




Identification of the Key Genes Involved in Proline-Mediated Modification of Cell Wall Components in Rice Seedlings under Trivalent Chromium Exposure


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Treatments
2.2. Microarray Analysis
2.3. RT-qPCR Analysis
2.4. Calculation of Gene Expression Variation Factors (GEVFs)
2.5. Data Analysis
3. Results
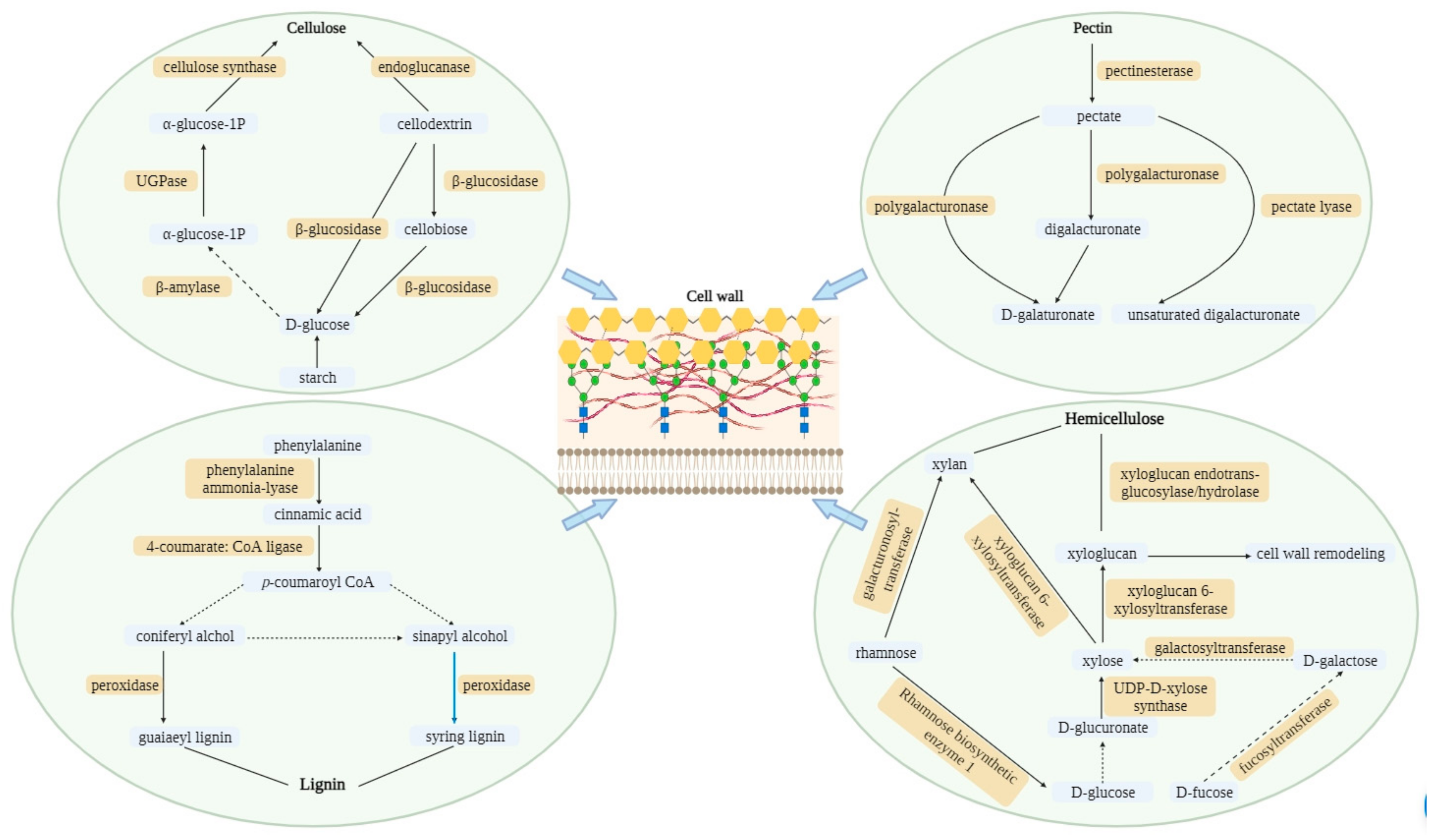
3.1. Identification of Genes Involved in Cell Wall Components
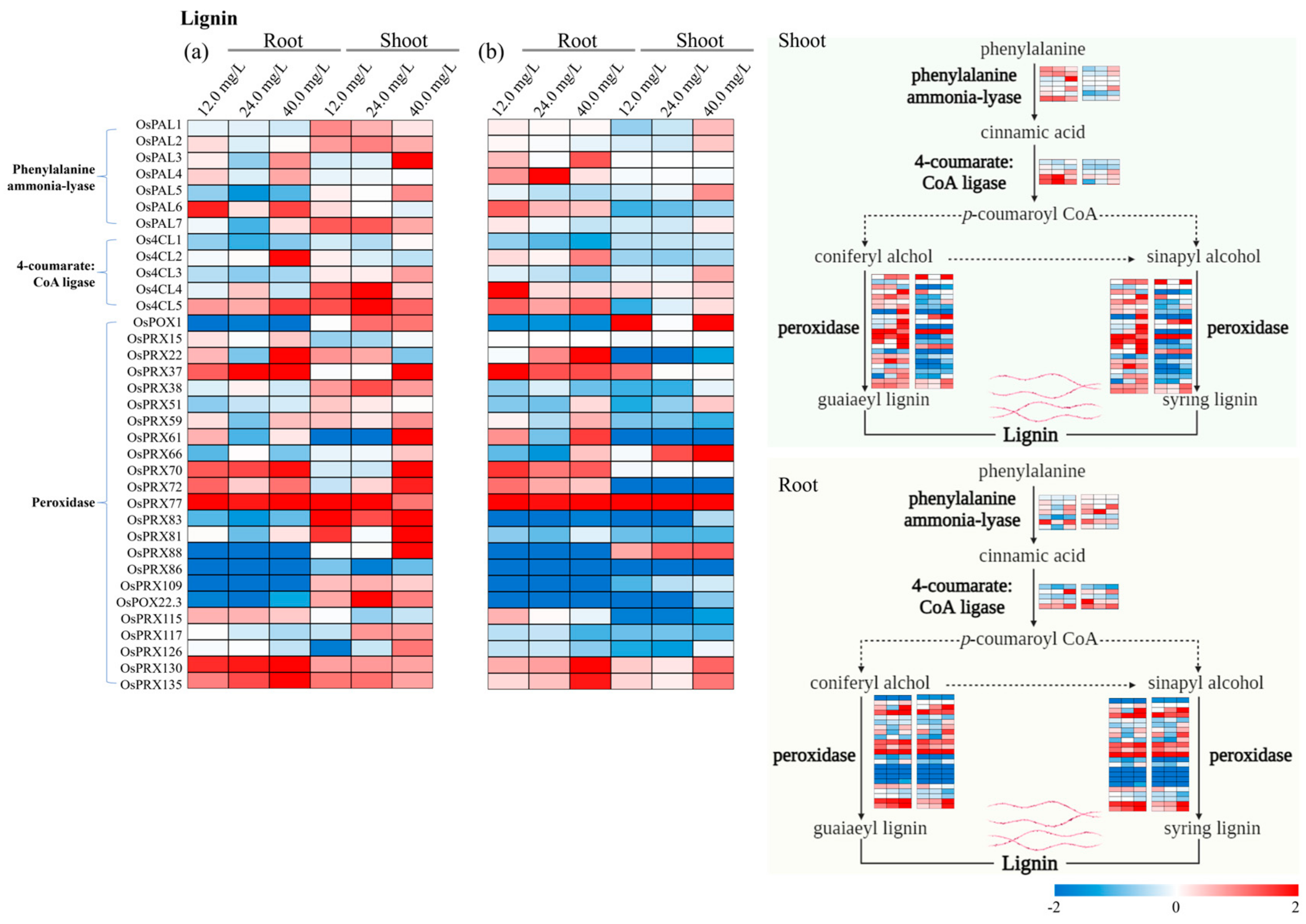
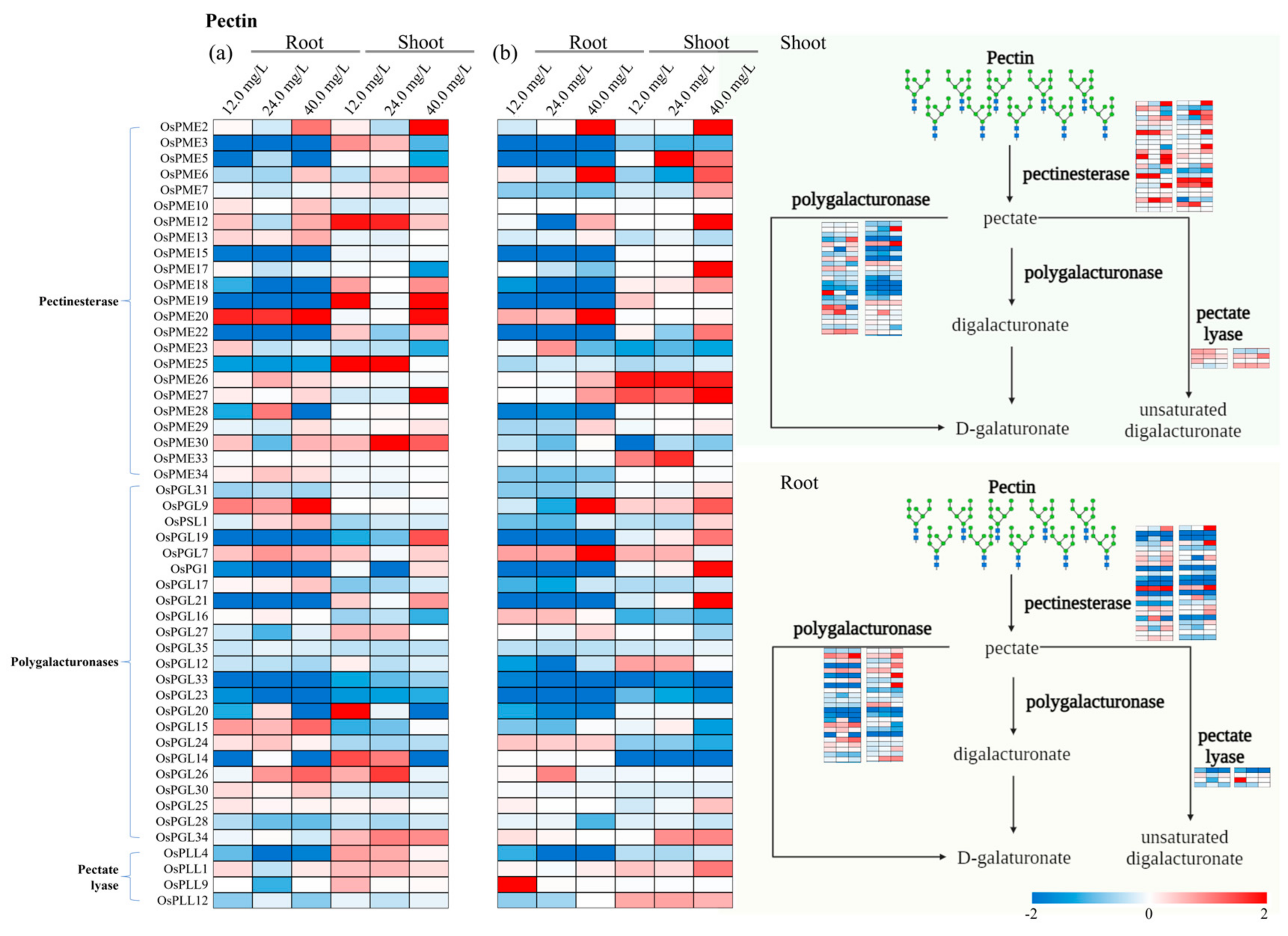
3.2. Differentially Expressed Genes Associated with Cell Wall Pathways
3.3. PCR Verification to Validate the Reliability of Rice Microarray
3.4. Specific Genes Activated in Cell Wall Biosynthesis
3.5. Estimation of Gene Expression Variation Factors (GEVFs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Danyal, Y.; Mahmood, K.; Ullah, S.; Rahim, A.; Raheem, G.; Khan, A.H.; Ullah, A. Phytoremediation of industrial effluents assisted by plant growth promoting bacteria. Environ. Sci. Pollut. Res. 2023, 30, 5296–5311. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.T.; Mfarrej, M.F.B.; Alam, P.; Rinklebe, J.; Ahmad, P. Accumulation of chromium in plants and its repercussion in animals and humans. Environ. Pollut. 2022, 301, 119044. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, D.; Wang, J.; Shahzad, B.; Kumar, V.; Bali, A.S.; Jasrotia, S.; Zheng, B.; Yuan, H.; Yan, D. Chromium bioaccumulation and its impacts on plants: An overview. Plants 2020, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, U.; Haider, F.U.; Ahmad, M.; Hussain, D.; Maqsood, M.F.; Ishfaq, M.; Eldin, S.M. Chromium toxicity, speciation, and remediation strategies in soil-plant interface: A critical review. Front. Plant Sci. 2023, 13, 5468. [Google Scholar] [CrossRef]
- Morkunas, I.; Woźniak, A.; Mai, V.C.; Rucińska-Sobkowiak, R.; Jeandet, P. The role of heavy metals in plant response to biotic stress. Molecules 2018, 23, 2320. [Google Scholar] [CrossRef]
- Parrotta, L.; Guerriero, G.; Sergeant, K.; Cai, G.; Hausman, J.F. Target or barrier? The cell wall of early-and later-diverging plants vs. cadmium toxicity: Differences in the response mechanisms. Front. Plant Sci. 2015, 6, 133. [Google Scholar] [CrossRef]
- Wu, Z.; McGrouther, K.; Chen, D.; Wu, W.; Wang, H. Subcellular distribution of metals within Brassica chinensis L. in response to elevated lead and chromium stress. J. Agric. Food Chem. 2013, 61, 4715–4722. [Google Scholar] [CrossRef]
- Yu, X.Z.; Feng, Y.X.; Liang, Y.P. Kinetics of phyto-accumulation of hexavalent and trivalent chromium in rice seedlings. Int. Biodeterior. Biodegrad. 2018, 128, 72–77. [Google Scholar] [CrossRef]
- Das, P.K.; Das, B.P.; Dash, P. Chromite mining pollution, environmental impact, toxicity and phytoremediation: A review. Environ. Chem. Lett. 2020, 19, 1369–1381. [Google Scholar] [CrossRef]
- Guo, X.; Ullah, A.; Siuta, D.; Kukfisz, B.; Iqbal, S. Role of WRKY transcription factors in regulation of abiotic stress responses in cotton. Life 2022, 12, 1410. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Wang, R.; Sun, X.; Liu, P.; Tang, J.; Zhang, C.; Liu, H. Heavy metal stress in plants: Ways to alleviate with exogenous substances. Sci. Total Environ. 2023, 897, 165397. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Asthir, B.J.B.P. Proline: A key player in plant abiotic stress tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Pan, X.; Ullah, A.; Feng, Y.X.; Tian, P.; Yu, X.Z. Proline-mediated activation of glyoxalase II improve methylglyoxal detoxification in Oryza sativa L. under chromium injury: Clarification via vector analysis of enzymatic activities and gene expression. Plant Physiol. Biochem. 2023, 201, 107867. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Hima Kumari, P.; Sunita, M.S.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [PubMed]
- Gothandam, K.M.; Nalini, E.; Karthikeyan, S.; Shin, J.S. OsPRP3, a flower specific proline-rich protein of rice, determines extracellular matrix structure of floral organs and its overexpression confers cold-tolerance. Plant Mol. Biol. 2010, 72, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Z.; Lu, C.J.; Tang, S.; Zhang, Q. Transcriptomic analysis of cytochrome P450 genes and pathways involved in chromium toxicity in Oryza sativa. Ecotoxicology 2020, 29, 503–513. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2018, 3, 1101–1108. [Google Scholar] [CrossRef]
- Kang, Y.; Zhang, Q.; Feng, Y.X.; Yang, L.; Yu, X. Exogenous proline activated an integrated response of NER and HR pathways to reduce DNA damage in rice seedlings under chromium stress. Environ. Sci. Pollut. Res. 2023, 30, 51792–51803. [Google Scholar] [CrossRef]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, N.K.; Dumat, C.; Rashid, M.I. Chromium speciation, bioavailability, uptake, toxicity and detoxification in soil-plant system: A review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef]
- Zhang, Q.; Feng, Y.X.; Lin, Y.J.; Yu, X.Z. Mathematical quantification of interactive complexity of transcription factors involved in proline-mediated regulative strategies in Oryza sativa under chromium stress. Plant Physiol. Biochem. 2022, 182, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Polko, J.K.; Kieber, J.J. The regulation of cellulose biosynthesis in plants. Plant Cell 2019, 31, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Singhania, R.R.; Patel, A.K.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Biores Technol. 2013, 127, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhao, X.; Shao, Z.; Zhu, L.; He, G. Multiple isoforms of UDP-glucose pyrophosphorylase in rice. Physiol. Plant 2007, 129, 725–736. [Google Scholar] [CrossRef]
- Cheng, X.; Wu, Y.; Guo, J.; Du, B.; Chen, R.; Zhu, L.; He, G. A rice lectin receptor-like kinase that is involved in innate immune responses also contributes to seed germination. Plant J. 2013, 76, 687–698. [Google Scholar] [CrossRef]
- Zhang, W.; Qin, W.; Li, H.; Wu, A.M. Biosynthesis and transport of nucleotide sugars for plant hemicellulose. Front. Plant Sci. 2021, 12, 723128. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, X.; Yao, W.; Gao, Y.; Zhao, K.; Guo, Q.; Zhou, B.; Jiang, T. Genome-wide identification and expression analysis of the xyloglucan endotransglucosylase/hydrolase gene family in poplar. BMC Genom. 2021, 22, 804. [Google Scholar] [CrossRef]
- Song, L.; Valliyodan, B.; Prince, S.; Wan, J.; Nguyen, H.T. Characterization of the XTH Gene Family: New Insight to the Roles in Soybean Flooding Tolerance. Int. J. Mol. Sci. 2018, 19, 2705. [Google Scholar] [CrossRef]
- Chhetri, H.B.; Macaya-Sanz, D.; Kainer, D.; Biswal, A.K.; Evans, L.M.; Chen, J.G.; DiFazio, S.P. Multitrait genome-wide association analysis of Populus trichocarpa identifies key polymorphisms controlling morphological and physiological traits. New Phytol. 2019, 223, 293–309. [Google Scholar] [CrossRef]
- Van Acker, R.; Vanholme, R.; Storme, V.; Mortimer, J.C.; Dupree, P.; Boerjan, W. Lignin biosynthesis perturbations affect secondary cell wall composition and saccharification yield in Arabidopsis thaliana. Biotechnol. Biofuels 2013, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Kunkowska, A.B.; Schippers, J.H. Role of reactive oxygen species during cell expansion in leaves. Plant Physiol. 2016, 172, 2098–2106. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Choi, H.S.; Cho, Y.C.; Kim, S.R. Cold-responsive regulation of a flower-preferential class III peroxidase gene, OsPOX1, in rice (Oryza sativa L.). J. Plant Biol. 2012, 55, 123–131. [Google Scholar] [CrossRef]
- Dash, L.; Swaminathan, S.; Šimura, J.; Gonzales, C.L.P.; Montes, C.; Solanki, N.; Kelley, D.R. Changes in cell wall composition due to a pectin biosynthesis enzyme GAUT10 impact root growth. Plant Physiol. 2023, 193, 2480–2497. [Google Scholar] [CrossRef]
- Wu, Q.; Ye, T.; Zhang, X.; Dong, X.; Xia, J.; Shen, R.; Zhu, X. Pectin methylesterases enhance root cell wall phosphorus remobilization in rice. Rice Sci. 2022, 29, 179–188. [Google Scholar] [CrossRef]
- Shi, D.; Ren, A.; Tang, X.; Qi, G.; Xu, Z.; Chai, G.; Hu, R.; Zhou, G.; Kong, Y. MYB52 negatively regulates pectin demethylesterification in seed coat mucilage. Plant Physiol. 2018, 176, 2737–2749. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Abbas, S.; Hassan, M.U.; Saeed, F.; Haider, S.; Varshney, R.K. Assessment of proline function in higher plants under extreme temperatures. Plant Biol. 2023, 25, 379–395. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, A.; Lin, Y.-J.; Zhang, H.; Yu, X.-Z. Identification of the Key Genes Involved in Proline-Mediated Modification of Cell Wall Components in Rice Seedlings under Trivalent Chromium Exposure. Toxics 2024, 12, 4. https://doi.org/10.3390/toxics12010004
Ullah A, Lin Y-J, Zhang H, Yu X-Z. Identification of the Key Genes Involved in Proline-Mediated Modification of Cell Wall Components in Rice Seedlings under Trivalent Chromium Exposure. Toxics. 2024; 12(1):4. https://doi.org/10.3390/toxics12010004
Chicago/Turabian StyleUllah, Abid, Yu-Juan Lin, Hua Zhang, and Xiao-Zhang Yu. 2024. "Identification of the Key Genes Involved in Proline-Mediated Modification of Cell Wall Components in Rice Seedlings under Trivalent Chromium Exposure" Toxics 12, no. 1: 4. https://doi.org/10.3390/toxics12010004
APA StyleUllah, A., Lin, Y.-J., Zhang, H., & Yu, X.-Z. (2024). Identification of the Key Genes Involved in Proline-Mediated Modification of Cell Wall Components in Rice Seedlings under Trivalent Chromium Exposure. Toxics, 12(1), 4. https://doi.org/10.3390/toxics12010004