmol MDA/L) | 24.3 ± 4 | 22.7 ± 2.8 | 39.4 ± 9.3 * | 28.7 ± 7.5 # |
| TAC (mmol/L) | 629.7 ± 157.8 | 693.8 ± 156.8 | 377 ± 98.9 * | 600 ± 124.3 # |
| Parameter | Control | Control + α-LA | CsA | CsA + α-LA |
|---|---|---|---|---|
| TBARS (nmol/g.tissue) | 12.6 ± 1.9 | 13.7 ± 2.5 | 35.9 ± 2.2 * | 21.5 ± 1.2 # |
| SOD (U/g.tissue) | 139.8 ± 5.2 | 125.7 ± 11.2 | 69.08 ± 4.2 * | 106.1 ± 4.7 # |
| CAT (U/g.tissue) | 5 ± 0.58 | 4.5 ± 0.37 | 1.7 ± 0.06 * | 3.2 ± 0.09 # |
| GSH (mmol/g.tissue) | 2.1 ± 0.08 | 2.5 ± 0.03 | 0.88 ± 0.16 * | 1.5 ± 0.15 # |
| Parameter | Control | Control + α-LA | CsA | CsA + α-LA |
|---|---|---|---|---|
| SOD gene expression (2−∆∆CT) | 0.99 ± 0.01 | 1.01 ± 0.02 | 0.53 ± 0.19 * | 0.79 ± 0.23 # |
| Nrf-2 gene expression (2−∆∆CT) | 1.05 ± 0.04 | 1.03 ± 0.02 | 0.47 ± 0.11 * | 0.88 ± 0.27 # |
| Parameter | Control | Control + α-LA | CsA-Treated Group | CsA + α-LA |
|---|---|---|---|---|
| TNF-α gene expression (2−∆∆CT) | 0.98 ± 0.03 | 0.01 ± 0.05 | 2.57 ± 0.64 * | 1.61 ± 0.37 # |
| NF-κB gene expression (2−∆∆CT) | 1.03 ± 0.05 | 1.06 ± 0.04 | 2.38 ± 0.74 * | 1.53 ± 0.41 # |
| NO (μmol/L) | 19.6 ± 4 | 20.2 ± 5 | 9.3 ± 3 * | 17.9 ± 2 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Mancy, E.M.; Elsherbini, D.M.A.; Al-Serwi, R.H.; El-Sherbiny, M.; Ahmed Shaker, G.; Abdel-Moneim, A.-M.H.; Enan, E.T.; Elsherbiny, N.M. α-Lipoic Acid Protects against Cyclosporine A-Induced Hepatic Toxicity in Rats: Effect on Oxidative Stress, Inflammation, and Apoptosis. Toxics 2022, 10, 442. https://doi.org/10.3390/toxics10080442
El-Mancy EM, Elsherbini DMA, Al-Serwi RH, El-Sherbiny M, Ahmed Shaker G, Abdel-Moneim A-MH, Enan ET, Elsherbiny NM. α-Lipoic Acid Protects against Cyclosporine A-Induced Hepatic Toxicity in Rats: Effect on Oxidative Stress, Inflammation, and Apoptosis. Toxics. 2022; 10(8):442. https://doi.org/10.3390/toxics10080442
Chicago/Turabian StyleEl-Mancy, Eman M., Dalia Mahmoud Abdelmonem Elsherbini, Rasha Hamed Al-Serwi, Mohamed El-Sherbiny, Gehan Ahmed Shaker, Abdel-Moneim Hafez Abdel-Moneim, Eman T. Enan, and Nehal M. Elsherbiny. 2022. "α-Lipoic Acid Protects against Cyclosporine A-Induced Hepatic Toxicity in Rats: Effect on Oxidative Stress, Inflammation, and Apoptosis" Toxics 10, no. 8: 442. https://doi.org/10.3390/toxics10080442