
Expression Profiling of Adipogenic and Anti-Adipogenic MicroRNA Sequences following Methylmercury Exposure in Caenorhabditis elegans


Department of Pharmaceutical Sciences, Husson University School of Pharmacy, Bangor, ME 04401, USA
*
Author to whom correspondence should be addressed.
Toxics 2023, 11(11), 934; https://doi.org/10.3390/toxics11110934
Submission received: 12 October 2023
/
Revised: 8 November 2023
/
Accepted: 15 November 2023
/
Published: 17 November 2023
(This article belongs to the Special Issue Dietary Exposure to Heavy Metals and Health Risks)
Abstract
:MicroRNA (miRNA) are important regulators of gene expression that respond not only to developmental and pathological cues, but also to environmental stimuli. Dyslipidemia is a hallmark of metabolic conditions and has been shown to significantly affect the expression of circulating miRNA sequences. Recently, our lab has shown that the environmental toxicant methylmercury (MeHg) causes dyslipidemia in the Caenorhabditis elegans model organism. While 10 and 20 μM MeHg increases the expression of adipogenic transcription factors and lipid-binding proteins in worms, there is limited information on how the toxicant affects the miRNA regulators of these genes. We hypothesized that MeHg would increase the expression of adipogenic miRNA sequences and/or decrease the expression of anti-adipogenic miRNA sequences. We further hypothesized that the target mRNA sequences for the miRNAs affected by MeHg would be consequently altered. We selected three potentially adipogenic (mir-34, mir-124, and mir-355) and three potentially anti-adipogenic (mir-240, mir-786, and let-7) miRNA sequences homologous to known human miRNA sequences altered in obesity, and quantified their levels 24 h and 48 h post MeHg treatment. At 24 h post exposure, MeHg significantly increased expression of both the adipogenic and anti-adipogenic miRNA sequences 1.5–3x above untreated control. By 48 h post exposure, only the adipogenic miRNA sequences were elevated, while the anti-adipogenic miRNA sequences were decreased by 50% compared to untreated control. These data suggest that there are developmental changes in miRNA expression over time following MeHg exposure. We next selected one target mRNA sequence for each miRNA sequence based on miRNA–mRNA relationships observed in humans. MeHg altered the gene expression of all the target genes assayed. Except for mir-34, all the tested miRNA–mRNA sequences showed a conserved relationship between nematode and humans. To determine whether the selected miRNA sequences were involved in lipid accumulation in response to MeHg, lipid storage was investigated in transgenic worm strains that lacked the specific miRNA strains. Of the six strains investigated, only the mir-124 and let-7 mutant worms had lipid storage levels that were statistically different from wild type, suggesting that these two sequences can be potential mediators of MeHg-induced lipid dysregulation.
1. Introduction
Dyslipidemia is a condition where serum lipid levels of cholesterol, low-density lipoprotein (LDL), high-density lipoprotein (HDL), and triglycerides become imbalanced. Imbalance of serum lipids can occur due to a variety of means, such as over production of cholesterol, elevated levels of LDL or decreased levels of HDL. Dyslipidemia can arise through a variety of sources, including genetics, diet, or other environmental factors, and can predispose individuals to cardiovascular disease and metabolic syndrome (MS). MS is defined as a multifactorial condition characterized by obesity, insulin resistance, diabetes mellitus (DM), and dyslipidemia. Environmental factors have recently been implicated in the development of MS [1,2].
Methylmercury (MeHg) is a well-known neurotoxin that also has metabolic effects. Inorganic Hg is released through volcanic eruption as well as by human-driven processes, such as mining, industrial production and waste, as well as artisanal gold processing [3]. This inorganic Hg is biomethylated to MeHg by microbes. MeHg bioaccumulates and biomagnifies up the food chain, where humans are exposed to it via fish and seafood consumption. The highest concentrations of MeHg are present in large predatory species, such as tuna, mackerel, swordfish, and sharks [3]. Significant levels of MeHg have also been found in rice and poultry in China [4,5]. Developmental exposure to MeHg causes behavioral and cognitive dysfunction in children. Cumulative exposure to MeHg over an adult’s lifetime has been linked to movement disorders and the development of neurodegenerative diseases, such as Parkinson’s disease [3,6]. Increased blood and toenail mercury levels in individuals with high fish consumption have been associated with the development of MS, obesity, and lipid dysregulation [7,8,9], as well as increased visceral adipose tissue [10,11]. Furthermore, exposure to environmentally relevant concentrations of MeHg has been shown to increase lipid dysregulation and aggravate cardiovascular risk factors in wild-type and in ApoE knockout mice [12,13]. Elevations in cholesterol and triglycerides in response to MeHg have been observed in experimental animals as diverse as rodents, fish, and worms [14,15,16]. We have previously demonstrated that acute early-life exposure to 10 and 20 μM MeHg causes lipid dysregulation in Caenorhabditis elegans (C. elegans) by increasing the size and number of lipid storage sites, triglyceride levels, and altering gene expression for genes associated with lipid synthesis, transport, and storage [17]. The metabolic effects of MeHg in C. elegans were also shown to be modified by diet; the lower the lipid content of the diet, the less lipid dysregulation was observed in worms exposed to MeHg [18]. Additionally, lipid dysregulation could be modified by altering the ability of the worms to produce miRNAs. Worms that expressed more miRNA than wild type were protected from MeHg toxicity and lipid dysregulation, whereas worms that could not produce functional miRNA had exacerbated toxicity and lipid dysregulation in response to MeHg [19].
MiRNAs are short, non-coding RNAs comprised of 20–24 nucleotides in length. These RNA molecules work post-transcriptionally to repress the expression of target genes by base pairing with the 3′ UTR region of mRNA target sequences, leading to translational repression [20]. MiRNA are responsive to environmental, pharmacological, and pathological stimuli, and thereby are useful as biomarkers of disease and chemical exposure in humans [20]. MiRNA expression has been shown to be altered in serum and adipose tissue of patients with obesity, type 2 diabetes, and heart disease [20,21,22,23,24,25,26,27]. Some miRNAs act to increase lipid content of adipocytes or serum and are found in higher levels in serum of obese individuals than in non-obese individuals [21,22,23,24,25,26]. Other miRNA sequences are expressed highly in non-obese individuals but are lower in expression in obese individuals [21,22,23,24,25,26]. These miRNA sequences repress lipid accumulation pathways or increase lipid oxidation in organs. Characterization of the adipogenic potential of individual miRNA sequences has mainly been conducted in mammalian adipocyte cell culture, human patients, and rodents.
C. elegans can be a useful tool in screening miRNA sequences for their potential to cause lipid dysregulation in response to environmental factors, such as MeHg. C. elegans contains homologs for many of the miRNA that mammals express [27], and some miRNA that mammals express were first identified in C. elegans, such as let-7. C. elegans’ let-7 is a multifunctional miRNA that is highly conserved (80–100%) with the human let-7 family (let-7 a-k) [28,29]. In 3T3-L1 adipocytes, let-7 has been shown to inhibit adipogenesis mediated by PPARγ [30]. Let-7 is also a regulator of insulin resistance; let-7 knockout mice do not develop insulin resistance despite diet-induced obesity [31]. In C. elegans, let-7 has been shown to regulate lipid levels by modulating the mTORC2/PQM-1 signaling pathway in the intestine [32]. However, for many of the miRNA sequences with high sequence homology to human miRNA sequences, conservation of mRNA targets or physiological effects due to changes in their expression have not been characterized. Worms contain homologous machinery for the expression of miRNA as well as for lipid metabolic pathways. The ease of genetic manipulation allows for the generation of miRNA-deficient worm strains, which can be used to screen for physiological effects of MeHg or other environmental contaminants.
Recently we have described how miRNA expression can affect how C. elegans respond to MeHg. Worms that could not express functional miRNA were more sensitive to MeHg, had increased oxidative burden, triglyceride content, and lipid storage sites, as well as increased feeding behaviors [19]. These data suggested that MeHg exposure induced expression of miRNA that could modify these responses in wild-type worms but were not present in the miRNA-deficient strains. We therefore hypothesized that MeHg would increase the expression of the potentially adipogenic miRNA sequences and/or decrease the expression of the potentially anti-adipogenic miRNA sequences. We tested this hypothesis by quantifying three miRNA sequences shown in mammals to be adipogenic and three miRNA sequences shown in mammals to be anti-adipogenic at various time points following MeHg exposure in C. elegans. We assessed whether altered miRNA expression in response to MeHg leads to altered gene expression of known mRNA sequences for each miRNA. Finally, we assessed whether expression of any of the six miRNA sequences affected lipid accumulation in C. elegans basally and in response to MeHg.
2. Materials and Methods
2.1. Reagents
All reagents were obtained from Sigma-Aldrich (St. Louis, MO, USA), unless otherwise stated. Methylmercury (II) chloride, >99.5% purity (Sigma, catalog number 442534). Primers used in this study include aak-2 (F: TTGCTCACGAGTGGTTCCAG, R: CGCTGAAACTCAGTGACCTCT), atgl-1 (F: CCGACTACAAGTAAACGTGATGC, R: GGTGGCTGGGATGATGTGAA), daf-2 (F: GCTCTCGGAACAACCACTGA, R: TGAATCGGGCGTCGTTCTAC), lbp-5 (F: CGTGTGCTGCAAAACCAACT, R: CTCGTCGAATTCCACTCCCA), paqr-2 (F: TTCGTTGGCACTTTGCTTGG, R: ATAAACCAACATCCGCCGGT), and tba-1 (F: AGACCAACAAGCCGATGGAG, R: TCCAGTGCGGATCTCATCAAC). TaqManTM miRNA expression assays used in this study include mir-34-3p (241995_mat), mir-124-5p (243964_mat), mir-240-3p (001373_mat), mir-355-5p (244849_mat), mir-786-3p (461852_mat), let-7a-5p (4427975_mat), and U18 (001764).
2.2. C. elegans Strains and Handling of the Worms
C. elegans strains were maintained at 25 °C on Nematode Growth Medium (NGM) agar plates seeded with the OP-50 strain of Escherichia coli, as previously described [33]. The following strains were used in this study: wild-type N2, MT7626 (let-7(n2853) X), MT13406 (mir-34(n4276) X), MT15873 (mir-240(n4541) X), MT16316 (mir-355(n4618) II), MT18043 (mir-240 and mir-786(n4541) X), and VT2527 (mir-124(n4255) IV). All strains were obtained from the Caenorhabditis Genetic Center (CGC; University of Minnesota). Populations of worms were synchronized to the same developmental stage (L1) using a bleaching method [34]. Eggs were isolated from gravid worm populations using a bleach solution (1% NaOCl and 0.25 M NaOH) followed by a 30% sucrose gradient to separate eggs from worm and bacterial debris. L1 worms were then treated with various MeHg concentrations for 30 min in liquid M9 buffer at 25 °C on an Eppendorf tube rotator. For N2 worms, we have previously shown 1, 10, and 20 μM are below the lethal dose 50% (LD50) for MeHg in C. elegans and correlate to intracellular concentrations of MeHg that are below the US EPA reference dose of 0.1 μg/kg/d [35,36]. For experiments utilizing the transgenic miRNA strains, concentrations of MeHg were chosen below the LD50 for each individual strain.
2.3. RNA Isolation, miRNA Expression, and Real-Time qPCR Gene Expression
Twenty thousand worms were treated with MeHg, and RNA was isolated 24 or 48 h later using Trizol followed by chloroform extraction, as previously described [37]. After RNA extraction, miRNA was synthesized from 1 mg of total RNA using the TaqMan™ MicroRNA Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, MA, USA). Specific TaqMan® miRNA expression assays for the miRNA sequences were used according to manufacturer’s instructions. Alternatively, following RNA extraction, cDNA was synthesized from 1 mg of total RNA using the Applied Biosystems’ High-Capacity cDNA Reverse Transcription kit (Thermo Fisher Scientific). Real-time PCR analysis was performed using PerfeCTa SYBR Green FastMix (QuantaBio, Beverly, MA, USA). The housekeeping genes used to normalize miRNA data were U18 and tba-1 for mRNA.
2.4. Dose–Response Curves
The lethal dose (LD50) of MeHg for the six mutant C. elegans strains was determined by treating 5000 synchronized L1 worms with doses ranging from 0 to 200 μM MeHg. All exposures were performed in triplicate and repeated 5 times. After treatment, worms were washed 3 times with M9 buffer, then transferred to OP-50-seeded NGM plates, and manually counted for lethality after 24 h.
2.5. Nile Red Staining
We have previously shown that fat storage sites are increased by 10 and 20 μM MeHg in N2 worms as assessed by three different dyes: BODIPY 493/503, Oil Red O, and Nile Red [17]. Of the three methods, Nile Red is optimal for screening multiple genotypes and treatment groups. Following MeHg treatment, 20,000 L1 worms were washed with M9 and transferred to agar plates for 72 h. The worms were then washed off the agar plates and manually counted, placing 1000 worms per treatment in Eppendorf tubes for fixation. Fixation and Nile Red staining was performed as previously described [38]. Worms were first washed with 0.1% Triton X-100 in PBS, fixed in 40% isopropanol for 3 min, and then incubated with 3 μg/mL Nile Red in 40% isopropanol for 30 min. After staining, worms were washed with M9 and loaded onto a 96-well black microtiter plate. Nile Red fluorescence was read at excitation 560 nm and emission 590 nm. Data were normalized to worm number and protein levels.
2.6. Statistics
Statistical analyses were performed using Prism 8.4.3 software (Graphpad, San Diego, CA, USA). Data were tested for normal distribution using the Shapiro–Wilk test, all of which had p values indicating normality. Statistical analysis of significance was carried out either using Student’s t-test for LD50 value comparisons or two-way analysis of variance (ANOVA) for all other data. Values of p < 0.05 were considered statistically significant.
3. Results
3.1. MeHg Increases Adipogenic miRNA Expression and Decreases Anti-Adipogenic miRNA Expression
We have previously shown that genetically decreasing the expression of total miRNA significantly increased the toxicity of MeHg in C. elegans [19]. Therefore, we were interested in how individual miRNA sequences might change following MeHg exposure. We selected three nematode miRNA sequences with high homology to mammalian miRNA that have been shown to be increased and three sequences that have been shown to be decreased in the serum of obese individuals (Table 1). RNA was collected from MeHg-treated wild-type (N2) worms 24 h and 48 h post exposure, and miRNA expression was then quantified. After 24 h, MeHg significantly increased the levels of all six sequences above the level of untreated control (Figure 1). By 48 h post exposure, the adipogenic sequences mir-124 and mir-355 remained elevated by MeHg, while the anti-adipogenic miRNA sequences were decreased below the level of the untreated control (Figure 2). The adipogeinc mir-34 levels were elevated by MeHg exposure at 24 h, but were not significantly different from the untreated worms at 48 h. These data suggest that MeHg increases the expression of adipogenic mir-124 and mir-355 sequences for several hours post treatment in worms. Conversely, MeHg initially increased production of the anti-adipogenic let-7, mir-240, and mir-786 sequences, but the levels dropped over time post treatment. Both the increased adipogenic miRNA sequence levels and decreased anti-adipogenic miRNA sequence levels could lead to a gene expression profile that promotes dyslipidemia in response to MeHg.
3.2. MeHg Decreases the Expression of Anti-Adipogenic Genes Modulated by mir-124 and mir-355
To examine whether the increased levels of mir-34, mir-124, and mir-355 in response to MeHg would alter gene expression, we quantified gene expression of one target mRNA for each miRNA (Table 1). We selected the mRNA target genes based on the criteria that the gene has been shown to be regulated by miRNA in mammals and that there exists a well-characterized orthologous gene in nematodes. As MeHg increases the expression of mir-124 and mir-355, we hypothesized that the target sequences would decrease in expression in response to MeHg treatment. RNA was collected 24 and 48 h post MeHg exposure and target gene expression was quantified using real-time qPCR. At 24 h, aak-2, the target gene for mir-34, was increased, while atgl-1 and daf-2 (target genes for mir-124 and mir-355, respectively) showed no significant change in expression in response to MeHg as compared to untreated control (Figure 3A–C). In Figure 1, we show that mir-34 levels in response to MeHg were increased at 24 h; we therefore expected that aak-2 levels would decrease. However, MeHg increased aak-2 levels at 24 h (Figure 3A). Similarly at 48 h, MeHg increased the expression of aak-2, (Figure 4A) while mir-34 levels were unchanged by MeHg (Figure 2). Since aak-2 was actually increased by MeHg at both 24 and 48 h independent of mir-34 levels, our data suggest that either mir-34 does not regulate the aak-2 sequence in nematodes or that there are other regulators of aak-2 that are altered by MeHg. This suggests that other gene expression regulators are activated by MeHg that lead to an increase in aak-2 expression. At 48 h, atgl-1 and daf-2 were both decreased in response to MeHg (Figure 4B,C). The increased mir-124 and mir-355 expression and decreased atgl-1 and daf-2 expression confirmed the miRNA–mRNA relationship in nematodes and humans. Furthermore, it demonstrates how MeHg-induced changes in miRNA have consequences on mRNA expression.
3.3. MeHg Increases the Expression of Adipogenic Genes Modulated by let-7, mir-240, and mir-786
To examine whether the decreased expression levels of let-7, mir-240, and mir-786 in response to MeHg would alter gene expression, we selected one gene that has been shown, in humans, to be a target for each miRNA sequence (Table 1). As MeHg decreases the expression of let-7, mir-240, and mir-786, we hypothesized that the target sequences would increase in expression in response to MeHg treatment. RNA was collected 24 and 48 h post MeHg exposure and target gene expression was quantified using real-time qPCR. At 24 h, the expression of lbp-5 (target mRNA for let-7) and paqr-2 (target mRNA for both mir-786 and mir-240) levels were unchanged in response to MeHg treatment (Figure 3D,E). However, at 48 h, MeHg increased the expression of both lbp-5 and paqr-2 (Figure 4D,E). The decreased let-7, mir-240, and mir-786 expression and increased lbp-5 and paqr-2 expression confirmed the miRNA–mRNA relationship in nematodes and humans.
3.4. Reduced miRNA Expression Sensitizes Worms to MeHg Toxicity
We have previously shown that worms with altered miRNA expression capabilities have differential sensitivity to MeHg as compared to wild-type worms [19]. To examine the effect of deleting specific miRNA sequences on the worm’s sensitivity to MeHg, we performed a survival dose–response assay. We exposed wild-type N2 worms, mutant mir-34, mir-124, mir-355, mir-240, mir-240 and mir-786, and let-7 strains to increasing concentrations of MeHg, and generated dose–response survival curves (Figure 5). Table 2 presents the LD50 concentrations of MeHg for each strain. All mutant miRNA strains of worms showed significant left-hand shifts in the survival curves as compared to N2, indicating that deletion of the specific miRNA strain sensitizes worms to the toxic effects of MeHg. These data suggest that each of these miRNA may have additional roles in either development, xenobiotic metabolism, or stress responses that when lost, make the worms more susceptible to exposure to MeHg.
3.5. MiRNA Expression Influences MeHg-Induced Lipid Accumulation
Lastly, we examined whether deleting any of these miRNA sequences could affect lipid accumulation in response to MeHg. It was hypothesized that the deletion of either of the adipogenic miRNAs would prevent lipid accumulation in response to MeHg, while deletion of either of the anti-adipogenic miRNAs would augment lipid accumulation in response to MeHg. N2 and mutant L1 worms were treated with MeHg and lipid storage sites were stained with Nile Red in adult worms (72 h after exposure). As the mutant worms were more susceptible to MeHg than N2, MeHg concentrations were selected at each strain’s LD50 or at each strain’s LD25. Interestingly, treatment of N2 worms with MeHg increased lipid accumulation at all concentrations (0.25, 0.5, 1, and 3 μM) (Figure 6A–F).
In examining lipid storage in the deletion mutants for the adipogenic miRNA sequence strains, we expected the untreated mutants to have similar or lower lipid storage levels to the N2 untreated worms. Surprisingly, the untreated mutant mir-34 and mutant mir-355 worms had significantly more lipid storage than the untreated N2 worms (Figure 6A,C). Furthermore, while both mutant mir-34 and mutant mir-355 worms treated with MeHg had elevated lipid storage, there was no statistical difference between the MeHg-treated N2 worms and the MeHg-treated mutant worms. There was also no difference between the untreated mutant worms and the MeHg-treated mutant worms. Combining these data suggests that the effects of MeHg on lipid storage were not dependent on the expression of mir-34 or mir-355. In contrast, untreated mir-124 mutant worms had similar lipid storage levels to untreated N2 worms (Figure 6B). Additionally, lipid storage levels were elevated in mir-124 mutant worms exposed to MeHg; however, they were significantly lower than N2 worms treated with MeHg (Figure 6B). This suggests that mir-124 may be required for MeHg-induced lipid dysregulation.
In examining lipid storage in deletion mutants for the anti-adipogenic miRNA sequence strains, we expected the untreated mutants to have higher lipid storage levels than untreated N2 worms. Indeed, all three mutant strains had elevated lipid storage in untreated worms as compared to untreated N2 (Figure 6D–F). Mutant mir-240 and the mir-240 and mir-786 worms treated with MeHg were not statistically different in terms of lipid storage level as compared to untreated mutant worms, nor were they different from the N2 worms treated with MeHg. These data suggest that neither MeHg-induced lipid dysregulation is independent of mir-240 or mir-786. This was not the case for let-7. Untreated mutant let-7 worms had significantly higher levels of lipid storage than the untreated N2 worms, and treatment with MeHg raised lipid storage levels significantly higher than the mutant let-7 untreated worms or the N2 MeHg-treated worms (Figure 6D). This suggests that let-7 may be an important regulator of MeHg-induced lipid dysregulation.
4. Discussion
MeHg is a neurotoxic agent that has recently been associated with metabolic effects. Due to evolutionarily conserved lipid metabolic pathways, C. elegans is a useful model organism to study the effects of metabolic changes induced by MeHg [44]. We have previously shown that MeHg increases levels of adipogenic transcription factors implicated in MS including cepb-1 (ortholog to human C/EBP), nhr-49 (ortholog to human peroxisome proliferator-activated receptor gamma, PPARγ), and sbp-1 (ortholog to human sterol response element binding protein-1, SREBP-1), as well as a number of other genes involved in lipid synthesis and transport [17,18]. Additionally, we have shown that lipid dysregulation in response to MeHg exposure can be modulated with miRNA expression [19]. MiRNA are small non-protein coding RNA that silence gene expression by targeting specific mRNA sequences for degradation, thus preventing their translation. Deletion of miRNA processing machinery or the RNA polymerases that synthesize miRNA significantly altered toxicity, lipid accumulation, and oxidative stress in response to MeHg in worms [19]. While our initial studies focused on the effects of altering global miRNA expression, we were therefore interested in whether MeHg altered specific miRNA strains important for regulating lipid homeostasis. Herein, we demonstrate that MeHg alters the expression of specific miRNAs, leading to altered levels of their target mRNA sequences, and that the deletion of miRNA has functional consequences for lipid accumulation in worms.
It has previously been reported that MeHg decreases the expression of specific miRNA sequences in multiple models, including in immortalized human embryonic neural progenitor cells, cortical neurons, and in C. elegans [45,46,47]. Additionally, Hu et al. have found miRNA sequences that are up-regulated with MeHg exposure in zebrafish [48]. It is important to note that we currently do not know the mechanism of how miRNA expression is altered by MeHg. There are multiple possibilities, from altering miRNA gene expression to miRNA processing, miRNA stability, or gene silencing functions. Further research on this mechanism of action is essential. To date, it is unknown whether MeHg alters the expression of miRNAs that regulate lipid homeostasis. We selected six miRNA sequences in C. elegans that have homology to miRNA sequences that have been shown in mammalian systems to be either increased or decreased in the serum, adipocytes, or liver of individuals with MS. This is the first study to address whether these C. elegans sequences have similar effects on lipid metabolism as those that have been established in mammals.
C. elegans’ mir-34 is highly conserved with human miR-34A, miR-34B, and miR-34C [27]. In diet-induced obese mice, miR-34a targets fatty acid metabolism and cholesterol biosynthesis in the liver by targeting the histone deacetylase SIRT1, which has inhibitory roles in these processes [49]. miR-34 is up-regulated in adipose tissue of obese mice and in 3T3-L1 adipocyte cells in vitro, and is involved in insulin resistance [50]. Additionally, miR-34 is overexpressed during differentiation processes involved in cytokine-mediated β-cell dysfunction in non-obese diabetic mice [51]. Known targets of miR-34 include SIRT1, uncoupling protein 1 (UCP1), PPARγ coactivator 1α (PGC-1α), and AMP-activated protein kinase (AMPK) [40,49,52]. C. elegans’ mir-124 is highly conserved with human miR-124 [27]. In mammals, miR-124 is involved in insulin release from β-cells, and has an adipogenic effect-targeting Dlx5 transcription factor that affects cell differentiation in 3T3-L1 adipocyte cells [53,54]. Furthermore, miR-124 is overexpressed in visceral adipose tissue of obese children [55]. Known targets of miR-124 include adipose triglyceride lipase (ATGL), preadipocyte factor 1 (Pref-1), and tribbles homolog 3 (TRB3, a modulator of AKT signaling [41,56,57]. Finally, mir-355 has been shown to regulate insulin signaling by targeting the insulin-like growth factor 1 receptor (IGF1R) [39]. Conversely, C. elegans’ mir-240 is highly conserved with human miR-193a and miR-193b [27]. In mammals, miR-193b regulates adiponectin production in white adipose tissue, as well as inducing myoblast differentiation into brown adipocytes [42]. Additionally, miR-193b is one of several miRNAs that gets down-regulated in high-fat diet-induced obesity in mice [58]. Known targets of miR-193b include AKT, cAMP-responsive element-binding protein (CREB), nuclear transcription factor γ (NF-γ), and adiponectin receptor [42,59]. C. elegans’ mir-786 is highly conserved with human miR-365 [27]. MiR-365 is an anti-adipogenic miRNA that clusters with miR-193 and regulates brown adipose tissue differentiation [43]. Known targets of mir-365 include Runx1t1 runt-related transcription factor 1 (Runx1t1), UCP1, PGC-1α, PPARα, and PPARγ [43]. As previously mentioned, let-7 is a multifunctional miRNA found to inhibit adipogenesis and regulate insulin resistance [30,31]. Known mRNA targets of let-7 include PPARγ, FABP, and AMPK [30,60].
Overall, we found that expression of miRNA sequences in response to MeHg varied with the time post exposure. All six miRNA sequences were increased in expression 24 h post MeHg exposure as compared to untreated control worms. At 48 h post MeHg exposure, however, there were differences in expression between the adipogenic and anti-adipogenic miRNA sequences in the worms. At 48 h post MeHg treatment, anti-adipogenic miRNA sequences (let-7, mir-240, and mir-786) were decreased in response to MeHg and the adipogenic sequences mir-124 and mir-34 were increased in response to MeHg as compared to the untreated control worms. At this time, it is unclear why miRNA expression varied significantly post MeHg exposure. It is known that expression of miRNA is a dynamic process that fluctuates following developmental and environmental signals. Sun et al. have found that the expression of mmu-miR-15a-5p, mmu-miR-125b-5p, and mmu-miR-132–5p fluctuated in expression following ischemic transient common carotid artery occlusion in Mongolian gerbils (Meriones unguiculatus), leading to altered expression of target genes TDP43, FUS/TLS, and Hsp70 in the hippocampus [61]. Similar fluctuations in miRNA expression were found after 2 months post birth in mouse hippocampus following neonatal hypoxia [62]. Time-dependent changes in miRNA levels have also been observed for tumorigenesis, infection, and adipogenesis [63,64,65]. Therefore, it should not be surprising that the miRNA levels fluctuated post MeHg exposure as the worms developed from L1 stage to adulthood.
In the examination of both miRNA expression and mRNA expression post MeHg exposure, we found that certain miRNA sequence–mRNA target relationships were well conserved between humans and C. elegans. These included mir-124 and atgl-1, let-7 and lbp-5, as well as mir-240 and mir-786 with paqr-2. These relationships followed the expected directionality, i.e., decreased expression of let-7 occurred with increased expression of its target gene lbp-5. Furthermore, the shape of the dose–response curves for the miRNA expression matched the shape of the dose–response curve for mRNA target gene expression, further strengthening the relationship between the two genes. These data suggests that functions of these miRNA are evolutionarily conserved. It is important to note that in our study, we focused only on one mRNA target per miRNA sequence. As there are multiple mRNA targets for each miRNA, it will be important to determine whether other known mRNA target genes are similarly regulated by mir-124, let-7, mir-240, and mir-786 in C. elegans. Furthermore, since the miRNA–mRNA relationship was not confirmed by mir-34 and aak-2, there is a possibility that other mRNA target genes may actually show the miRNA–target mRNA relationship in worms. Finally, our data suggest that the effects of MeHg on miRNA sequences in C. elegans may have functional consequences in mammalian systems.
Genetic strains lacking each of the miRNA strains evaluated in this study were used to determine whether the miRNA sequence plays an important role in lipid accumulation in worms in the presence or absence of MeHg. Worms that were deficient in mir-34, mir-124, mir-355, let-7, mir-24, or mir-240 and mir-786 all were more sensitive to MeHg than N2 worms; dose–response survival curves for all the mutant strains were shifted to the left of N2 worms. These data are in agreement with our previous findings that worms that are deficient in miRNA due to a mutation in either pash-1 (partner of Drosha) or nrde-2 (nuclear RNAi defective) are more sensitive to MeHg [19]. MiRNA have multiple roles in metabolism and stress responses, as well as in development. As both the deletion of adipogenic or anti-adipogenic miRNA sequences made the worms more sensitive to MeHg, it is hypothesized that their roles in survival are independent of their roles in lipid regulation. It is unclear at this time how deletion of these miRNA sequences cause the worms to be more sensitive to MeHg.
Due to the increased sensitivity of the miRNA mutant worms to MeHg, we used significantly lower concentrations of MeHg than we have previously used in N2 worms to investigate lipid accumulation. Herein, we show that concentrations of MeHg as low as 0.25 μM increased lipid storage in N2 worms as measured with Nile Red staining. It was expected that when an adipogenic miRNA sequence was mutated, MeHg-induced lipid accumulation would be attenuated. This finding was observed for mir-124 mutant worms in response to MeHg, but not for mir-34 or mir-355 mutant worms. This suggests that mir-124 induction in response to MeHg has functional consequences in lipid accumulation. Previous studies of mir-124 in C. elegans has implicated the role of miRNA in neuronal functions such as neural cell specification, long-range patterning, and sensory neuronal morphogenesis [66,67,68]; however, its role in lipid metabolism had not been explored. Our data show a conserved role for mir-124 in lipid accumulation in worms and mammals. We also expected that mutation of an anti-adipogenic miRNA sequence would augment MeHg-induced lipid accumulation. Mutation of let-7, mir-240, and mir-240 and mir-276 increased lipid accumulation independent of MeHg treatment, demonstrating their conserved role in regulating lipid accumulation. These data agree with previous reports of let-7 regulating lipid levels in C. elegans [32]. Only mutation in let-7 augmented lipid accumulation in response to MeHg, suggesting let-7 down-regulation in response to MeHg has functional consequences.
5. Conclusions
Altogether, this study demonstrates conserved roles in lipid accumulation and mRNA gene expression in humans for four of the six investigated miRNA sequences in C. elegans (the adipogenic mir-124 and the anti-adipogenic mir-240, mir-276, and let-7). Of these four sequences, only mir-124 and let-7 were responsive to MeHg treatment. Our study was limited to only six miRNA sequences and one target mRNA sequence for each miRNA. Since there are multiple mRNA targets per miRNA, there can be differential effects of MeHg on these sequences. Further profiling of miRNA and target mRNA sequences in response to MeHg exposure is warranted to have a better understanding of mechanisms of metabolic alterations of lipids by MeHg.
Author Contributions
Conceptualization, S.C.; methodology, G.G., T.N. and S.C.; software, S.C.; validation, T.N. and G.G.; formal analysis, S.C.; investigation, G.G. and T.N.; data curation, S.C.; writing—original draft preparation, S.C.; writing—review and editing, S.C., G.G. and T.N.; visualization, S.C.; supervision, S.C.; project administration, S.C.; funding acquisition, S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Husson University School of Pharmacy Research Grant and Husson University Research Fund Program.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chamorro-Garcia, R.; Blumberg, B. Current Research Approaches and Challenges in the Obesogen Field. Front. Endocrinol. 2019, 10, 167. [Google Scholar] [CrossRef]
- Kassotis, C.D.; Stapleton, H.M. Endocrine-Mediated Mechanisms of Metabolic Disruption and New Approaches to Examine the Public Health Threat. Front. Endocrinol. 2019, 10, 39. [Google Scholar] [CrossRef]
- Clarkson, T.W.; Magos, L. The toxicology of mercury and its chemical compounds. Crit. Rev. Toxicol. 2006, 36, 609–662. [Google Scholar] [CrossRef]
- Li, Y.; He, X.; Wang, Y.; Guan, J.; Guo, J.; Xu, B.; Chen, Y.H.; Wang, G. Organic fertilizer amendment increases methylmercury accumulation in rice plants. Chemosphere 2020, 249, 126166. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Zhang, W.; Sun, G.; Feng, Z.; Hurley, J.P.; Yang, L.; Shang, L.; Feng, X. Mercury risk in poultry in the Wanshan Mercury Mine, China. Environ. Pollut. 2017, 230, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Landrigan, P.J.; Sonawane, B.; Butler, R.N.; Trasande, L.; Callan, R.; Droller, D. Early environmental origins of neurodegenerative disease in later life. Environ. Health Perspect. 2005, 113, 1230–1233. [Google Scholar] [CrossRef] [PubMed]
- Bulka, C.M.; Persky, V.W.; Daviglus, M.L.; Durazo-Arvizu, R.A.; Argos, M. Multiple metal exposures and metabolic syndrome: A cross-sectional analysis of the National Health and Nutrition Examination Survey 2011–2014. Environ. Res. 2019, 168, 397–405. [Google Scholar] [CrossRef]
- Wang, X.; Mukherjee, B.; Park, S.K. Associations of cumulative exposure to heavy metal mixtures with obesity and its comorbidities among U.S. adults in NHANES 2003–2014. Environ. Int. 2018, 121, 683–694. [Google Scholar] [CrossRef]
- Lee, K. Blood mercury concentration in relation to metabolic and weight phenotypes using the KNHANES 2011–2013 data. Int. Arch. Occup. Environ. Health 2018, 91, 185–193. [Google Scholar] [CrossRef]
- Park, J.S.; Ha, K.H.; He, K.; Kim, D.J. Association between Blood Mercury Level and Visceral Adiposity in Adults. Diabetes Metab. J. 2017, 41, 113–120. [Google Scholar] [CrossRef]
- Park, K.; Seo, E. Association between Toenail Mercury and Metabolic Syndrome Is Modified by Selenium. Nutrients 2016, 8, 424. [Google Scholar] [CrossRef] [PubMed]
- Lacerda Leocadio, P.C.; Dias, R.P.; Pinto, D.V.; Reis, J.M.; Rodrigues Nascimento, J.C.; Anne de Castro Brito, G.; Valenca, J.T., Jr.; Foureaux, G.; Ferreira, A.J.; Windmoller, C.C.; et al. Pollutants and nutrition: Are methylmercury effects on blood pressure and lipoprotein profile comparable to high-fat diet in mice? Ecotoxicol. Environ. Saf. 2020, 204, 111036. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.L.; Leocadio, P.C.L.; Reis, J.M.; Campos, G.P.; Capettini, L.S.A.; Foureaux, G.; Ferreira, A.J.; Windmoller, C.C.; Santos, F.A.; Oria, R.B.; et al. Oral methylmercury intoxication aggravates cardiovascular risk factors and accelerates atherosclerosis lesion development in ApoE knockout and C57BL/6 mice. Toxicol. Res. 2021, 37, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Guaraldi, G.; Lonardo, A.; Maia, L.; Palella, F.J., Jr. Metabolic concerns in aging HIV-infected persons: From serum lipid phenotype to fatty liver. AIDS 2017, 31, S147–S156. [Google Scholar] [CrossRef] [PubMed]
- Alam, R.T.M.; Abu Zeid, E.H.; Khalifa, B.A.; Arisha, A.H.; Reda, R.M. Dietary exposure to methyl mercury chloride induces alterations in hematology, biochemical parameters, and mRNA expression of antioxidant enzymes and metallothionein in Nile tilapia. Environ. Sci. Pollut. Res. Int. 2021, 28, 31391–31402. [Google Scholar] [CrossRef] [PubMed]
- Dutta, H.M.; Haghighi, A.Z. Methylmercuric chloride and serum cholesterol level in the bluegill (Lepomis macrochirus). Bull. Environ. Contam. Toxicol. 1986, 36, 181–185. [Google Scholar] [CrossRef]
- Caito, S.W.; Newell-Caito, J.; Martell, M.; Crawford, N.; Aschner, M. Methylmercury Induces Metabolic Alterations in Caenorhabditis elegans: Role for C/EBP Transcription Factor. Toxicol. Sci. 2020, 174, 112–123. [Google Scholar] [CrossRef]
- Crawford, N.; Martell, M.; Nielsen, T.; Khalil, B.; Imtiaz, F.; Nguidjo, E.; Newell-Caito, J.L.; Bornhorst, J.; Schwerdtle, T.; Caito, S.W. Methylmercury-Induced Metabolic Alterations in Caenorhabditis elegans Are Diet-Dependent. Toxics 2021, 9, 287. [Google Scholar] [CrossRef]
- Nielsen, T.; Crawford, N.; Martell, M.; Khalil, B.; Imtiaz, F.; Newell-Caito, J.L.; Caito, S. MicroRNA Expression Influences Methylmercury-Induced Lipid Accumulation and Mitochondrial Toxicity in Caenorhabditis elegans. Chem. Res. Toxicol. 2022, 35, 77–88. [Google Scholar] [CrossRef]
- Iacomino, G.; Siani, A. Role of microRNAs in obesity and obesity-related diseases. Genes. Nutr. 2017, 12, 23. [Google Scholar] [CrossRef]
- Zampetaki, A.; Kiechl, S.; Drozdov, I.; Willeit, P.; Mayr, U.; Prokopi, M.; Mayr, A.; Weger, S.; Oberhollenzer, F.; Bonora, E.; et al. Plasma microRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ. Res. 2010, 107, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Guay, C.; Regazzi, R. Circulating microRNAs as novel biomarkers for diabetes mellitus. Nat. Rev. Endocrinol. 2013, 9, 513–521. [Google Scholar] [CrossRef]
- Karolina, D.S.; Tavintharan, S.; Armugam, A.; Sepramaniam, S.; Pek, S.L.; Wong, M.T.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. Circulating miRNA profiles in patients with metabolic syndrome. J. Clin. Endocrinol. Metab. 2012, 97, E2271-6. [Google Scholar] [CrossRef]
- Pan, C.H.; Chien, S.C.; Chen, C.J.; Shih, C.M.; Hsieh, M.H.; Huang, C.Y.; Bi, W.F.; Chan, C.S.; Kao, Y.T.; Hsiao, C.Y.; et al. Circulating level of microRNA-142-5p is a potential biomarker for predicting in-stent restenosis: A case-control study. BMC Cardiovasc. Disord. 2021, 21, 77. [Google Scholar] [CrossRef]
- Marsetti, P.S.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A.; Lorente-Cebrian, S. Changes in miRNA expression with two weight-loss dietary strategies in a population with metabolic syndrome. Nutrition 2021, 83, 111085. [Google Scholar] [CrossRef]
- Heneghan, H.M.; Miller, N.; McAnena, O.J.; O’Brien, T.; Kerin, M.J. Differential miRNA expression in omental adipose tissue and in the circulation of obese patients identifies novel metabolic biomarkers. J. Clin. Endocrinol. Metab. 2011, 96, E846–E850. [Google Scholar] [CrossRef]
- Ibanez-Ventoso, C.; Vora, M.; Driscoll, M. Sequence relationships among C. elegans, D. melanogaster and human microRNAs highlight the extensive conservation of microRNAs in biology. PLoS ONE 2008, 3, e2818. [Google Scholar] [CrossRef]
- Lee, H.; Han, S.; Kwon, C.S.; Lee, D. Biogenesis and regulation of the let-7 miRNAs and their functional implications. Protein Cell 2016, 7, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Muller, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef]
- Sun, T.; Fu, M.; Bookout, A.L.; Kliewer, S.A.; Mangelsdorf, D.J. MicroRNA let-7 regulates 3T3-L1 adipogenesis. Mol. Endocrinol. 2009, 23, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.J.; Olson, E.N. Control of glucose homeostasis and insulin sensitivity by the Let-7 family of microRNAs. Proc. Natl. Acad. Sci. USA 2011, 108, 21075–21080. [Google Scholar] [CrossRef]
- Dowen, R.H.; Breen, P.C.; Tullius, T.; Conery, A.L.; Ruvkun, G. A microRNA program in the C. elegans hypodermis couples to intestinal mTORC2/PQM-1 signaling to modulate fat transport. Genes Dev. 2016, 30, 1515–1528. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans. In C. elegans: A. Practical. Approach; Hope, I.A., Ed.; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Caito, S.W.; Aschner, M. NAD+ Supplementation Attenuates Methylmercury Dopaminergic and Mitochondrial Toxicity in Caenorhabditis Elegans. Toxicol. Sci. 2016, 151, 139–149. [Google Scholar] [CrossRef]
- Martinez-Finley, E.J.; Chakraborty, S.; Slaughter, J.C.; Aschner, M. Early-life exposure to methylmercury in wildtype and pdr-1/parkin knockout C. elegans. Neurochem. Res. 2013, 38, 1543–1552. [Google Scholar] [CrossRef]
- Chomczynski, P.; Mackey, K. Short technical reports. Modification of the TRI reagent procedure for isolation of RNA from polysaccharide- and proteoglycan-rich sources. Biotechniques 1995, 19, 942–945. [Google Scholar]
- Pino, E.C.; Webster, C.M.; Carr, C.E.; Soukas, A.A. Biochemical and high throughput microscopic assessment of fat mass in Caenorhabditis elegans. J. Vis. Exp. 2013, 73, 50180. [Google Scholar]
- Zhi, L.; Yu, Y.; Jiang, Z.; Wang, D. mir-355 Functions as An Important Link between p38 MAPK Signaling and Insulin Signaling in the Regulation of Innate Immunity. Sci. Rep. 2017, 7, 14560. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; An, C.; Wu, X.; Yang, Y.; Xu, J.; Liu, Y.; Wang, C.; Nie, L.; Fang, H.; Yang, Z. MiR-34a regulates mitochondrial content and fat ectopic deposition induced by resistin through the AMPK/PPARalpha pathway in HepG2 cells. Int. J. Biochem. Cell Biol. 2018, 94, 133–145. [Google Scholar] [CrossRef]
- Fang, Q.H.; Shen, Q.L.; Li, J.J.; Yang, Y.; Guo, J.J.; Cheng, Y.; Zhou, H.C.; Niu, W.Y.; Chen, L.M.; Li, C.J.; et al. Inhibition of microRNA-124a attenuates non-alcoholic fatty liver disease through upregulation of adipose triglyceride lipase and the effect of liraglutide intervention. Hepatol. Res. 2019, 49, 743–757. [Google Scholar] [CrossRef]
- Belarbi, Y.; Mejhert, N.; Lorente-Cebrian, S.; Dahlman, I.; Arner, P.; Ryden, M.; Kulyte, A. MicroRNA-193b Controls Adiponectin Production in Human White Adipose Tissue. J. Clin. Endocrinol. Metab. 2015, 100, E1084-8. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xie, H.; Mori, M.A.; Alexander, R.; Yuan, B.; Hattangadi, S.M.; Liu, Q.; Kahn, C.R.; Lodish, H.F. Mir193b-365 is essential for brown fat differentiation. Nat. Cell Biol. 2011, 13, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.T.; Ashrafi, K. Caenorhabditis elegans as an emerging model for studying the basic biology of obesity. Dis. Models Mech. 2009, 2, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Rudgalvyte, M.; Peltonen, J.; Lakso, M.; Wong, G. Chronic MeHg exposure modifies the histone H3K4me3 epigenetic landscape in Caenorhabditis elegans. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 191, 109–116. [Google Scholar] [CrossRef]
- Guida, N.; Valsecchi, V.; Laudati, G.; Serani, A.; Mascolo, L.; Molinaro, P.; Montuori, P.; Di Renzo, G.; Canzoniero, L.M.; Formisano, L. The miR206-JunD Circuit Mediates the Neurotoxic Effect of Methylmercury in Cortical Neurons. Toxicol. Sci. 2018, 163, 569–578. [Google Scholar] [CrossRef]
- Wang, X.; Yan, M.; Zhao, L.; Wu, Q.; Wu, C.; Chang, X.; Zhou, Z. Low-Dose Methylmercury-Induced Genes Regulate Mitochondrial Biogenesis via miR-25 in Immortalized Human Embryonic Neural Progenitor Cells. Int. J. Mol. Sci. 2016, 17, 2058. [Google Scholar] [CrossRef]
- Hu, H.; Shi, Y.; Zhang, Y.; Wu, J.; Asweto, C.O.; Feng, L.; Yang, X.; Duan, J.; Sun, Z. Comprehensive gene and microRNA expression profiling on cardiovascular system in zebrafish co-exposured of SiNPs and MeHg. Sci. Total. Environ. 2017, 607–608, 795–805. [Google Scholar] [CrossRef]
- Lee, J.; Padhye, A.; Sharma, A.; Song, G.; Miao, J.; Mo, Y.Y.; Wang, L.; Kemper, J.K. A pathway involving farnesoid X receptor and small heterodimer partner positively regulates hepatic sirtuin 1 levels via microRNA-34a inhibition. J. Biol. Chem. 2010, 285, 12604–12611. [Google Scholar] [CrossRef]
- Cornejo, P.J.; Vergoni, B.; Ohanna, M.; Angot, B.; Gonzalez, T.; Jager, J.; Tanti, J.F.; Cormont, M. The Stress-Responsive microRNA-34a Alters Insulin Signaling and Actions in Adipocytes through Induction of the Tyrosine Phosphatase PTP1B. Cells 2022, 11, 2581. [Google Scholar] [CrossRef]
- Dumortier, O.; Hinault, C.; Van Obberghen, E. MicroRNAs and metabolism crosstalk in energy homeostasis. Cell Metab. 2013, 18, 312–324. [Google Scholar] [CrossRef]
- Fu, T.; Seok, S.; Choi, S.; Huang, Z.; Suino-Powell, K.; Xu, H.E.; Kemper, B.; Kemper, J.K. MicroRNA 34a inhibits beige and brown fat formation in obesity in part by suppressing adipocyte fibroblast growth factor 21 signaling and SIRT1 function. Mol. Cell Biol. 2014, 34, 4130–4142. [Google Scholar] [CrossRef] [PubMed]
- Qadir, A.S.; Woo, K.M.; Ryoo, H.M.; Baek, J.H. Insulin suppresses distal-less homeobox 5 expression through the up-regulation of microRNA-124 in 3T3-L1 cells. Exp. Cell Res. 2013, 319, 2125–2134. [Google Scholar] [CrossRef]
- Lovis, P.; Gattesco, S.; Regazzi, R. Regulation of the expression of components of the exocytotic machinery of insulin-secreting cells by microRNAs. Biol. Chem. 2008, 389, 305–312. [Google Scholar] [CrossRef]
- Roy, D.; Modi, A.; Ghosh, R.; Ghosh, R.; Benito-Leon, J. Visceral Adipose Tissue Molecular Networks and Regulatory microRNA in Pediatric Obesity: An In Silico Approach. Int. J. Mol. Sci. 2022, 23, 11036. [Google Scholar] [CrossRef]
- Wang, G.; Zou, H.; Lai, C.; Huang, X.; Yao, Y.; Xiang, G. Repression of MicroRNA-124-3p Alleviates High-Fat Diet-Induced Hepatosteatosis by Targeting Pref-1. Front. Endocrinol. 2020, 11, 589994. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, J.; Liu, Q.; Xiong, X.; Zhang, Z.; Jiao, Y.; Li, X.; Liu, B.; Li, Y.; Lu, Y. MicroRNA-124 promotes hepatic triglyceride accumulation through targeting tribbles homolog 3. Sci. Rep. 2016, 6, 37170. [Google Scholar] [CrossRef] [PubMed]
- Chartoumpekis, D.V.; Zaravinos, A.; Ziros, P.G.; Iskrenova, R.P.; Psyrogiannis, A.I.; Kyriazopoulou, V.E.; Habeos, I.G. Differential expression of microRNAs in adipose tissue after long-term high-fat diet-induced obesity in mice. PLoS ONE 2012, 7, e34872. [Google Scholar] [CrossRef]
- Kang, Z.; Zhang, S.; Jiang, E.; Wan, F.; Lan, X.; Liu, M. Mir-193b Regulates the Differentiation, Proliferation, and Apoptosis of Bovine Adipose Cells by Targeting the ACSS2/AKT Axis. Animals 2020, 10, 1265. [Google Scholar] [CrossRef]
- Simino, L.A.P.; Panzarin, C.; Fontana, M.F.; de Fante, T.; Geraldo, M.V.; Ignacio-Souza, L.M.; Milanski, M.; Torsoni, M.A.; Ross, M.G.; Desai, M.; et al. MicroRNA Let-7 targets AMPK and impairs hepatic lipid metabolism in offspring of maternal obese pregnancies. Sci. Rep. 2021, 11, 8980. [Google Scholar] [CrossRef]
- Sun, M.; Yamashita, T.; Shang, J.; Liu, N.; Deguchi, K.; Feng, J.; Abe, K. Time-dependent profiles of microRNA expression induced by ischemic preconditioning in the gerbil hippocampus. Cell Transplant. 2015, 24, 367–376. [Google Scholar] [CrossRef]
- Leavy, A.; Brennan, G.P.; Jimenez-Mateos, E.M. MicroRNA Profiling Shows a Time-Dependent Regulation within the First 2 Months Post-Birth and after Mild Neonatal Hypoxia in the Hippocampus from Mice. Biomedicines 2022, 10, 2740. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, S.B.; Ozturk, B.; Kocak, N.; Unlu, A. Differences of time-dependent microRNA expressions in breast cancer cells. Noncoding RNA Res. 2021, 6, 15–22. [Google Scholar] [CrossRef]
- Li, H.; Lin, L.; Chong, L.; Gu, S.; Wen, S.; Yu, G.; Hu, X.; Dong, L.; Zhang, H.; Li, C. Time-resolved mRNA and miRNA expression profiling reveals crucial coregulation of molecular pathways involved in epithelial-pneumococcal interactions. Immunol. Cell Biol. 2020, 98, 726–742. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.S.; Aneesh Kumar, A.; Abdul Jaleel, K.A. Time-dependent alterations in mRNA, protein and microRNA during in vitro adipogenesis. Mol. Cell Biochem. 2018, 448, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.M.; Goldstein, L.D.; Tevlin, M.; Tavare, S.; Shaham, S.; Miska, E.A. The microRNA miR-124 controls gene expression in the sensory nervous system of Caenorhabditis elegans. Nucleic Acids. Res. 2010, 38, 3780–3793. [Google Scholar] [CrossRef]
- Chen, J.S.; Gumbayan, A.M.; Zeller, R.W.; Mahaffy, J.M. An expanded Notch-Delta model exhibiting long-range patterning and incorporating MicroRNA regulation. PLoS. Comput. Biol. 2014, 10, e1003655. [Google Scholar] [CrossRef]
- Wulczyn, F.G.; Smirnova, L.; Rybak, A.; Brandt, C.; Kwidzinski, E.; Ninnemann, O.; Strehle, M.; Seiler, A.; Schumacher, S.; Nitsch, R. Post-transcriptional regulation of the let-7 microRNA during neural cell specification. FASEB. J. 2007, 21, 415–426. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Methylmercury increases miRNA expression 24 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 24 h later. Levels of adipogenic miRNA sequences (A) mir-34, (B) mir-124, (C) mir-355 and anti-adipogenic miRNA sequences (D) let-7, (E) mir-240, and (F) mir-786 were quantified using qPCR and normalized to U18 C/D box small nucleolar RNA as housekeeping gene. Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated control.
Figure 1.
Methylmercury increases miRNA expression 24 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 24 h later. Levels of adipogenic miRNA sequences (A) mir-34, (B) mir-124, (C) mir-355 and anti-adipogenic miRNA sequences (D) let-7, (E) mir-240, and (F) mir-786 were quantified using qPCR and normalized to U18 C/D box small nucleolar RNA as housekeeping gene. Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated control.
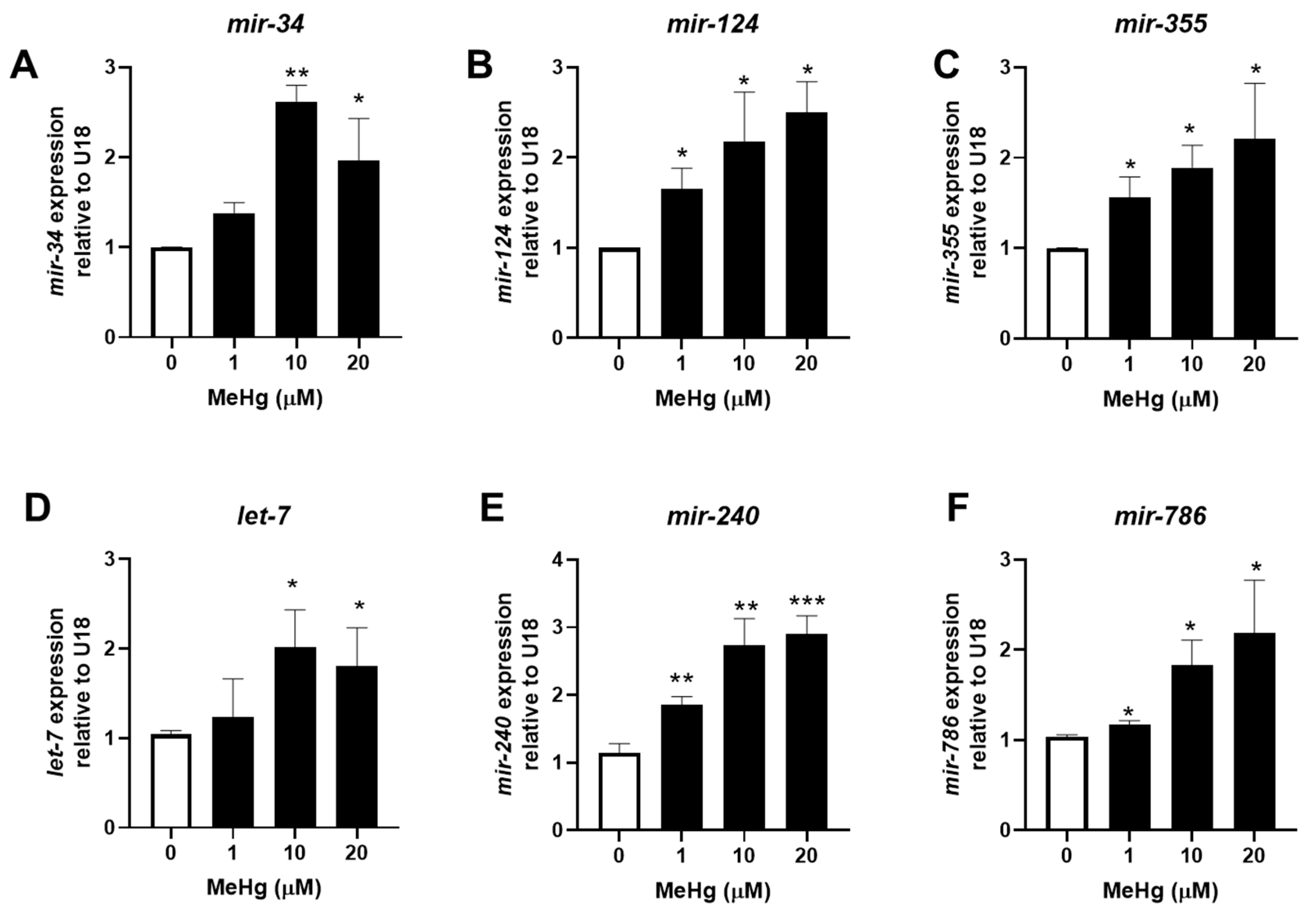
Figure 2.
Methylmercury increases adipogenic miRNA expression and decreases anti-adipogenic miRNA expression 48 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 48 h later. Levels of adipogenic miRNA sequences (A) mir-34, (B) mir-124, (C) mir-355 and anti-adipogenic miRNA sequences (D) let-7, (E) mir-240, and (F) mir-786 were quantified using qPCR and normalized to U18 C/D box small nucleolar RNA as housekeeping gene. Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated control.
Figure 2.
Methylmercury increases adipogenic miRNA expression and decreases anti-adipogenic miRNA expression 48 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 48 h later. Levels of adipogenic miRNA sequences (A) mir-34, (B) mir-124, (C) mir-355 and anti-adipogenic miRNA sequences (D) let-7, (E) mir-240, and (F) mir-786 were quantified using qPCR and normalized to U18 C/D box small nucleolar RNA as housekeeping gene. Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated control.

Figure 3.
MeHg alters aak-2 but none of the other mRNA sequences of miRNA-responsive genes 24 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 24 h later. Total RNA was extracted, cDNA was made, and used for qPCR. Expression of (A) aak-2, (B) atgl-1, (C) daf-2, (D) lbp-5, and (E) paqr-2 were compared relative to tubulin alpha chain 1 (tba-1). Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05 as compared with untreated control.
Figure 3.
MeHg alters aak-2 but none of the other mRNA sequences of miRNA-responsive genes 24 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 24 h later. Total RNA was extracted, cDNA was made, and used for qPCR. Expression of (A) aak-2, (B) atgl-1, (C) daf-2, (D) lbp-5, and (E) paqr-2 were compared relative to tubulin alpha chain 1 (tba-1). Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05 as compared with untreated control.

Figure 4.
MeHg alters mRNA expression of miRNA-responsive genes 48 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 48 h later. Total RNA was extracted, cDNA was made, and used for qPCR. Expression of (A) aak-2, (B) atgl-1, (C) daf-2 (D) lbp-5 and (E) paqr-2 were compared relative to tubulin alpha chain 1 (tba-1). Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated control.
Figure 4.
MeHg alters mRNA expression of miRNA-responsive genes 48 h post exposure. N2 worms were exposed to either 0, 1, 10, or 20 μM of MeHg for 30 min and harvested 48 h later. Total RNA was extracted, cDNA was made, and used for qPCR. Expression of (A) aak-2, (B) atgl-1, (C) daf-2 (D) lbp-5 and (E) paqr-2 were compared relative to tubulin alpha chain 1 (tba-1). Data are representative of the mean ± SEM from 6 independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated control.

Figure 5.
Knockout of specific miRNA sequences affects MeHg toxicity. N2 and transgenic worms with mutations in (A) mir-134, (B) mir-124, (C) mir-355, (D) let-7, (E) mir-240, and (F) mir-240 and mir-786 were treated with increasing concentrations of MeHg for 30 min and transferred to agar plates spread with OP50 E. coli. Worms were manually counted 24 h post exposure. Dose–response curves were generated from five independent experiments.
Figure 5.
Knockout of specific miRNA sequences affects MeHg toxicity. N2 and transgenic worms with mutations in (A) mir-134, (B) mir-124, (C) mir-355, (D) let-7, (E) mir-240, and (F) mir-240 and mir-786 were treated with increasing concentrations of MeHg for 30 min and transferred to agar plates spread with OP50 E. coli. Worms were manually counted 24 h post exposure. Dose–response curves were generated from five independent experiments.

Figure 6.
Lipid content in response to MeHg is affected by miRNA expression. Lipid storage sites were quantified from N2 and transgenic worms with mutations in (A) mir-134, (B) mir-124, (C) mir-355, (D) let-7, (E) mir-240, and (F) mir-240 and mir-786 treated with MeHg for 30 min. At 72 h after MeHg treatment, worms were fixed, stained with Nile Red, and fluorescence was measured. Data are expressed as mean Nile Red fluorescence normalized to worm number and protein content ± SEM from five independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated N2 control.
Figure 6.
Lipid content in response to MeHg is affected by miRNA expression. Lipid storage sites were quantified from N2 and transgenic worms with mutations in (A) mir-134, (B) mir-124, (C) mir-355, (D) let-7, (E) mir-240, and (F) mir-240 and mir-786 treated with MeHg for 30 min. At 72 h after MeHg treatment, worms were fixed, stained with Nile Red, and fluorescence was measured. Data are expressed as mean Nile Red fluorescence normalized to worm number and protein content ± SEM from five independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 as compared with untreated N2 control.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
miRNA and target mRNA sequences.
| miRNA | Effect | mRNA Target Gene (Mammalian Homolog) | References |
|---|---|---|---|
| mir-355 | Adipogenic | daf-2 (insulin-like growth factor receptor 1) | [39] |
| mir-34 | Adipogenic | aak-2 (AMP kinase) | [40] |
| mir-124 | Adipogenic | atgl-1 (adipose triglyceride lipase) | [41] |
| mir-240 | Anti-adipogenic | paqr-2 (adiponectin receptor) | [42] |
| mir-786 | Anti-adipogenic | paqr-2 | [43] |
| let-7 | Anti-adipogenic | lbp-5 (Fatty acid-binding protein 4) | [30] |
Table 2.
MeHg is more toxic in transgenic worms lacking specific miRNA sequences.
| Strain | LD50 (μM) | p-Value |
|---|---|---|
| N2 | 20.43 | |
| Mutant mir-34 | 2.53 | <0.0001 |
| Mutant mir-124 | 0.66 | <0.0001 |
| Mutant mir-355 | 1.43 | <0.0001 |
| Mutant let-7 | 3.95 | <0.0001 |
| Mutant mir-240 | 1.77 | <0.0001 |
| Mutant mir-240 and mir-786 | 1.66 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Garofalo, G.; Nielsen, T.; Caito, S. Expression Profiling of Adipogenic and Anti-Adipogenic MicroRNA Sequences following Methylmercury Exposure in Caenorhabditis elegans. Toxics 2023, 11, 934. https://doi.org/10.3390/toxics11110934
AMA Style
Garofalo G, Nielsen T, Caito S. Expression Profiling of Adipogenic and Anti-Adipogenic MicroRNA Sequences following Methylmercury Exposure in Caenorhabditis elegans. Toxics. 2023; 11(11):934. https://doi.org/10.3390/toxics11110934
Chicago/Turabian StyleGarofalo, Giancarlo, Tyson Nielsen, and Samuel Caito. 2023. "Expression Profiling of Adipogenic and Anti-Adipogenic MicroRNA Sequences following Methylmercury Exposure in Caenorhabditis elegans" Toxics 11, no. 11: 934. https://doi.org/10.3390/toxics11110934
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.