

The Effects of Co-Exposure to Antifoulants and Microplastics on the Survival, Oxidative Status, and Cholinergic System of a Marine Mysid



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Maintenance
2.2. Exposure
2.3. Feeding Rate
2.4. Analysis of Oxidative Stress and Antioxidant Parameters
2.5. Measurement of Cholinergic Enzymes
2.6. Statistics
3. Results
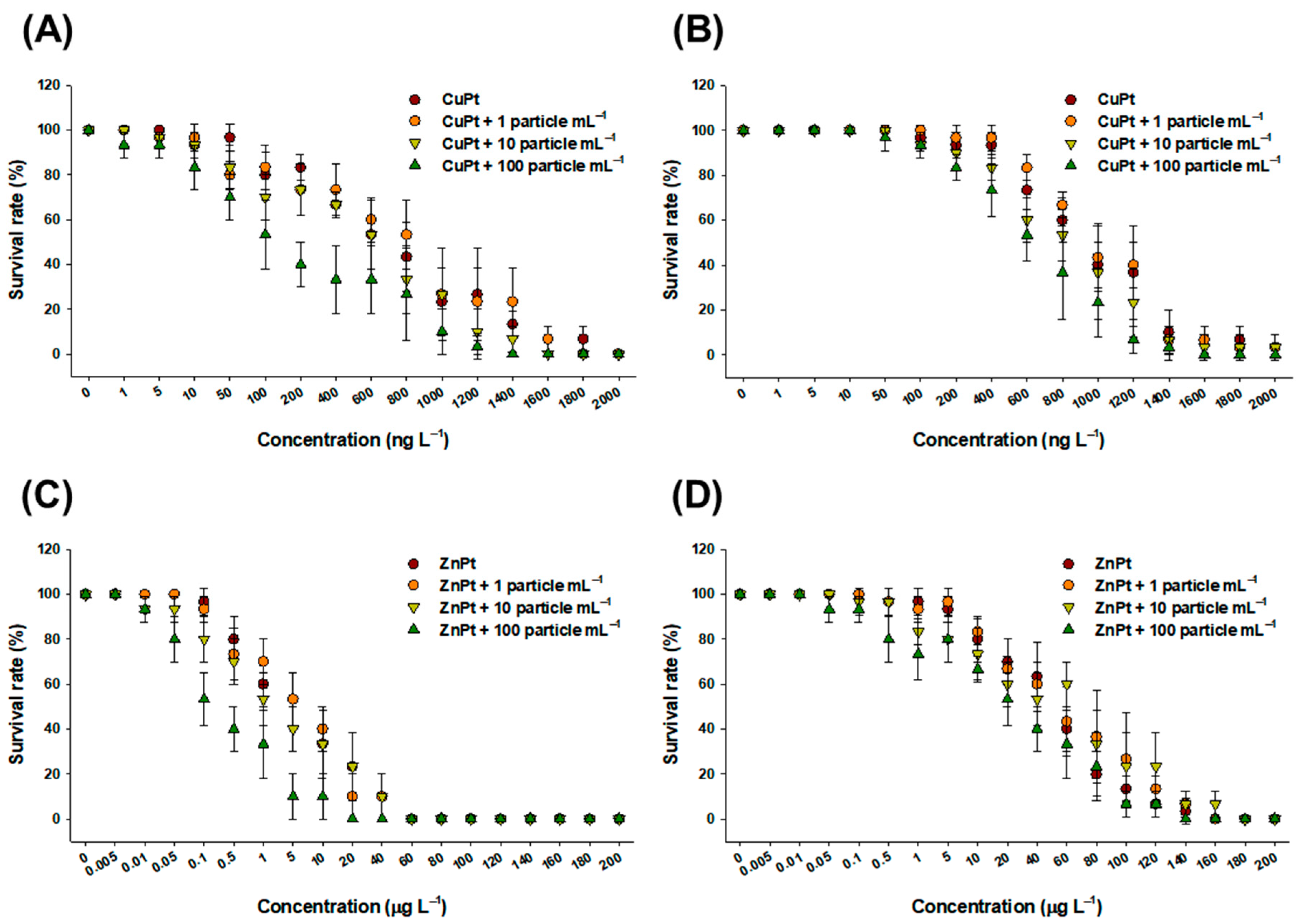
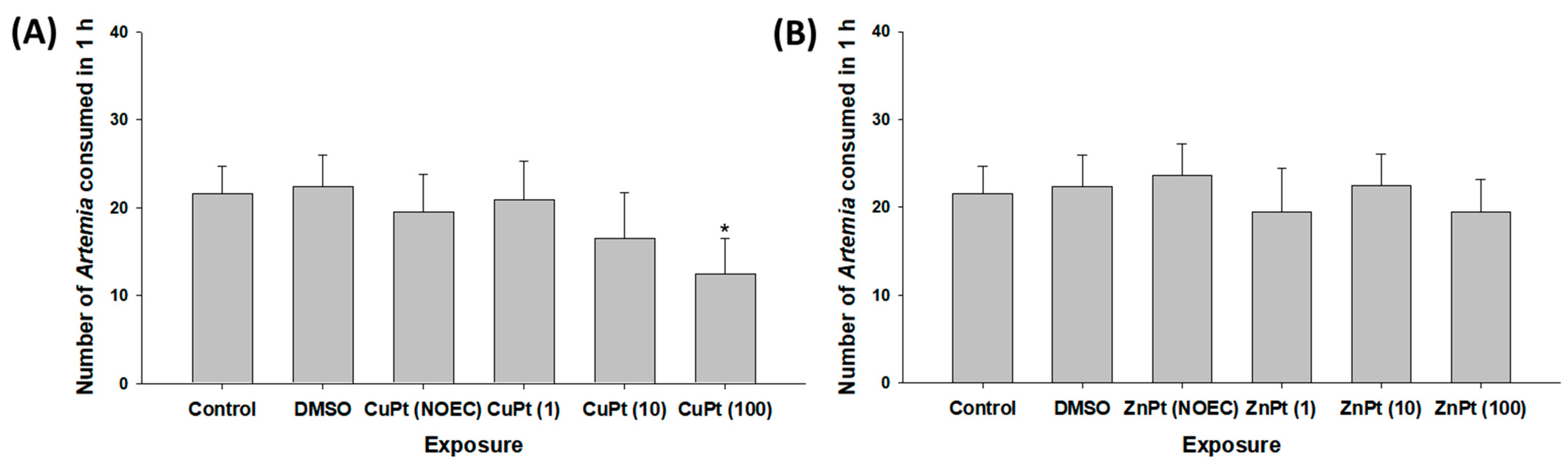
3.1. Effect on Survival and Feeding Rates
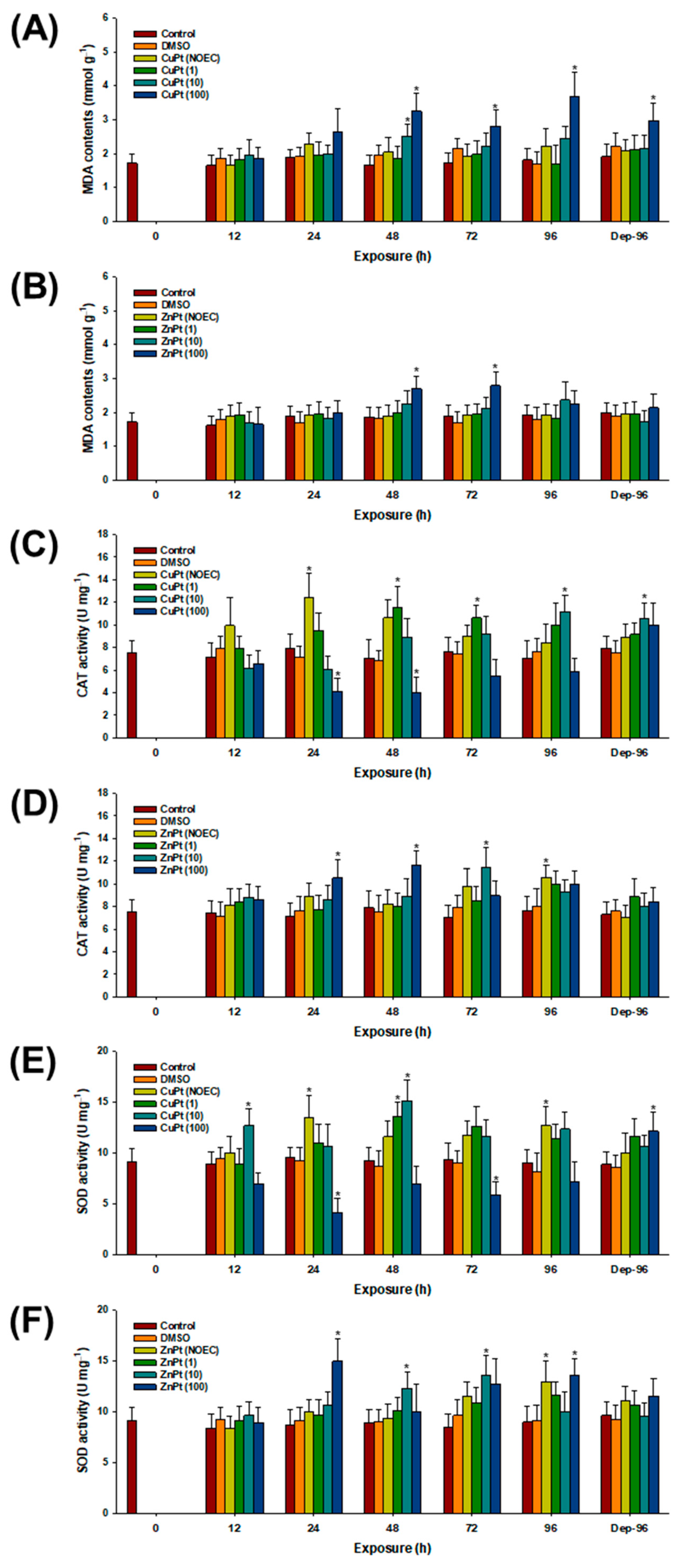
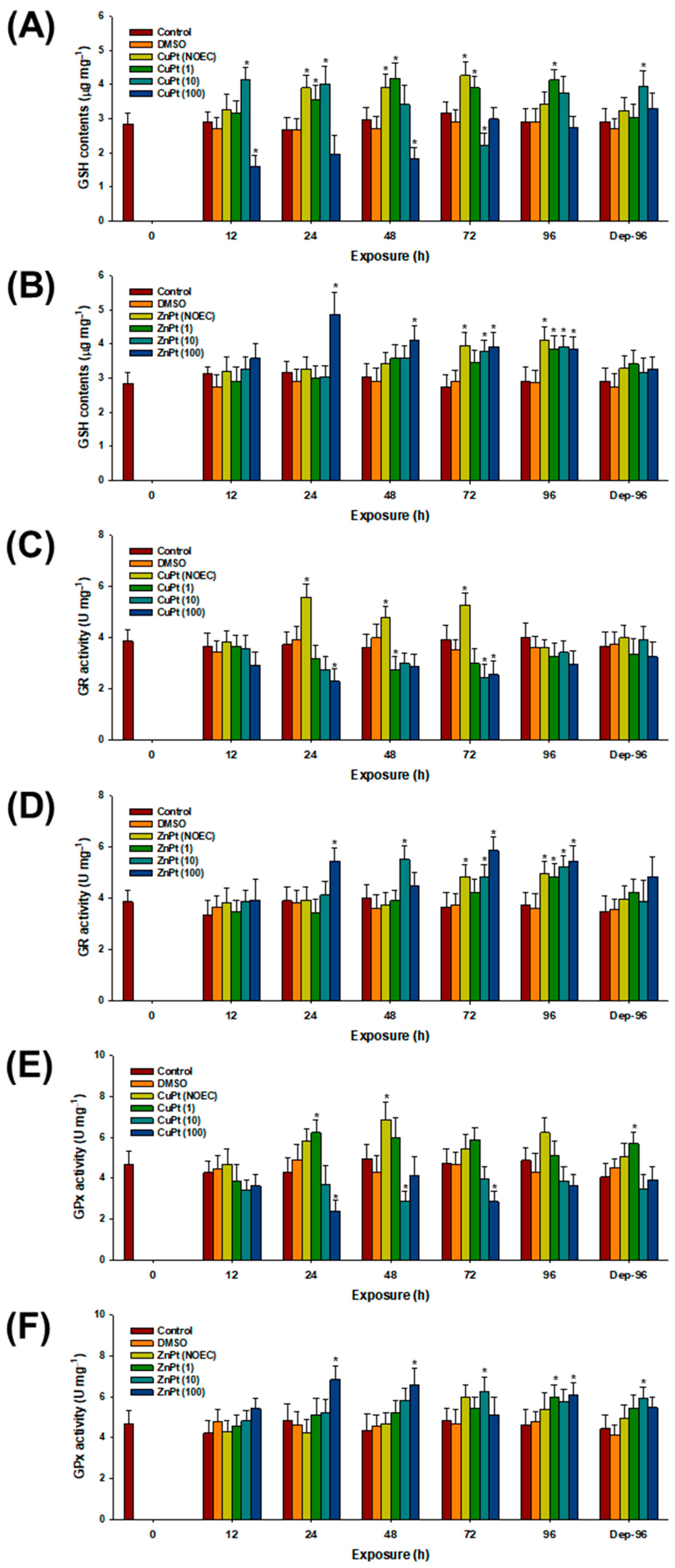
3.2. Oxidative Stress and Response of Antioxidant Parameters
3.3. Effect on AChE Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Maritime Organisation (IMO). International Convention on the Control of Harmful Antifouling Systems on Ships; International Maritime Organisation: London, UK, 2009. [Google Scholar]
- Konstantinou, I.; Albanis, T. Worldwide occurrence and effects of antifouling paint booster biocides in the aquatic environment: A review. Environ. Int. 2004, 30, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Yebra, D.M.; Kiil, S.; Dam-Johansen, K. Antifouling technology—Past, present and future steps towards efficient and environmentally friendly antifouling coatings. Prog. Org. Coat. 2004, 50, 75–104. [Google Scholar] [CrossRef]
- Martins, S.E.; Fillmann, G.; Lillicrap, A.; Thomas, K.V. Ecotoxicity of organic and organo-metallic antifouling co-biocides and implications for environmental hazard and risk assessments in aquatic ecosystems. Biofouling 2018, 34, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Turley, P.; Fenn, R.; Ritter, J. Pyrithiones as antifoulants: Environmental chemistry and preliminary risk assessment. Biofouling 2000, 15, 175–182. [Google Scholar] [CrossRef]
- Bao, V.W.; Leung, K.M.; Kwok, K.W.; Zhang, A.Q.; Lui, G.C. Synergistic toxic effects of zinc pyrithione and copper to three marine species: Implications on setting appropriate water quality criteria. Mar. Pollut. Bull. 2008, 57, 616–623. [Google Scholar] [CrossRef]
- Bellas, J. Toxicity assessment of the antifouling compound zinc pyrithione using early developmental stages of the ascidian Ciona intestinalis. Biofouling 2005, 21, 289–296. [Google Scholar] [CrossRef]
- Goka, K. Embryotoxicity of zinc pyrithione, an antidandruff chemical, in fish. Environ. Res. 1999, 81, 81–83. [Google Scholar] [CrossRef]
- Haque, M.N.; Nam, S.-E.; Eom, H.-J.; Kim, S.-K.; Rhee, J.-S. Exposure to sublethal concentrations of zinc pyrithione inhibits growth and survival of marine polychaete through induction of oxidative stress and DNA damage. Mar. Pollut. Bull. 2020, 156, 111276. [Google Scholar] [CrossRef]
- Jung, S.; Bae, J.; Kang, S.; Son, J.; Jeon, J.; Lee, H.; Jeon, J.; Sidharthan, M.; Ryu, S.; Shin, H. Acute toxicity of organic antifouling biocides to phytoplankton Nitzschia pungens and zooplankton Artemia larvae. Mar. Pollut. Bull. 2017, 124, 811–818. [Google Scholar] [CrossRef]
- Mochida, K.; Ito, K.; Harino, H.; Kakuno, A.; Fujii, K. Acute toxicity of pyrithione antifouling biocides and joint toxicity with copper to red sea bream (Pagrus major) and toy shrimp (Heptacarpus futilirostris). Environ. Toxicol. Chem. 2006, 25, 3058–3064. [Google Scholar] [CrossRef]
- Mochida, K.; Ito, K.; Harino, H.; Onduka, T.; Kakuno, A.; Fujii, K. Early life-stage toxicity test for copper pyrithione and induction of skeletal anomaly in a teleost, the mummichog (Fundulus heteroclitus). Environ. Toxicol. Chem. 2008, 27, 367–374. [Google Scholar] [CrossRef]
- Okamura, H.; Watanabe, T.; Aoyama, I.; Hasobe, M. Toxicity evaluation of new antifouling compounds using suspension-cultured fish cells. Chemosphere 2002, 46, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Y.; Sun, J.; Sha, H.; Yang, Y.; Ye, Q.; Yang, Q.; Huang, B.; Yu, Y.; Huang, H. Acute toxic responses of embryo-larval zebrafish to zinc pyrithione (ZPT) reveal embryological and developmental toxicity. Chemosphere 2018, 205, 62–70. [Google Scholar] [CrossRef]
- Lee, S.; Haque, M.N.; Lee, D.-H.; Rhee, J.-S. Comparison of the effects of sublethal concentrations of biofoulants, copper pyrithione and zinc pyrithione on a marine mysid-A multigenerational study. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2023, 271, 109694. [Google Scholar] [CrossRef] [PubMed]
- Barboza, L.G.A.; Vethaak, A.D.; Lavorante, B.R.; Lundebye, A.-K.; Guilhermino, L. Marine microplastic debris: An emerging issue for food security, food safety and human health. Mar. Pollut. Bull. 2018, 133, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Van Sebille, E.; Aliani, S.; Law, K.L.; Maximenko, N.; Alsina, J.M.; Bagaev, A.; Bergmann, M.; Chapron, B.; Chubarenko, I.; Cózar, A. The physical oceanography of the transport of floating marine debris. Environ. Res. Lett. 2020, 15, 023003. [Google Scholar] [CrossRef]
- Xu, S.; Ma, J.; Ji, R.; Pan, K.; Miao, A.-J. Microplastics in aquatic environments: Occurrence, accumulation, and biological effects. Sci. Total Environ. 2020, 703, 134699. [Google Scholar] [CrossRef]
- Bhagat, J.; Nishimura, N.; Shimada, Y. Toxicological interactions of microplastics/nanoplastics and environmental contaminants: Current knowledge and future perspectives. J. Hazard. Mater. 2021, 405, 123913. [Google Scholar] [CrossRef]
- Rubin, A.E.; Zucker, I. Interactions of microplastics and organic compounds in aquatic environments: A case study of augmented joint toxicity. Chemosphere 2022, 289, 133212. [Google Scholar] [CrossRef]
- Thomas, J.; Patil, R.S.; Patil, M.; John, J. Addressing the Sustainability Conundrums and Challenges within the Polymer Value Chain. Sustainability 2023, 15, 15758. [Google Scholar] [CrossRef]
- Godoy, V.; Blázquez, G.; Calero, M.; Quesada, L.; Martín-Lara, M. The potential of microplastics as carriers of metals. Environ. Pollut. 2019, 255, 113363. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yang, C.; Zhu, Z.; Bai, X.; Ma, J. Adsorption behavior of organic pollutants and metals on micro/nanoplastics in the aquatic environment. Sci. Total Environ. 2019, 694, 133643. [Google Scholar] [CrossRef]
- Lee, D.-H.; Nam, S.-E.; Eom, H.-J.; Rhee, J.-S. Analysis of effects of environmental fluctuations on the marine mysid Neomysis awatschensis and its development as an experimental model animal. J. Sea Res. 2020, 156, 101834. [Google Scholar] [CrossRef]
- Verslycke, T.A.; Fockedey, N.; McKenney, C.L., Jr.; Roast, S.D.; Jones, M.B.; Mees, J.; Janssen, C.R. Mysid crustaceans as potential test organisms for the evaluation of environmental endocrine disruption: A review. Environ. Toxicol. Chem. 2004, 23, 1219–1234. [Google Scholar] [CrossRef]
- Hook, S.E.; Gallagher, E.P.; Batley, G.E. The role of biomarkers in the assessment of aquatic ecosystem health. Integr. Environ. Assess. Manag. 2014, 10, 327–341. [Google Scholar] [CrossRef]
- Nam, S.-E.; Bae, D.-Y.; Ki, J.-S.; Ahn, C.-Y.; Rhee, J.-S. The importance of multi-omics approaches for the health assessment of freshwater ecosystems. Mol. Cell. Toxicol. 2023, 19, 3–11. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef]
- Livingstone, D. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Massoulié, J.; Sussman, J.; Bon, S.; Silman, I. Structure and functions of acetylcholinesterase and butyrylcholinesterase. Prog. Brain Res. 1993, 98, 139–146. [Google Scholar]
- Lee, D.-H.; Lee, S.; Rhee, J.-S. Consistent exposure to microplastics induces age-specific physiological and biochemical changes in a marine mysid. Mar. Pollut. Bull. 2021, 162, 111850. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Holmes, L.A.; Turner, A.; Thompson, R.C. Interactions between trace metals and plastic production pellets under estuarine conditions. Mar. Chem. 2014, 167, 25–32. [Google Scholar] [CrossRef]
- Rochman, C.M.; Hentschel, B.T.; Teh, S.J. Long-term sorption of metals is similar among plastic types: Implications for plastic debris in aquatic environments. PLoS ONE 2014, 9, e85433. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, L.; Zheng, H.; Wang, M.; Fu, Y.; Luo, X.; Li, F.; Wang, Z. Polystyrene microplastics impaired the feeding and swimming behavior of mysid shrimp Neomysis japonica. Mar. Pollut. Bull. 2020, 150, 110660. [Google Scholar] [CrossRef]
- Jitrapat, H.; Sivaipram, I.; Piumsomboon, A.; Suttiruengwong, S.; Xu, J.; Vo, T.L.T.; Li, D. Ingestion and adherence of microplastics by estuarine mysid shrimp. Mar. Environ. Res. 2024, 197, 106455. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Haque, M.N.; Rhee, J.-S. Acute and mutigenerational effects of environmental concentration of the antifouling agent dichlofluanid on the mysid model, Neomysis awatschensis. Environ. Pollut. 2022, 311, 119996. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Nam, S.-E.; Jung, J.-H.; Kim, M.; Rhee, J.-S. Hull-cleaning wastewater poses serious acute and chronic toxicity to a marine mysid—A multigenerational study. J. Hazard. Mater. 2024, 469, 133959. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Rikans, L.E.; Hornbrook, K.R. Lipid peroxidation, antioxidant protection and aging. Biochim. Biophys. Acta Mol. Basis Dis. 1997, 1362, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Chan, H.; Yu, L. Glutathione peroxidase and glutathione reductase activities are partially responsible for determining the susceptibility of cells to oxidative stress. Toxicology 2006, 226, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Felten, V.; Toumi, H.; Masfaraud, J.-F.; Billoir, E.; Camara, B.I.; Férard, J.-F. Microplastics enhance Daphnia magna sensitivity to the pyrethroid insecticide deltamethrin: Effects on life history traits. Sci. Total Environ. 2020, 714, 136567. [Google Scholar] [CrossRef]
- Nunes, B. The use of cholinesterases in ecotoxicology. Rev. Environ. Contam. Toxicol. 2011, 212, 29–59. [Google Scholar] [PubMed]
- Trang, A.; Khandhar, P. Physiology, Acetylcholinesterase. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Eom, H.-J.; Haque, M.N.; Lee, S.; Rhee, J.-S. Exposure to metals premixed with microplastics increases toxicity through bioconcentration and impairs antioxidant defense and cholinergic response in a marine mysid. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 249, 109142. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Cidlowski, J.A. Glutathione efflux and cell death. Antioxid. Redox. Signal 2012, 17, 1694–1713. [Google Scholar] [CrossRef]
- Gomiero, A.; Strafella, P.; Pellini, G.; Salvalaggio, V.; Fabi, G. Comparative effects of ingested PVC micro particles with and without adsorbed benzo (a) pyrene vs. spiked sediments on the cellular and sub cellular processes of the benthic organism Hediste diversicolor. Front. Mar. Sci. 2018, 5, 99. [Google Scholar] [CrossRef]
- Pacheco, A.; Martins, A.; Guilhermino, L. Toxicological interactions induced by chronic exposure to gold nanoparticles and microplastics mixtures in Daphnia magna. Sci. Total Environ. 2018, 628, 474–483. [Google Scholar] [CrossRef]
- Qiao, R.; Lu, K.; Deng, Y.; Ren, H.; Zhang, Y. Combined effects of polystyrene microplastics and natural organic matter on the accumulation and toxicity of copper in zebrafish. Sci. Total Environ. 2019, 682, 128–137. [Google Scholar] [CrossRef]
- Yoo, J.-W.; Choi, T.-J.; Park, J.-S.; Kim, J.; Han, S.; Kim, C.-B.; Lee, Y.-M. Pathway-dependent toxic interaction between polystyrene microbeads and methylmercury on the brackish water flea Diaphanosoma celebensis: Based on mercury bioaccumulation, cytotoxicity, and transcriptomic analysis. J. Hazard. Mater. 2023, 459, 132055. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Haque, M.N.; Lee, D.-H.; Rhee, J.-S. The Effects of Co-Exposure to Antifoulants and Microplastics on the Survival, Oxidative Status, and Cholinergic System of a Marine Mysid. Toxics 2024, 12, 651. https://doi.org/10.3390/toxics12090651
Lee S, Haque MN, Lee D-H, Rhee J-S. The Effects of Co-Exposure to Antifoulants and Microplastics on the Survival, Oxidative Status, and Cholinergic System of a Marine Mysid. Toxics. 2024; 12(9):651. https://doi.org/10.3390/toxics12090651
Chicago/Turabian StyleLee, Somyeong, Md. Niamul Haque, Do-Hee Lee, and Jae-Sung Rhee. 2024. "The Effects of Co-Exposure to Antifoulants and Microplastics on the Survival, Oxidative Status, and Cholinergic System of a Marine Mysid" Toxics 12, no. 9: 651. https://doi.org/10.3390/toxics12090651
APA StyleLee, S., Haque, M. N., Lee, D.-H., & Rhee, J.-S. (2024). The Effects of Co-Exposure to Antifoulants and Microplastics on the Survival, Oxidative Status, and Cholinergic System of a Marine Mysid. Toxics, 12(9), 651. https://doi.org/10.3390/toxics12090651