
Biodegradation of Dibutyl Phthalate by the New Strain Acinetobacter baumannii DP-2

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Medium
2.1.1. Chemicals
2.1.2. Culture Medium
2.2. Isolation of DBP-Degrading Bacteria
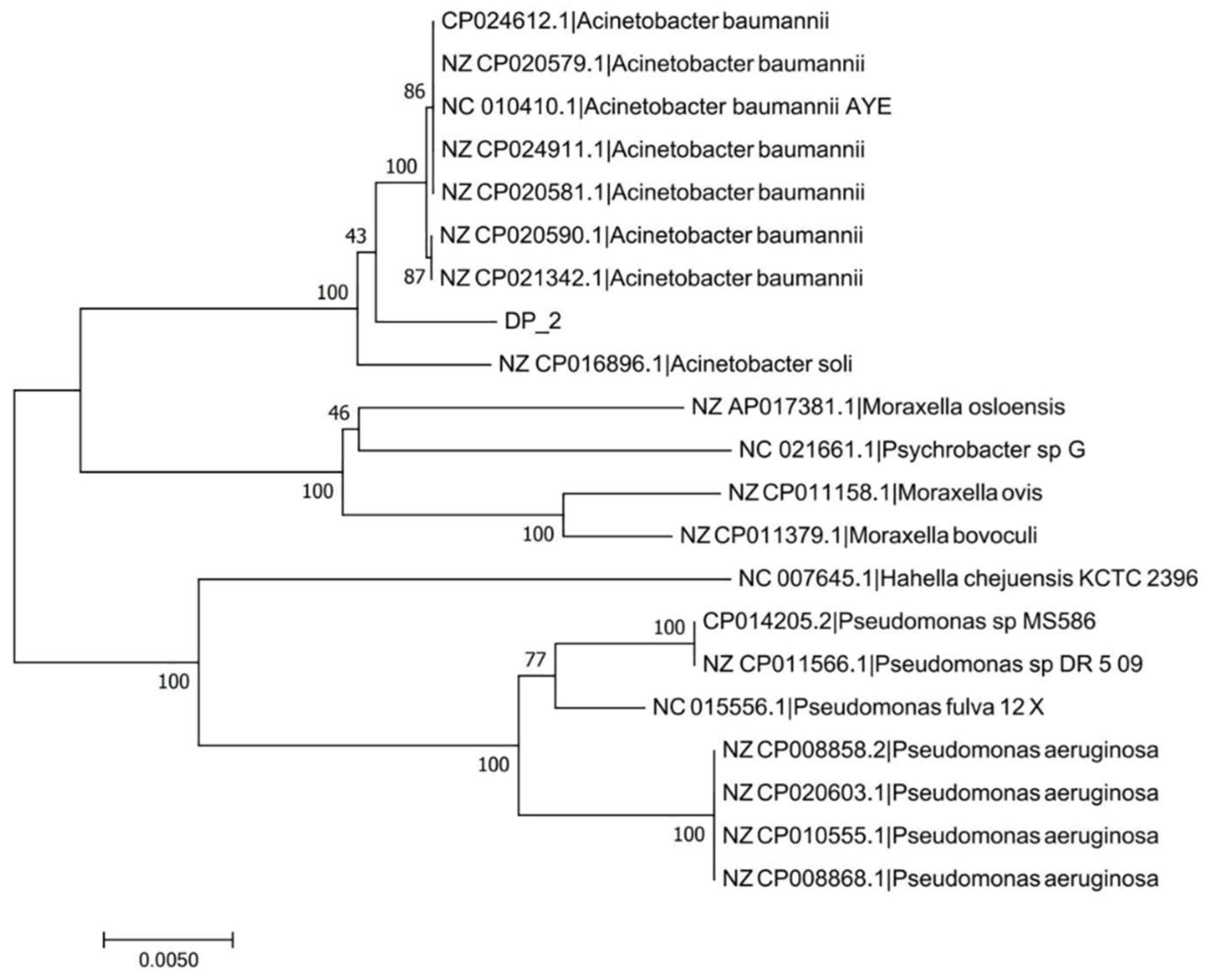
2.3. 16S rRNA Gene Amplification, Sequencing, and Phylogenetic Analysis
2.4. Optimization of Culture Conditions of DP-2
2.4.1. Single-Factor Experiments
2.4.2. Response Surface Optimization Experiments
2.5. Identification of the Degradation Intermediate Products of DP-2
2.6. Calculations
3. Results
3.1. Identification and Characterization of DP-2
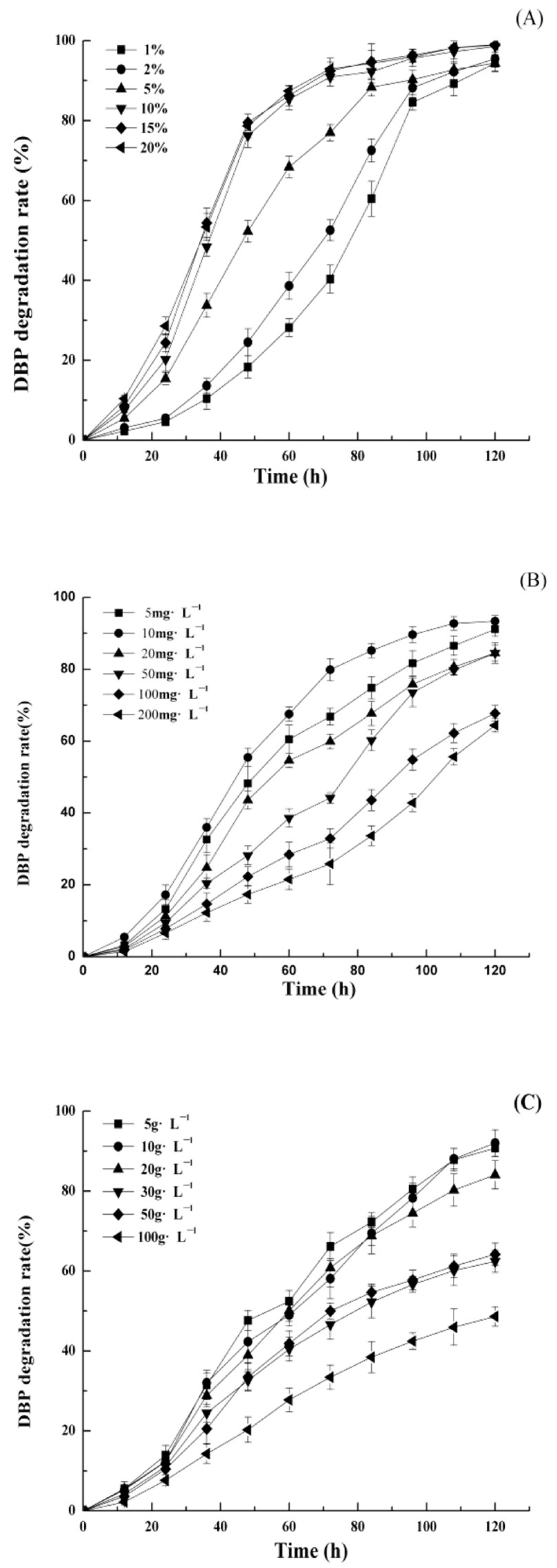
3.2. Effects of Culture Conditions on the DBP Degradation Rates of DP-2
3.2.1. Inoculation Amount
3.2.2. DBP Concentration
3.2.3. NaCl Concentration
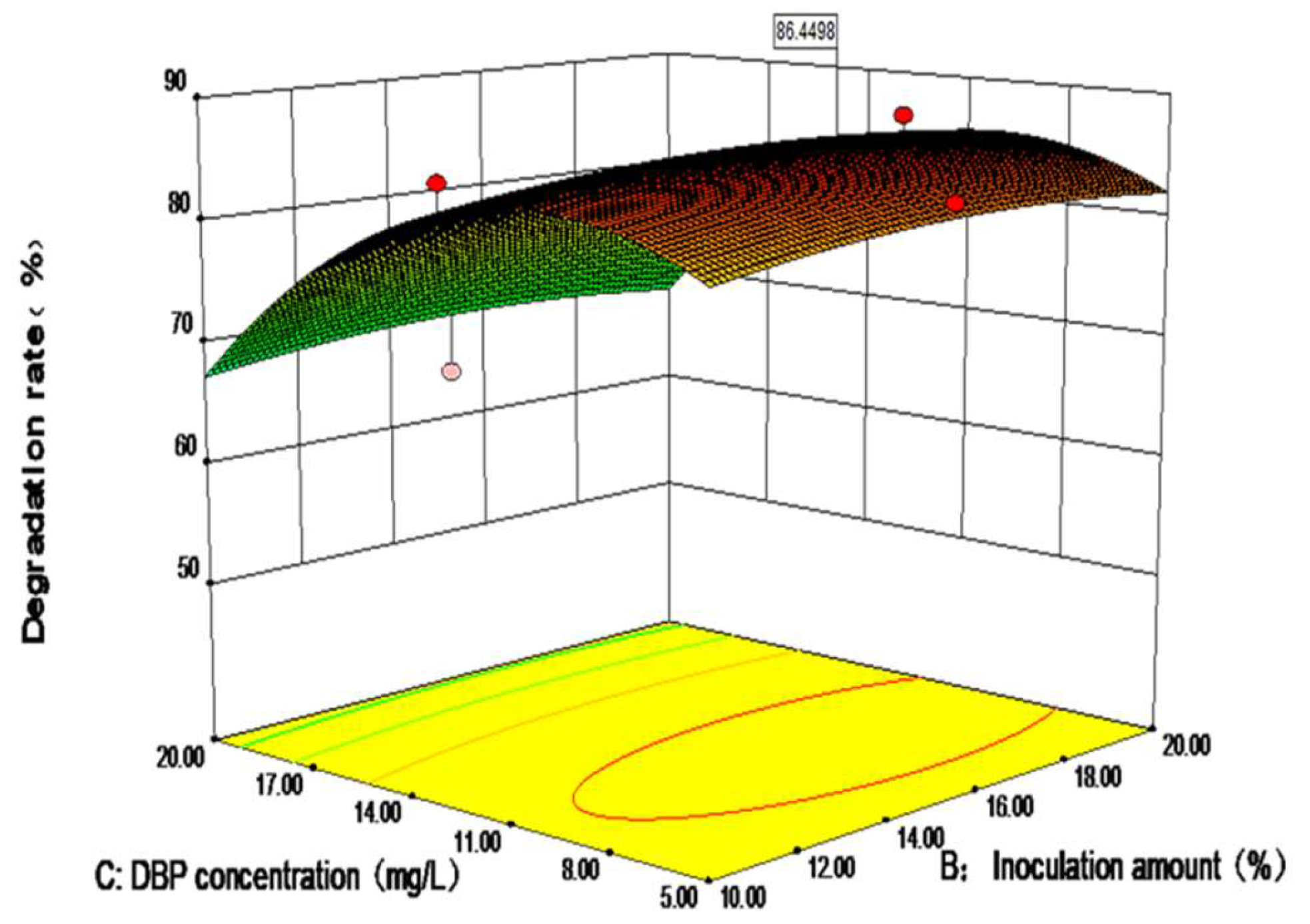
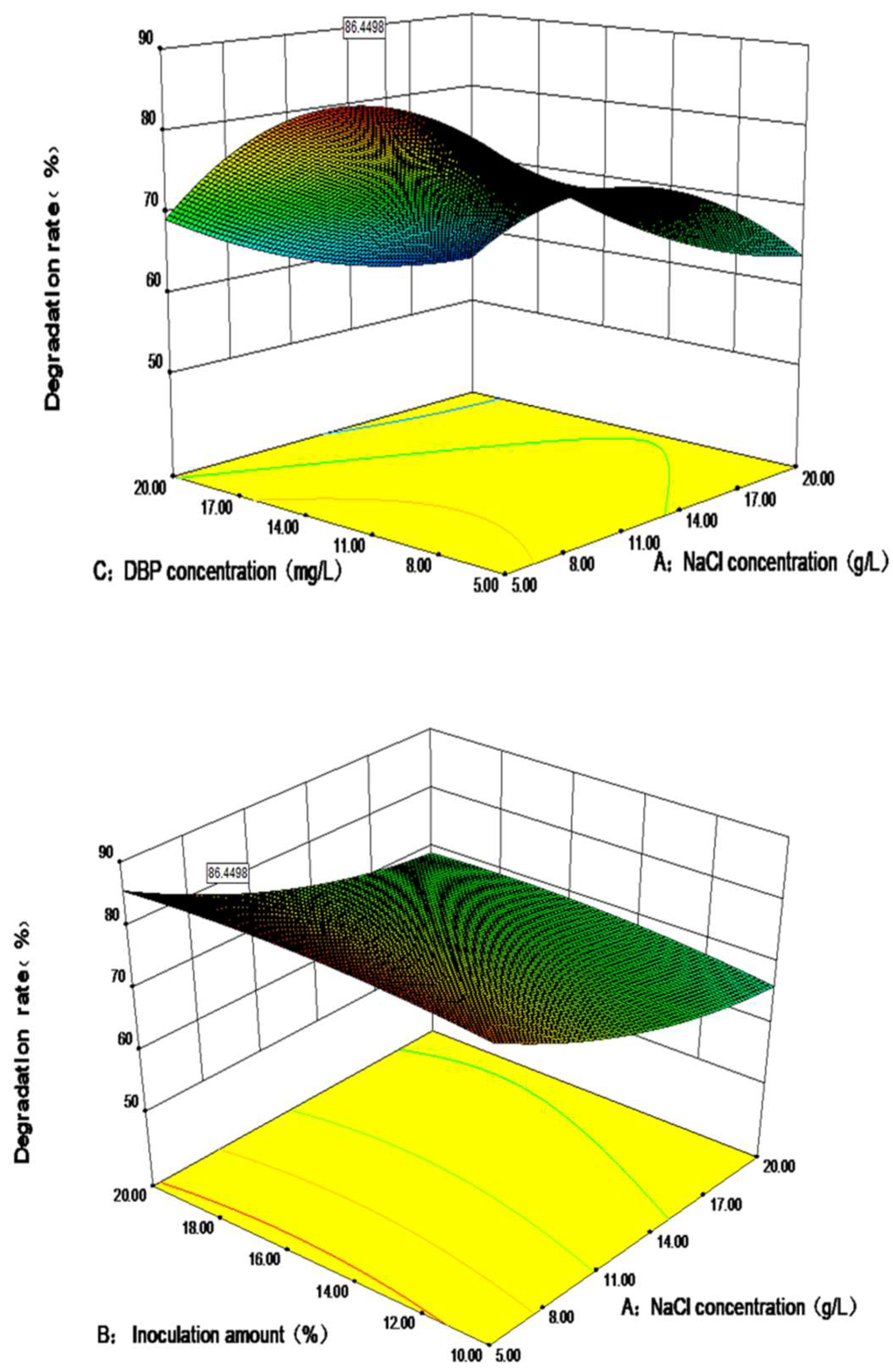
3.3. Optimization of the Degradation Performance of DP-2 via the Response Surface Method
3.4. The DBP Biodegradation Kinetics of Acinetobacter baumannii DP-2
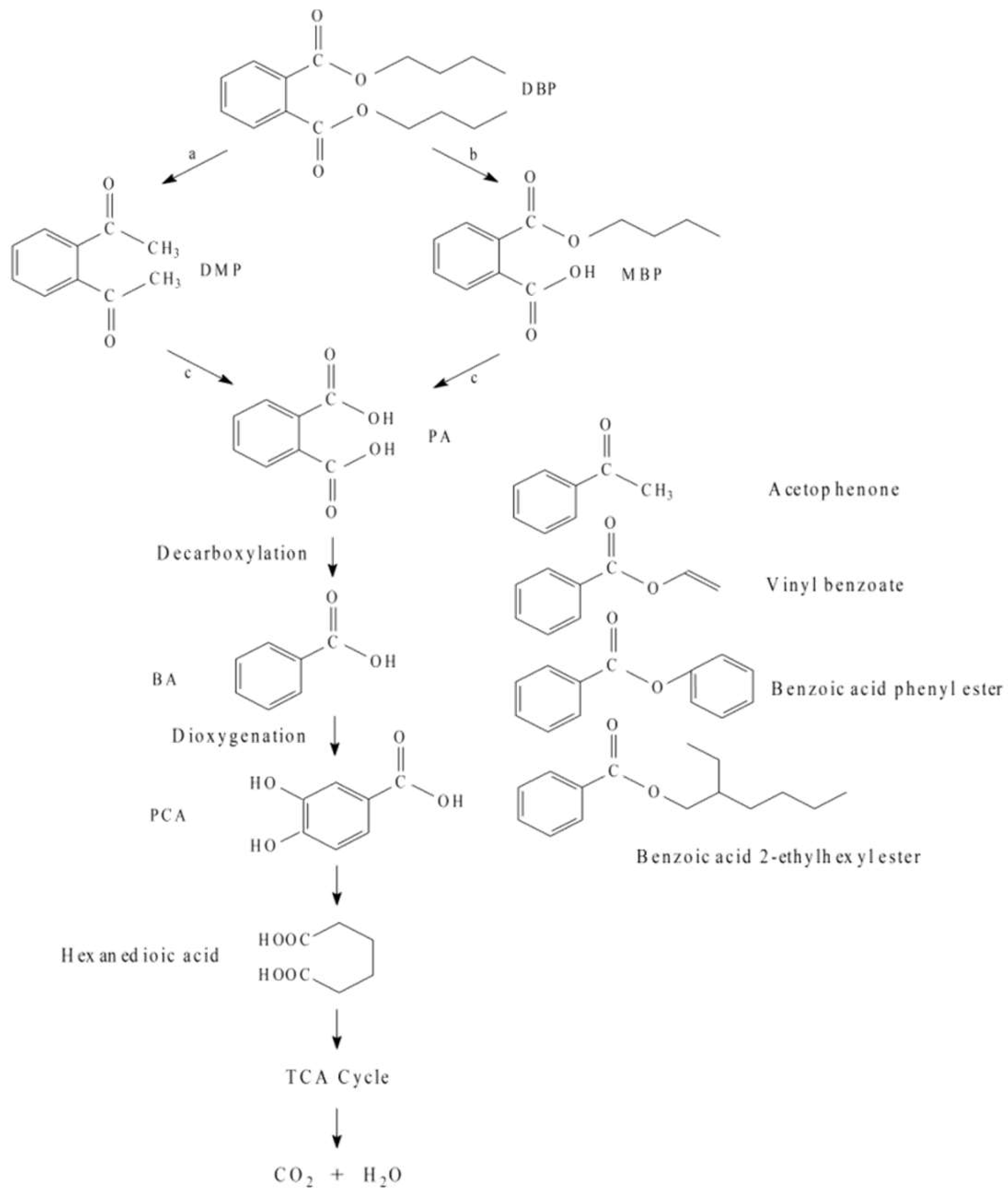
3.5. The DBP Biodegradation Pathway of Acinetobacter baumannii DP-2
4. Discussion
4.1. The DBP Biodegradation Ability of Acinetobacter baumannii DP-2
4.2. Biodegradation Pathways of Acinetobacter baumannii DP-2
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, X.Y.; Wang, P.Y.; Wu, Y.C.; Zhou, Y.; Sheng, Y.F.; Lao, K.J. Microplastic acts as a vector for contaminants: The release behavior of dibutyl phthalate from polyvinyl chloride pipe fragments in water phase. Environ. Sci. Pollut. Res. 2020, 27, 42082–42091. [Google Scholar] [CrossRef]
- Wu, E.; Gu, J.D. Biodegradability of synthetic plastics and polymeric materials: An illusion or reality in waste managements. Appl. Environ. Biotechnol. 2020, 5, 9–27. [Google Scholar] [CrossRef]
- Feng, N.X.; Yu, J.; Mo, C.H.; Zhao, M.H.; Li, Y.W.; Wu, B.X.; Cai, Q.Y.; Li, H.; Zhou, D.M.; Wong, M.H. Biodegradation of di-n-butyl phthalate (DBP) by a novel endophytic Bacillus megaterium strain YJB3. Sci. Total Environ. 2018, 616, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.X.; Feng, Y.X.; Liang, Q.F.; Chen, X.; Xiang, L.; Zhao, H.M.; Liu, B.L.; Cao, G.; Li, Y.W.; Li, H.; et al. Complete biodegradation of di-n-butyl phthalate (DBP) by a novel Pseudomonas sp.YJB6. Sci. Total Environ. 2020, 761, 143208. [Google Scholar] [CrossRef]
- He, Z.F.; Li, Z.Y.; Zhang, Q.Y.; Wei, Z.; Duo, J.; Pan, X.L. Simultaneous remediation of As(III) and dibutyl phthalate (DBP) in soil by a manganese-oxidizing bacterium and its mechanisms. Chemosphere 2019, 220, 837–844. [Google Scholar] [CrossRef]
- Li, Y.T.; Wang, J.; Yang, S.; Zhang, S. Occurrence, health risks and soil-air exchange of phthalate acid esters: A case study in plastic film greenhouses of Chongqing, China. Chemosphere 2020, 268, 128821. [Google Scholar] [CrossRef]
- Ma, T.T.; Wu, L.H.; Chen, L.; Zhang, H.B.; Teng, Y.; Luo, Y.M. Phthalate esters contamination in soils and vegetables of plastic film greenhouses of suburb Nanjing, China and the potential human health risk. Environ. Sci. Pollut. Res. 2015, 22, 12018–12028. [Google Scholar] [CrossRef]
- Wei, S.; Wang, F.H.; Chen, Y.J.; Tao, L.; Zhang, S.T. The joint toxicity effect of five antibiotics and dibutyl phthalate to luminescent bacteria (Vibrio fischeri). Environ. Sci. Pollut. Res. 2018, 25, 26504–26511. [Google Scholar] [CrossRef]
- Shi, M.; Sun, Y.Y.; Wang, Z.H.; He, G.; Quan, H.X.; He, H.X. Plastic film mulching increased the accumulation and human health risks of phthalate esters in wheat grains. Environ. Pollut. 2019, 250, 1–7. [Google Scholar] [CrossRef]
- Qi, X.; Li, T.C.; Wang, F.H.; Dai, Y.R.; Liang, W. Removal efficiency and enzymatic mechanism of dibutyl phthalate (DBP) by constructed wetlands. Environ. Sci. Pollut. Res. 2018, 25, 23009–23017. [Google Scholar] [CrossRef]
- Gao, D.W.; Wen, Z.D. Phthalate esters in the environment: A critical review of their occurrence, biodegradation, and removal during wastewater treatment processes. Sci. Total Environ. 2016, 541, 986–1001. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Guo, C.L.; Liu, S.S.; Liu, W.T.; Wang, H.; Dang, Z.; Lu, G.N. Characterization of a di-n-butyl phthalate-degrading bacterial consortium and its application in contaminated soil. Environ. Sci. Pollut. Res. 2018, 25, 17645–17653. [Google Scholar] [CrossRef]
- Fan, S.H.; Wang, J.H.; Li, K.; Yang, T.; Jia, Y.; Zhao, B.S.; Yan, Y.C. Complete genome sequence of Gordonia sp. YC-JH1, a bacterium efficiently degrading a wide range of phthalic acid esters. J. Biotechnol. 2018, 279, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.C.; Zhang, Y.; Wang, L.; Li, X.Q.; Chen, Y.Y.; Qu, J.H.; Zhen, K. Uptake mechanism of di-n-butyl phthalate by Novosphingobium species DNB-S3. Int. Biodeterior. Biodegrad. 2020, 148, 104910. [Google Scholar] [CrossRef]
- Tang, W.J.; Zhang, L.S.; Fang, Y.; Zhou, Y.; Ye, B.C. Biodegradation of phthalate esters by newly isolated Rhizobium sp. LMB-1 and its biochemical pathway of di-n-butyl phthalate. J. Appl. Microbiol. 2016, 121, 177–186. [Google Scholar] [CrossRef]
- Cai, M.Y.; Dong, X.Z. Common Bacterial System Identification Manual; Science Press: Beijing, China, 2001; pp. 67–105. [Google Scholar]
- Huang, Y.H.; Huang, X.J.; Chen, X.H.; Cai, Q.Y.; Chen, S.H.; Mo, C.H.; Lü, H.X.; Wong, M.H. Biodegradation of di-butyl phthalate (DBP) by a novel endophytic bacterium Bacillus subtilis and its bioaugmentation for removing DBP from vegetation slurry. J. Environ. Manag. 2018, 224, 1–9. [Google Scholar] [CrossRef]
- Wu, J.; Liao, X.W.; Yu, F.B.; Wei, Z.B.; Yang, L.Y. Cloning of a dibutyl phthalate hydrolase gene from Acinetobacter sp. strain M673 and functional analysis of its expression product in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 2483–2491. [Google Scholar] [CrossRef]
- Xu, J.M.; Lu, Q.H.; Toledo, R.A.D.; Shim, H. Degradation of di-2-ethylhexyl phthalate (DEHP) by an indigenous isolate Acinetobacter sp. SN13. Int. Biodeterior. Biodegrad. 2017, 117, 205–214. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, L.S.; Wang, J.; Zhou, Y.; Ye, B.C. Biodegradation of Phthalate Esters by a Newly Isolated Acinetobacter sp. Strain LMB-5 and Characteristics of Its Esterase. Pedosphere 2017, 27, 606–615. [Google Scholar] [CrossRef]
- Chen, X.P.; Xu, S.S.; Tan, T.F.; Lee, S.T.; Cheng, S.H.; Lee, F.W.; Xu, S.J.L.; Ho, K.C. Toxicity and estrogenic endocrine disrupting activity of phthalates and their mixtures. Int. J. Environ. Res. Public Health 2014, 11, 3156–3168. [Google Scholar] [CrossRef]
- Hsu, P.C.; Kuo, Y.T.; Guo, Y.L.L.; Chen, J.R.; Tsai, S.S.; Chao, H.R.; Teng, Y.N.; Pan, M.H. The adverse effects of low-dose exposure to di(2-ethylhexyl) phthalate during adolescence on sperm function in adult rats. Environ. Toxicol. 2016, 31, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Lin, Z.; Liu, H.M.; Hu, H.Q. Bacteria-mediated phthalic acid esters degradation and related molecular mechanisms. Appl. Environ. Biotechnol. 2018, 102, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, L.; Lin, Y.L.; Liu, W.X.; Tuyiringire, D.; Jiao, Y.Q.; Zhang, L.; Meng, Q.J.; Zhang, Y. Complete metabolic study by dibutyl phthalate degrading Pseudomonas sp. DNB-S1. Ecotoxicol. Environ. Saf. 2020, 194, 110378. [Google Scholar] [CrossRef]
- Zhang, J.F.; Zhang, C.N.; Zhu, Y.P.; Li, J.L.; Li, X.T. Biodegradation of seven phthalate esters by Bacillus mojavensis B1811. Int. Biodeterior. Biodegrad. 2018, 132, 200–207. [Google Scholar] [CrossRef]
- Benjamin, S.; Pradeep, S.; Josh, M.S.; Kumar, S.; Masai, E. A monograph on the remediation of hazardous phthalates. J. Hazard. Mater. 2015, 298, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Yan, J.L.; Wang, L.; Zhang, Y.Z.; Liu, D.L.; Geng, H.; Xiong, L. Characterization of the phthalate acid catabolic gene cluster in phthalate acid esters transforming bacterium-Gordonia sp. strain HS-NH1. Int. Biodeterior. Biodegrad. 2016, 106, 34–40. [Google Scholar] [CrossRef]
- Zhao, H.M.; Du, H.; Feng, N.X.; Xiang, L.; Li, Y.W.; Li, H.; Cai, Q.Y.; Mo, C.H. Biodegradation of di-n-butylphthalate and phthalic acid by a novel Providencia sp. 2D and its stimulation in a compost-amended soil. Biol. Fertil. Soils 2016, 52, 65–76. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.L.; Yang, Y.; Yue, D.M.; Xiao, L.; Yang, L.Y. Biodegradation of an endocrine-disrupting chemical di-n-butyl phthalate by newly isolated Camelimonas sp. and enzymatic properties of its hydrolase. Biodegradation 2015, 26, 171–182. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Code | Level | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| NaCl concentration (g·L−1) | A | 5 | 10 | 20 |
| Inoculum (%) | B | 10 | 15 | 20 |
| Initial DBP concentration (mg·L−1) | C | 5 | 10 | 20 |
| Physiological and Biochemical Indexes | Test Results | Physiological and Biochemical Indexes | Test Results |
|---|---|---|---|
| Glucose fermentation (acid production) | + | Hydrogen peroxide | + |
| Glucose fermentation (gas production) | − | ||
| Fructose fermentation (acid production) | + | Nitrate reduction | − |
| Oxidase | − | Urease | − |
| Sucrose fermentation | + | Methyl red experiment | + |
| Mannose fermentation | + | Acetyl methanol test | − |
| Mannitol fermentation | + | Xylose fermentation | − |
| Hydrogen sulfide production | − | Gelatin liquefaction | − |
| Lactose fermentation | − | Tween 80 | − |
| Arabinose fermentation | − | Starch hydrolysis | + |
| Gossypose fermentation | − | Indole experiment | − |
| Inositol | − | Citrate utilization | + |
| No. | A (Inoculum Amount) | B (Initial DBP Concentration) | C (NaCl Concentration) | Degradation Rate (%) |
|---|---|---|---|---|
| 1 | 1 | 1 | 0 | 66.78 |
| 2 | 0 | 1 | 1 | 62.23 |
| 3 | −1 | 0 | 1 | 64.05 |
| 4 | 0 | −1 | 1 | 60.11 |
| 5 | −1 | 0 | −1 | 83.64 |
| 6 | 1 | −1 | 0 | 64.48 |
| 7 | 0 | 0 | 0 | 74.43 |
| 8 | 1 | 0 | 1 | 54.83 |
| 9 | −1 | 1 | 0 | 86.23 |
| 10 | −1 | −1 | 0 | 85.36 |
| 11 | 0 | 1 | −1 | 66.87 |
| 12 | 1 | 0 | −1 | 68.73 |
| 13 | 0 | 0 | 0 | 73.66 |
| 14 | 0 | 0 | 0 | 75.23 |
| 15 | 0 | 0 | 0 | 72.97 |
| 16 | 0 | 0 | 0 | 73.56 |
| 17 | 0 | −1 | −1 | 64.22 |
| Source | Sum of Squares | df | Mean Square Error | F-Value | p-Value | Significance |
|---|---|---|---|---|---|---|
| Model | 1189.37 | 9 | 132.15 | 7.93 | 0.0062 | * |
| A-salinity | 535.63 | 1 | 535.63 | 32.16 | 0.0008 | ** |
| B-inoculum | 9.99 | 1 | 9.99 | 0.60 | 0.4640 | |
| C-substrate concentration | 223.03 | 1 | 223.03 | 13.39 | 0.0081 | * |
| AB | 0.046 | 1 | 0.046 | 2.775 × 10−3 | 0.9595 | |
| AC | 8.09 | 1 | 8.09 | 0.49 | 0.5082 | |
| BC | 0.070 | 1 | 0.070 | 4.216 × 10−3 | 0.9500 | |
| A2 | 43.76 | 1 | 43.76 | 2.63 | 0.1491 | |
| B2 | 6.38 | 1 | 6.38 | 0.38 | 0.5555 | |
| C2 | 370.56 | 1 | 370.56 | 22.25 | 0.0022 | ** |
| Residual error | 116.60 | 7 | 16.66 | |||
| Lack of fit items | 113.53 | 3 | 37.84 | 49.42 | 0.1013 | |
| Pure error | 3.06 | 4 | 0.77 | |||
| Total error | 1305.96 | 16 |
| Initial Concentration (mg·L−1) | Kinetic Equation | Kinetic Parameter (K/h−1) | t1/2/h | R2 |
|---|---|---|---|---|
| 5 | lnC = −0.0193t + 1.8746 | 0.0193 | 15.91 | 0.9927 |
| 10 | lnC = −0.0277t + 2.7451 | 0.0277 | 25.02 | 0.9869 |
| 20 | lnC = −0.0191t + 3.3530 | 0.0191 | 36.28 | 0.9856 |
| 50 | lnC = −0.0187t + 4.3777 | 0.0187 | 37.06 | 0.9631 |
| 100 | lnC = −0.0115t + 4.8737 | 0.0115 | 60.26 | 0.9641 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Liu, C.; Li, R.; Liu, Y.; Xie, J.; Li, B. Biodegradation of Dibutyl Phthalate by the New Strain Acinetobacter baumannii DP-2. Toxics 2022, 10, 532. https://doi.org/10.3390/toxics10090532
Li C, Liu C, Li R, Liu Y, Xie J, Li B. Biodegradation of Dibutyl Phthalate by the New Strain Acinetobacter baumannii DP-2. Toxics. 2022; 10(9):532. https://doi.org/10.3390/toxics10090532
Chicago/Turabian StyleLi, Cheng, Chunjing Liu, Rongzhen Li, Yue Liu, Jianzhi Xie, and Bowen Li. 2022. "Biodegradation of Dibutyl Phthalate by the New Strain Acinetobacter baumannii DP-2" Toxics 10, no. 9: 532. https://doi.org/10.3390/toxics10090532
APA StyleLi, C., Liu, C., Li, R., Liu, Y., Xie, J., & Li, B. (2022). Biodegradation of Dibutyl Phthalate by the New Strain Acinetobacter baumannii DP-2. Toxics, 10(9), 532. https://doi.org/10.3390/toxics10090532