
Perfluorooctanoic Acid Promotes Recruitment and Exocytosis of Rodlet Cells in the Renal Hematopoietic Tissue of Common Carp




Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Experimental Design
2.2. Light and Transmission Electron Microscopy
2.3. Biometry
2.4. Statistics
3. Results
3.1. Light Microscopy
3.2. Biometry
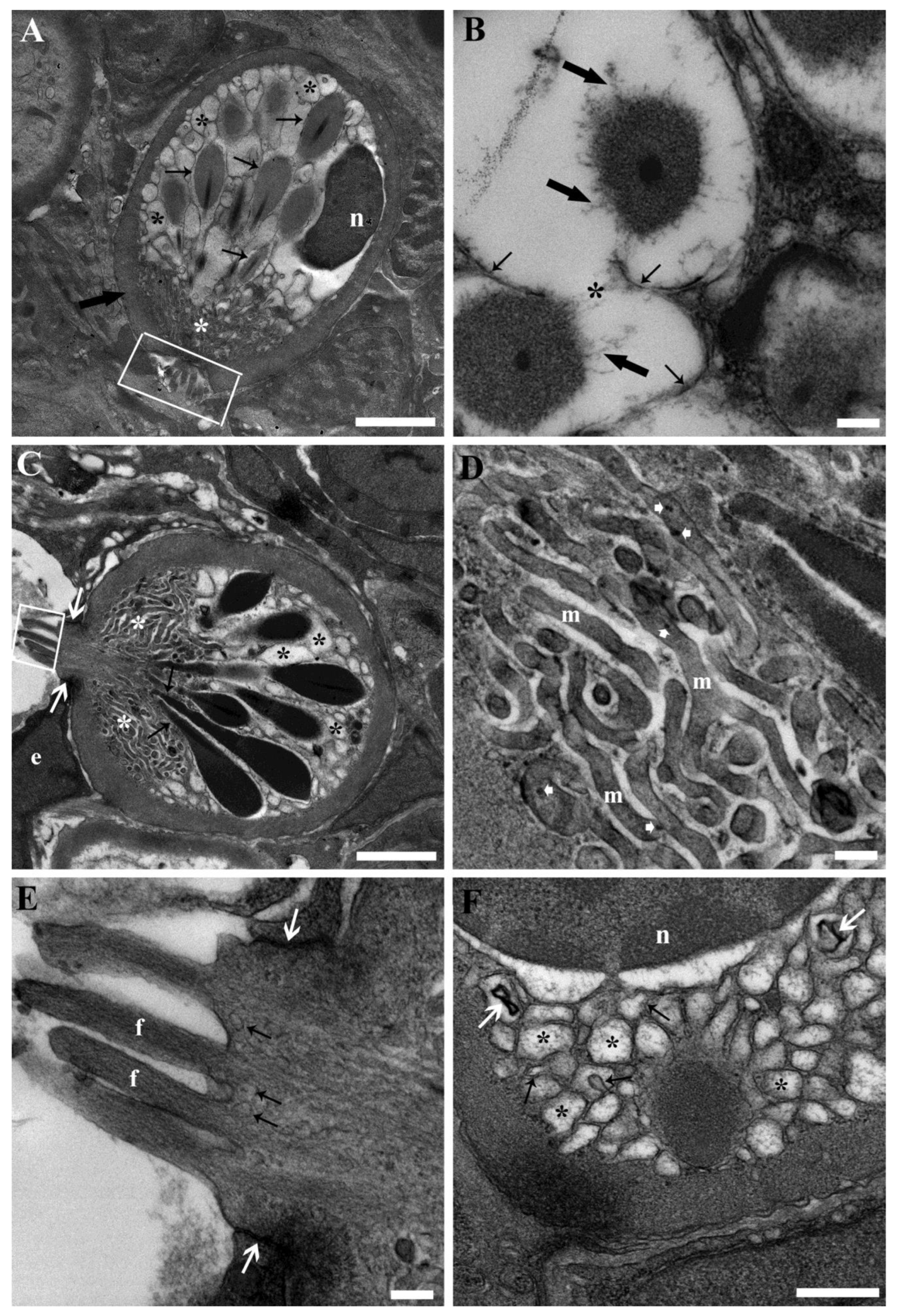
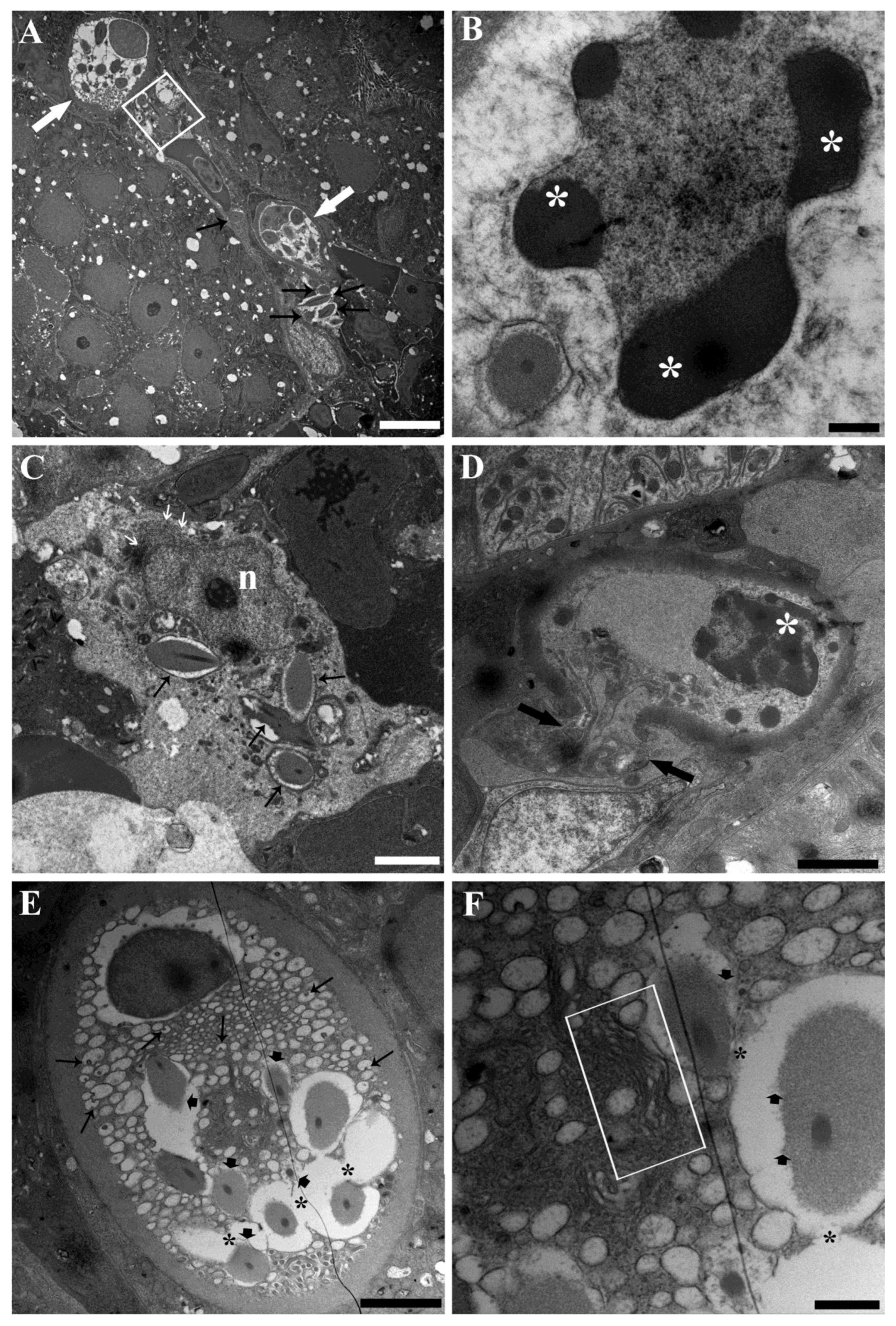
3.3. Ultrastructure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cousins, I.T.; Dewitt, J.C.; Glüge, J.; Goldenman, G.; Herzke, D.; Lohmann, R.; Miller, M.; Ng, C.A.; Scheringer, M.; Vierke, L.; et al. Strategies for grouping per-and polyfluoroalkyl substances (PFAS) to protect human and environmental health. Environ. Sci. Process. Impacts 2020, 22, 1444–1460. [Google Scholar] [CrossRef]
- Ding, G.; Peijnenburg, W.J.G.M. Physicochemical properties and aquatic toxicity of poly- and perfluorinated compounds. Crit. Rev. Environ. Sci. Technol. 2013, 43, 598–678. [Google Scholar] [CrossRef]
- Sinclair, G.M.; Long, S.M.; Jones, O.A.H. What are the effects of PFAS exposure at environmentally relevant concentrations? Chemosphere 2020, 258, 127340. [Google Scholar] [CrossRef]
- Evich, M.G.; Davis, M.J.B.; McCord, J.P.; Acrey, B.; Awkerman, J.A.; Knappe, D.R.U.; Lindstrom, A.B.; Speth, T.F.; Tebes-Stevens, C.; Strynar, M.J.; et al. Per- and polyfluoroalkyl substances in the environment. Science 2022, 375, eabg9065. [Google Scholar] [CrossRef]
- Wee, S.Y.; Aris, A.Z. Revisiting the “forever chemicals”, PFOA and PFOS exposure in drinking water. NPJ Clean Water 2023, 6, 57. [Google Scholar] [CrossRef]
- Dong, H.; Lu, G.; Wang, X.; Zhang, P.; Yang, H.; Yan, Z.; Liu, J.; Jiang, R. Tissue-specific accumulation, depuration, and effects of perfluorooctanoic acid on fish: Influences of aqueous pH and sex. Sci. Total Environ. 2023, 861, 160567. [Google Scholar] [CrossRef]
- Ahrens, L. Polyfluoroalkyl compounds in the aquatic environment: A review of their occurrence and fate. J. Environ. Monit. 2011, 13, 20–31. [Google Scholar] [CrossRef]
- Galloway, J.E.; Moreno, A.V.P.; Lindstrom, A.B.; Strynar, M.J.; Newton, S.; May, A.A.; May, A.A.; Weavers, L.K.; Weavers, L.K. Evidence of air dispersion: HFPO-DA and PFOA in Ohio and West Virginia surface water and soil near a fluoropolymer production facility. Environ. Sci. Technol. 2020, 54, 7175–7184. [Google Scholar] [CrossRef]
- Fang, Z.; Li, Y.; Li, Y.; Yang, D.; Zhang, H.; Jones, K.C.; Gu, C.; Luo, J. Development and applications of novel DGT passive samplers for measuring 12 per- and polyfluoroalkyl substances in natural waters and wastewaters. Environ. Sci. Technol. 2021, 55, 9548–9556. [Google Scholar] [CrossRef]
- Lee, J.W.; Choi, K.; Park, K.; Sung, C.; Yu, S.D.; Kim, P.; Seong, C.; Yu, S.D.; Kim, P. Adverse effects of perfluoroalkyl acids on fish and other aquatic organisms: A review. Sci. Total Environ. 2020, 707, 135334. [Google Scholar] [CrossRef]
- Du, D.; Lu, Y.; Zhou, Y.; Li, Q.; Zhang, M.; Han, G.; Cui, H.; Jeppesen, E. Bioaccumulation, trophic transfer and biomagnification of perfluoroalkyl acids (PFAAs) in the marine food web of the South China Sea. J. Hazard. Mater. 2021, 405, 124681. [Google Scholar] [CrossRef]
- Sunderland, E.M.; Hu, X.C.; Dassuncao, C.; Tokranov, A.K.; Wagner, C.C.; Allen, J.G. A review of the pathways of human exposure to poly- and perfluoroalkyl substances (PFASs) and present understanding of health effects. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 131–147. [Google Scholar] [CrossRef]
- Haschek, W.M.; Rousseaux, C.G.; Wallig, M.A. Toxicologic Pathology: An Introduction. In Haschek and Rousseaux’s Handbook of Toxicologic Pathology, Third Edition: Volume 1–3; Academic Press: Cambridge, MA, USA, 2013; Volume 1, pp. 1–9. ISBN 9780124157590. [Google Scholar]
- Friese, C.; Nuyts, N. Posthumanist critique and human health: How nonhumans (could) figure in public health research. Crit. Public Health 2017, 27, 303–313. [Google Scholar] [CrossRef]
- Falk, S.; Failing, K.; Georgii, S.; Brunn, H.; Stahl, T. Tissue specific uptake and elimination of perfluoroalkyl acids (PFAAs) in adult rainbow trout (Oncorhynchus mykiss) after dietary exposure. Chemosphere 2015, 129, 150–156. [Google Scholar] [CrossRef]
- Zhang, H.; Shen, L.; Fang, W.; Zhang, X.; Zhong, Y. Perfluorooctanoic acid-induced immunotoxicity via NF-kappa B pathway in zebrafish (Danio rerio) kidney. Fish Shellfish Immunol. 2021, 113, 9–19. [Google Scholar] [CrossRef]
- Manera, M.; Castaldelli, G.; Guerranti, C.; Giari, L. Effect of waterborne exposure to perfluorooctanoic acid on nephron and renal hemopoietic tissue of common carp Cyprinus carpio. Ecotoxicol. Environ. Saf. 2022, 234, 113407. [Google Scholar] [CrossRef]
- Manera, M.; Castaldelli, G.; Giari, L. Perfluorooctanoic acid affects thyroid follicles in common carp (Cyprinus carpio). Int. J. Environ. Res. Public Health 2022, 19, 9049. [Google Scholar] [CrossRef]
- Manera, M.; Casciano, F.; Giari, L. Ultrastructural alterations of the glomerular filtration barrier in fish experimentally exposed to perfluorooctanoic acid. Int. J. Environ. Res. Public Health 2023, 20, 5253. [Google Scholar] [CrossRef]
- Geven, E.J.W.; Klaren, P.H.M. The teleost head kidney: Integrating thyroid and immune signalling. Dev. Comp. Immunol. 2017, 66, 73–83. [Google Scholar] [CrossRef]
- Yang, J.-H.H. Perfluorooctanoic acid induces peroxisomal fatty acid oxidation and cytokine expression in the liver of male Japanese medaka (Oryzias latipes). Chemosphere 2010, 81, 548–552. [Google Scholar] [CrossRef]
- Du, G.; Huang, H.; Hu, J.; Qin, Y.; Wu, D.; Song, L.; Xia, Y.; Wang, X. Endocrine-related effects of perfluorooctanoic acid (PFOA) in zebrafish, H295R steroidogenesis and receptor reporter gene assays. Chemosphere 2013, 91, 1099–1106. [Google Scholar] [CrossRef]
- Chen, J.; Zheng, L.; Tian, L.; Wang, N.; Lei, L.; Wang, Y.; Dong, Q.; Huang, C.; Yang, D. Chronic PFOS exposure disrupts thyroid structure and function in Zebrafish. Bull. Environ. Contam. Toxicol. 2018, 101, 75–79. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Giari, L.; Guerranti, C.; Tognon, M.; Castaldelli, G.; Fano, E.A.; Martini, F. Environmental doses of perfluorooctanoic acid change the expression of genes in target tissues of common carp. Environ. Toxicol. Chem. 2018, 37, 942–948. [Google Scholar] [CrossRef]
- Olivares-Rubio, H.F.; Vega-López, A. Fatty acid metabolism in fish species as a biomarker for environmental monitoring. Environ. Pollut. 2016, 218, 297–312. [Google Scholar] [CrossRef]
- Godfrey, A.; Hooser, B.; Abdelmoneim, A.; Sepúlveda, M.S. Sex-specific endocrine-disrupting effects of three halogenated chemicals in Japanese medaka. J. Appl. Toxicol. 2019, 39, 1215–1223. [Google Scholar] [CrossRef]
- Ye, X.; Schoenfuss, H.L.; Jahns, N.D.; Delinsky, A.D.; Strynar, M.J.; Varns, J.; Nakayama, S.F.; Helfant, L.; Lindstrom, A.B. Perfluorinated compounds in common carp (Cyprinus carpio) fillets from the Upper Mississippi River. Environ. Int. 2008, 34, 932–938. [Google Scholar] [CrossRef]
- Jantzen, C.E.; Toor, F.; Annunziato, K.A.; Cooper, K.R. Effects of chronic perfluorooctanoic acid (PFOA) at low concentration on morphometrics, gene expression, and fecundity in zebrafish (Danio rerio). Reprod. Toxicol. 2017, 69, 34–42. [Google Scholar] [CrossRef]
- Dong, H.; Lu, G.; Yan, Z.; Liu, J.; Ji, Y. Molecular and phenotypic responses of male crucian carp (Carassius auratus) exposed to perfluorooctanoic acid. Sci. Total Environ. 2019, 653, 1395–1406. [Google Scholar] [CrossRef]
- Wang, L.; Wu, W.M.; Bolan, N.S.; Tsang, D.C.W.; Li, Y.; Qin, M.; Hou, D. Environmental fate, toxicity and risk management strategies of nanoplastics in the environment: Current status and future perspectives. J. Hazard. Mater. 2021, 401, 123415. [Google Scholar] [CrossRef]
- Dewitt, J.C.; Williams, W.C.; Creech, N.J.; Luebke, R.W. Suppression of antigen-specific antibody responses in mice exposed to perfluorooctanoic acid: Role of PPAR and T- and B-cell targeting. J. Immunotoxicol. 2016, 13, 38–45. [Google Scholar] [CrossRef]
- Chiu, W.; Braun, J.; Corsini, E.; Granum, B.; Keil, D.; Woolhiser, M. Immunotoxicity Associated with Exposure to Perfluorooctanoic Acid or Perfluorooctane Sulfonate. In National Toxicology Program Monograph; Research Triangle Park: Raleigh, NC, USA, 2016; pp. 1–147. [Google Scholar]
- Pecquet, A.M.; Maier, A.; Kasper, S.; Sumanas, S.; Yadav, J. Exposure to perfluorooctanoic acid (PFOA) decreases neutrophil migration response to injury in zebrafish embryos. BMC Res. Notes 2020, 13, 408. [Google Scholar] [CrossRef]
- Manera, M.; Dezfuli, B.S. Rodlet cells in teleosts: A new insight into their nature and functions. J. Fish Biol. 2004, 65, 597–619. [Google Scholar] [CrossRef]
- Sayyaf Dezfuli, B.; Pironi, F.; Maynard, B.; Simoni, E.; Bosi, G. Rodlet cells, fish immune cells and a sentinel of parasitic harm in teleost organs. Fish Shellfish Immunol. 2022, 121, 516–534. [Google Scholar] [CrossRef]
- Sayyaf Dezfuli, B.; Giari, L.; Bosi, G. Survival of Metazoan Parasites in Fish: Putting into Context the Protective Immune Responses of Teleost Fish. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2021; Volume 112, pp. 77–132. ISBN 9780323900836. [Google Scholar]
- Reite, O.B. Mast cells/eosinophilic granule cells of salmonids: Staining properties and responses to noxious agents. Fish Shellfish Immunol. 1997, 7, 567–584. [Google Scholar] [CrossRef]
- Reite, O.B. The rodlet cells of teleostean fish: Their potential role in host defence in relation to the role of mast cells/eosinophilic granule cells. Fish Shellfish Immunol. 2005, 19, 253–267. [Google Scholar] [CrossRef]
- Bosi, G.; DePasquale, J.A.; Manera, M.; Castaldelli, G.; Giari, L.; Sayyaf Dezfuli, B. Histochemical and immunohistochemical characterization of rodlet cells in the intestine of two teleosts, Anguilla anguilla and Cyprinus carpio. J. Fish Dis. 2018, 41, 475–485. [Google Scholar] [CrossRef]
- Sayyaf Dezfuli, B.; Castaldelli, G.; Lorenzoni, M.; Carosi, A.; Ovcharenko, M.; Bosi, G. Rodlet cells provide first line of defense against swimbladder nematode and intestinal coccidian in Anguilla anguilla. Fishes 2023, 8, 66. [Google Scholar] [CrossRef]
- Imagawa, T.; Hashimoto, Y.; Kon, Y.; Sugimura, M. Lectin histochemistry as special markers for rodlet cells in carp, Cyprinus carpio L. J. Fish Dis. 1990, 13, 537–540. [Google Scholar] [CrossRef]
- Smith, S.A.; Caceci, T.; Marei, H.E.; El-Habback, H.A. Observations on rodlet cells found in the vascular system and extravascular space of angelfish (Pterophyllum scalare scalare). J. Fish Biol. 1995, 46, 241–254. [Google Scholar] [CrossRef]
- Silphaduang, U.; Colorni, A.; Noga, E.J. Evidence for widespread distribution of piscidin antimicrobial peptides in teleost fish. Dis. Aquat. Organ. 2006, 72, 241–252. [Google Scholar] [CrossRef]
- Ronza, P.; Losada, A.P.; Villamarín, A.; Bermúdez, R.; Quiroga, M.I. Immunolocalization of tumor necrosis factor alpha in turbot (Scophthalmus maximus, L.) tissues. Fish Shellfish Immunol. 2015, 45, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Pergolizzi, S.; Capillo, G.; Lo Cascio, P.; Lauriano, E.R. Rodlet cells in kidney of goldfish (Carassius auratus, Linnaeus 1758): A light and confocal microscopy study. Acta Histochem. 2022, 124, 151876. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Bosi, G.; DePasquale, J.A.; Manera, M.; Giari, L. Fish innate immunity against intestinal helminths. Fish Shellfish Immunol. 2016, 50, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Leino, R.L. Ultrastructure of immature, developing, and secretory rodlet cells in fish. Cell Tissue Res. 1974, 155, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Laurà, R.; Germanà, G.P.; Levanti, M.B.; Guerrera, M.C.; Radaelli, G.; de Carlos, F.; Suárez, A.Á.; Ciriaco, E.; Germanà, A. Rodlet cells development in the intestine of sea bass (Dicentrarchus labrax). Microsc. Res. Tech. 2012, 75, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Kramer, C.R.; Potter, H. Ultrastructural observations on rodlet-cell development in the head kidney of the southern platyfish, Xiphophorus maculatus (Teleostei: Poeciliidae). Can. J. Zool. 2002, 80, 1422–1436. [Google Scholar] [CrossRef]
- Mazon, A.F.; Huising, M.O.; Taverne-Thiele, A.J.; Bastiaans, J.; Verburg-van Kemenade, B.M.L. The first appearance of Rodlet cells in carp (Cyprinus carpio L.) ontogeny and their possible roles during stress and parasite infection. Fish Shellfish Immunol. 2007, 22, 27–37. [Google Scholar] [CrossRef]
- Hawkins, W.E. Ultrastructure of rodlet cells: Response to cadmium damage in the kidney of the Spot Leiostomus xanthurus Lacépède. Gulf Res. Rep. 1984, 7, 365–372. [Google Scholar] [CrossRef]
- Iger, Y.; Abraham, M. Rodlet cells in the epidermis of fish exposed to stressors. Tissue Cell 1997, 29, 431–438. [Google Scholar] [CrossRef]
- Giari, L.; Manera, M.; Simoni, E.; Dezfuli, B.S. Cellular alterations in different organs of European sea bass Dicentrarchus labrax (L.) exposed to cadmium. Chemosphere 2007, 67, 1171–1181. [Google Scholar] [CrossRef]
- Giari, L.; Simoni, E.; Manera, M.; Dezfuli, B.S. Histo-cytological responses of Dicentrarchus labrax (L.) following mercury exposure. Ecotoxicol. Environ. Saf. 2008, 70, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Giari, L.; Dezfuli, B.S.; Lanzoni, M.; Castaldelli, G. The impact of an oil spill on organs of bream Abramis brama in the Po River. Ecotoxicol. Environ. Saf. 2012, 77, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Araujo, N.D.S.; Carlos, J.; Borges, S. Rodlet cells changes in Oreochromis niloticus in response to organophosphate pesticide and their relevance as stress biomarker in teleost fishes. Int. J. Aquat. Biol. 2015, 3, 398–408. [Google Scholar]
- Alesci, A.; Cicero, N.; Fumia, A.; Petrarca, C.; Mangifesta, R.; Nava, V.; Cascio, P.L.; Gangemi, S.; Di Gioacchino, M.; Lauriano, E.R. Histological and chemical analysis of heavy metals in kidney and gills of Boops boops: Melanomacrophages centers and rodlet cells as environmental biomarkers. Toxics 2022, 10, 218. [Google Scholar] [CrossRef]
- Vickers, T. A study of the intestinal epithelium of the Goldfish Carassius auratus: Its normal structure, the dynamics of cell replacement, and the changes induced by salts of cobalt and manganese. J. Cell Sci. 1962, 103, 93–110. [Google Scholar] [CrossRef]
- Giari, L.; Vincenzi, F.; Badini, S.; Guerranti, C.; Dezfuli, B.S.; Fano, E.A.; Castaldelli, G. Common carp Cyprinus carpio responses to sub-chronic exposure to perfluorooctanoic acid. Environ. Sci. Pollut. Res. 2016, 23, 15321–15330. [Google Scholar] [CrossRef]
- Loos, R.; Locoro, G.; Huber, T.; Wollgast, J.; Christoph, E.H.; de Jager, A.; Manfred Gawlik, B.; Hanke, G.; Umlauf, G.; Zaldívar, J.M. Analysis of perfluorooctanoate (PFOA) and other perfluorinated compounds (PFCs) in the River Po watershed in N-Italy. Chemosphere 2008, 71, 306–313. [Google Scholar] [CrossRef]
- Wei, Y.; Dai, J.; Liu, M.; Wang, J.; Xu, M.; Zha, J.; Wang, Z. Estrogen-like properties of perfluorooctanoic acid as revealed by expressing hepatic estrogen-responsive genes in rare minnows (Gobiocypris rarus). Environ. Toxicol. Chem. 2007, 26, 2440–2447. [Google Scholar] [CrossRef]
- Flood, M.T.; Nigrelli, R.F.; Gennaro, J.F. Some aspects of the infrastructure of the ‘Stäbchendrüscnzellen’, a peculiar cell associated with the endothelium of the Bulbus arteriosus and with other fish tissues. J. Fish Biol. 1975, 7, 129–138. [Google Scholar] [CrossRef]
- Van Es, B.; Klaassen, C.A.J.; Mnatsakanov, R.M. Estimating the structural distribution function of cell probabilities 1 The structural distribution function 2 A simulation. Austrian J. Stat. 2003, 32, 85–98. [Google Scholar] [CrossRef]
- Lindén, A.; Mäntyniemi, S. Using the negative binomial distribution to model overdispersion in ecological count data. Ecology 2011, 92, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- El-shaarawi, A.H. Negative Binomial Distribution. Encycl. Environmetrics 2012. [Google Scholar] [CrossRef]
- Yirga, A.A.; Melesse, S.F.; Mwambi, H.G.; Ayele, D.G. Negative binomial mixed models for analyzing longitudinal CD4 count data. Sci. Rep. 2020, 10, 16742. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Liu, C.; Preisser, J.S.; Wu, D. A bivariate zero-inflated negative binomial model and its applications to biomedical settings. Stat. Methods Med. Res. 2023, 32. [Google Scholar] [CrossRef] [PubMed]
- Bliss, C.I.; Fisher, R.A. Fitting the negative binomial distribution to biological data. Biometrics 1953, 9, 176. [Google Scholar] [CrossRef]
- Gurland, J. Some applications of the negative binomial and other contagious distributions. Am. J. Public Health 1959, 49, 1388–1399. [Google Scholar] [CrossRef]
- Hammami, I.; Garcia, A.; Nuel, G. Evidence for overdispersion in the distribution of malaria parasites and leukocytes in thick blood smears. Malar. J. 2013, 12, 398. [Google Scholar] [CrossRef]
- Hu, X.; Chi, Q.; Liu, Q.; Wang, D.; Zhang, Y.; Li, S. Atmospheric H2S triggers immune damage by activating the TLR-7/MyD88/NF-ΚB pathway and NLRP3 inflammasome in broiler thymus. Chemosphere 2019, 237, 124427. [Google Scholar] [CrossRef]
- DePasquale, J.A. Tropomyosin and alpha-actinin in teleost rodlet cells. Acta Zool. 2021, 323–332. [Google Scholar] [CrossRef]
- Vigliano, F.A.; Bermúdez, R.; Nieto, J.M.; Quiroga, M.I. Development of rodlet cells in the gut of turbot (Psetta maxima L.): Relationship between their morphology and S100 protein immunoreactivity. Fish Shellfish Immunol. 2009, 26, 146–153. [Google Scholar] [CrossRef]
- Leino, R.L. Rodlet cells in the gill and intestine of Catostomus commersoni and Perca flavescens: A comparison of their light and electron microscopic cytochemistry with that of mucous and granular cells. Can. J. Zool. 1982, 60, 2768–2782. [Google Scholar] [CrossRef]
- Ignarro, L.J. Nitric oxide. A novel signal transduction mechanism for transcellular communication. Hypertension 1990, 16, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Bielek, E. Development of the endoplasmic reticulum in the rodlet cell of two teleost species. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 2005, 283, 239–249. [Google Scholar] [CrossRef]
- Della Salda, L.; Manera, M.; Biavati, S. Ultrastructural features of associated rodlet cells in the renal epithelium of Sparus aurata L. J. Submicrosc. Cytol. Pathol. 1998, 30, 189–192. [Google Scholar]
- Mattey, D.L.; Morgan, M.; Wright, D.E. Distribution and development of rodlet cells in the gills and pseudobranch of the bass, Dicentrachus labrax (L). J. Fish Biol. 1979, 15, 363–370. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Giari, L.; Simoni, E.; Palazzi, D.; Manera, M. Alteration of rodlet cells in chub caused by the herbicide Stam® M-4 (Propanil). J. Fish Biol. 2003, 63, 232–239. [Google Scholar] [CrossRef]
- Giari, L.; Manera, M.; Simoni, E.; Dezfuli, B.S. Changes to chloride and rodlet cells in gills, kidney and intestine of Dicentrarchus labrax (L.) exposed to reduced salinities. J. Fish Biol. 2006, 69, 590–600. [Google Scholar] [CrossRef]
- Blank, U.; Madera-Salcedo, I.K.; Danelli, L.; Claver, J.; Tiwari, N.; Sánchez-Miranda, E.; Vázquez-Victorio, G.; Ramírez-Valadez, K.A.; Macias-Silva, M.; González-Espinosa, C. Vesicular trafficking and signaling for cytokine and chemokine secretion in mast cells. Front. Immunol. 2014, 5, 112088. [Google Scholar] [CrossRef]
- Klein, O.; Sagi-Eisenberg, R. Anaphylactic degranulation of mast cells: Focus on compound exocytosis. J. Immunol. Res. 2019, 2019, 9542656. [Google Scholar] [CrossRef]
- Crivellato, E.; Nico, B.; Mallardi, F.; Beltrami, C.A.; Ribatti, D. Piecemeal degranulation as a general secretory mechanism? Anat. Rec.Part A Discov. Mol. Cell. Evol. Biol. 2003, 274, 778–784. [Google Scholar] [CrossRef]
- Sandig, H.; Bulfone-Paus, S. TLR signaling in mast cells: Common and unique features. Front. Immunol. 2012, 3, 27433. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.S.K.; Lee, S.; Kim, H.H.; Choi, J.K.; Kim, S.H. Perfluorooctanoic acid induces mast cell-mediated allergic inflammation by the release of histamine and inflammatory mediators. Toxicol. Lett. 2012, 210, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, K.; Yoshino, S. Enhancement of Fce{open}RI-mediated degranulation response in the rat basophilic leukemia cell line RBL2H3 by the fluorosurfactants perfluorooctanoic acid and perfluorooctane sulfonate. Environ. Toxicol. Pharmacol. 2010, 29, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Sim, K.H.; Shrestha, P.; Yang, J.H.; Lee, Y.J. Perfluorooctane sulfonate and bisphenol A induce a similar level of mast cell activation via a common signaling pathway, Fyn-Lyn-Syk activation. Food Chem. Toxicol. 2021, 156, 112478. [Google Scholar] [CrossRef]
- Chang, H.W.; Kanegasaki, S.; Jin, F.; Deng, Y.; You, Z.; Chang, J.H.; Kim, D.Y.; Timilshina, M.; Kim, J.R.; Lee, Y.J.; et al. A common signaling pathway leading to degranulation in mast cells and its regulation by CCR1-ligand. Allergy Eur. J. Allergy Clin. Immunol. 2020, 75, 1371–1381. [Google Scholar] [CrossRef]
- Li, X.; Kanegasaki, S.; Jin, F.; Deng, Y.; Kim, J.R.; Chang, H.W.; Tsuchiya, T. Simultaneous induction of HSP70 expression, and degranulation, in IgE/Ag-stimulated or extracellular HSP70-stimulated mast cells. Allergy Eur. J. Allergy Clin. Immunol. 2018, 73, 361–368. [Google Scholar] [CrossRef]
- Siderits, D.; Bielek, E. Rodlet cells in the thymus of the zebrafish Danio rerio (Hamilton, 1822). Fish Shellfish Immunol. 2009, 27, 539–548. [Google Scholar] [CrossRef]
- Eguchi, Y.; Shimizu, S.; Tsujimoto, Y. Intracellular ATP levels determine cell death fate by apoptosis or necrosis. Cancer Res. 1997, 57, 1835–1840. [Google Scholar]
- Leist, M.; Single, B.; Castoldi, A.F.; Kühnle, S.; Nicotera, P. Intracellular adenosine triphosphate (ATP) concentration: A switch in the decision between apoptosis and necrosis. J. Exp. Med. 1997, 185, 1481–1486. [Google Scholar] [CrossRef]
- Gardel, M.L.; Kasza, K.E.; Brangwynne, C.P.; Liu, J.; Weitz, D.A. Mechanical Response of Cytoskeletal Networks. Methods Cell Biol. 2008, 89, 487–519. [Google Scholar]
- Baggiolini, M.; Horisberger, U.; Martin, U. Phagocytosis of mast cell granules by mononuclear phagocytes, neutrophils and eosinophils during anaphylaxis. Int. Arch. Allergy Immunol. 1982, 67, 219–226. [Google Scholar] [CrossRef]
- Welsh, R.A.; Geer, J.C. Phagocytosis of mast cell granule by the eosinophilic leukocyte in the rat. Am. J. Pathol. 1959, 35, 103–111. [Google Scholar] [PubMed]
- Miyata, K.; Takaya, K. Uptake of released mast cell granules by reticular cells of the rat lymph node. Cell Tissue Res. 1985, 240, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Subba Rao, P.V.; Friedman, M.M.; Atkins, F.M.; Metcalfe, D.D. Phagocytosis of mast cell granules by cultured fibroblasts. J. Immunol. 1983, 130, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Imagawa, T.; Kitagawa, H.; Uehara, M. An association between rodlet cells and the vascular endothelial cells in the head kidney of carp, Cyprinus carpio L.: Ultrastructural observations. J. Fish Dis. 1998, 21, 153–157. [Google Scholar] [CrossRef]
- Grunberg, W.; Hager, G. Ultrastructural and histochemical aspects of the rodlet cells from the bulbus arteriosus of Cyprinus carpio L.(Pisces: Cyprinidae)(author’s transl). Anat. Anz. 1978, 143, 277–290. [Google Scholar]
- Manera, M.; Simoni, E.; Dezfuli, B.S. The effect of dexamethasone on the occurrence and ultrastructure of rodlet cells in goldfish. J. Fish Biol. 2001, 59, 1239–1248. [Google Scholar] [CrossRef]
- Kim, H.R.; Park, J.S.; Soh, W.C.; Kim, N.Y.; Moon, H.Y.; Lee, J.S.; Jun, C.D. T cell microvilli: Finger-shaped external structures linked to the fate of T cells. Immune Netw. 2023, 23, e3. [Google Scholar] [CrossRef]
- Orbach, R.; Su, X. Surfing on membrane waves: Microvilli, curved membranes, and immune signaling. Front. Immunol. 2020, 11, 567890. [Google Scholar] [CrossRef]
- Fishelson, L.; Becker, K. Rodlet cells in the head and trunk kidney of the domestic carp (Cyprinus carpio): Enigmatic gland cells or coccidian parasites. Naturwissenschaften 1999, 86, 400–403. [Google Scholar] [CrossRef]
- Manera, M.; Castaldelli, G.; Fano, E.A.; Giari, L. Perfluorooctanoic acid-induced cellular and subcellular alterations in fish hepatocytes. Environ. Toxicol. Pharmacol. 2021, 81, 103548. [Google Scholar] [CrossRef] [PubMed]
- Manera, M.; Giari, L.; Vincenzi, F.; Guerranti, C.; DePasquale, J.A.; Castaldelli, G. Texture analysis in liver of common carp (Cyprinus carpio) sub-chronically exposed to perfluorooctanoic acid. Ecol. Indic. 2017, 81, 54–64. [Google Scholar] [CrossRef]
- National Research Council. Biologic Markers in Reproductive Toxicology; National Academy Press: Washington, DC, USA, 1989; ISBN 0309568145. [Google Scholar]
- World Health Orgnization. Biomarkers and Risk Assessment: Concepts and Principles; Environmental Health Criteria 155; World Health Organization: Geneva, Switzerland, 1993; ISBN 9241571551. [Google Scholar]
- Health and Ecological Criteria Division—Office of Science and Technology—Office of Water. Interim Drinking Water Health Advisory: Perfluorooctanoic Acid (PFOA); CASRN 335-67-1; U.S. Environmental Protection Agency: Washington, DC, USA, 2022. [Google Scholar]
- Teymoorian, T.; Munoz, G.; Vo Duy, S.; Liu, J.; Sauvé, S. Tracking PFAS in drinking water: A review of analytical methods and worldwide occurrence trends in tap water and bottled water. ACS EST Water 2023, 3, 246–261. [Google Scholar] [CrossRef]
- Rehman, A.U.; Crimi, M.; Andreescu, S. Current and emerging analytical techniques for the determination of PFAS in environmental samples. Trends Environ. Anal. Chem. 2023, 37, e00198. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n. of RCs Per Microscopic Field | Unexposed | 200 ng L−1 PFOA | 2 mg L−1 PFOA |
|---|---|---|---|
| 0 | 89 | 40 | 24 |
| 1 | 23 | 13 | 10 |
| 2 | 16 | 13 | 14 |
| 3 | 9 | 16 | 6 |
| 4 | 5 | 10 | 7 |
| 5 | 1 | 12 | 13 |
| 6 | 1 | 5 | 13 |
| 7 | 0 | 7 | 13 |
| 8 | 1 | 9 | 7 |
| 9 | 0 | 1 | 2 |
| 10 | 0 | 4 | 6 |
| 11 | 0 | 5 | 6 |
| 12 | 0 | 1 | 4 |
| 13 | 0 | 3 | 4 |
| 14 | 0 | 1 | 2 |
| 15 | 0 | 2 | 1 |
| 16 | 0 | 0 | 4 |
| 17 | 0 | 0 | 0 |
| 18 | 0 | 0 | 0 |
| 19 | 0 | 0 | 1 |
| 20 | 0 | 0 | 1 |
| 21 | 0 | 0 | 0 |
| 22 | 0 | 0 | 1 |
| 23 | 0 | 1 | 1 |
| 24 | 0 | 0 | 2 |
| 25 | 0 | 0 | 1 |
| 26 | 0 | 0 | 1 |
| Statistics | p | |
|---|---|---|
| Unexposed vs. 200 ng L−1 PFOA | 2.40 | 0.020 |
| Unexposed vs. 2 mg L−1 PFOA | 4.80 | <0.001 |
| 200 ng L−1 PFOA vs. 2 mg L−1 PFOA | 2.40 | 0.020 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manera, M.; Castaldelli, G.; Giari, L. Perfluorooctanoic Acid Promotes Recruitment and Exocytosis of Rodlet Cells in the Renal Hematopoietic Tissue of Common Carp. Toxics 2023, 11, 831. https://doi.org/10.3390/toxics11100831
Manera M, Castaldelli G, Giari L. Perfluorooctanoic Acid Promotes Recruitment and Exocytosis of Rodlet Cells in the Renal Hematopoietic Tissue of Common Carp. Toxics. 2023; 11(10):831. https://doi.org/10.3390/toxics11100831
Chicago/Turabian StyleManera, Maurizio, Giuseppe Castaldelli, and Luisa Giari. 2023. "Perfluorooctanoic Acid Promotes Recruitment and Exocytosis of Rodlet Cells in the Renal Hematopoietic Tissue of Common Carp" Toxics 11, no. 10: 831. https://doi.org/10.3390/toxics11100831
APA StyleManera, M., Castaldelli, G., & Giari, L. (2023). Perfluorooctanoic Acid Promotes Recruitment and Exocytosis of Rodlet Cells in the Renal Hematopoietic Tissue of Common Carp. Toxics, 11(10), 831. https://doi.org/10.3390/toxics11100831