
The Mechanism of Arsenic Release in Contaminated Paddy Soil with Added Biochar: The Role of Dissolved Organic Matter, Fe, and Bacteria


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Biochar Preparation and Characterization
2.2. Description of Soil Samples
2.3. Experiment Design
2.4. The Analysis Methods
2.4.1. Analyses of Supernatant pH, Eh, and DOM Content
2.4.2. The Analysis of Fe and As Levels
2.4.3. The 16s rRNA Analysis
2.5. Data Analysis
3. Results
3.1. pH, Eh, and DOM Dynamics in the Supernatant
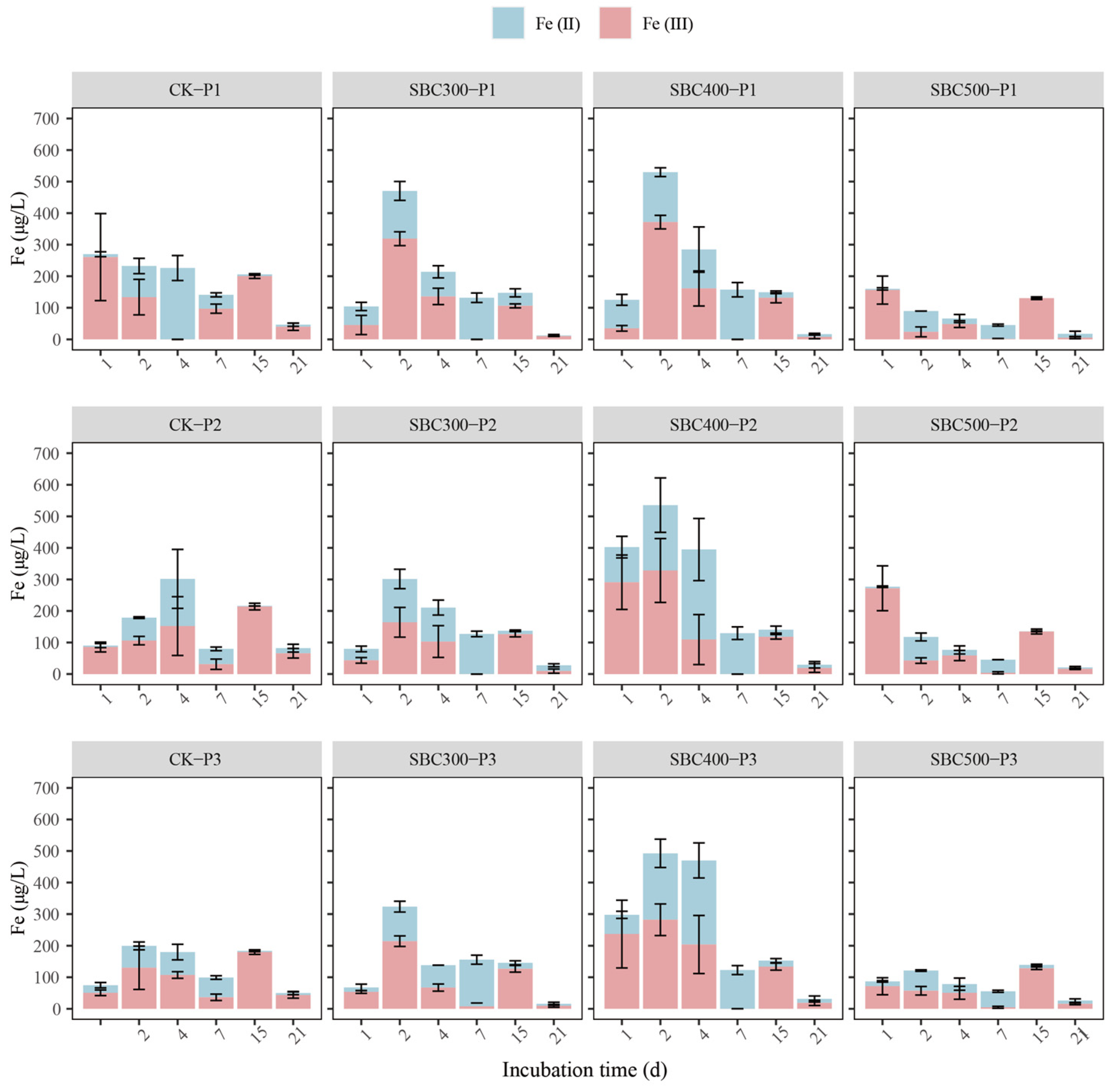
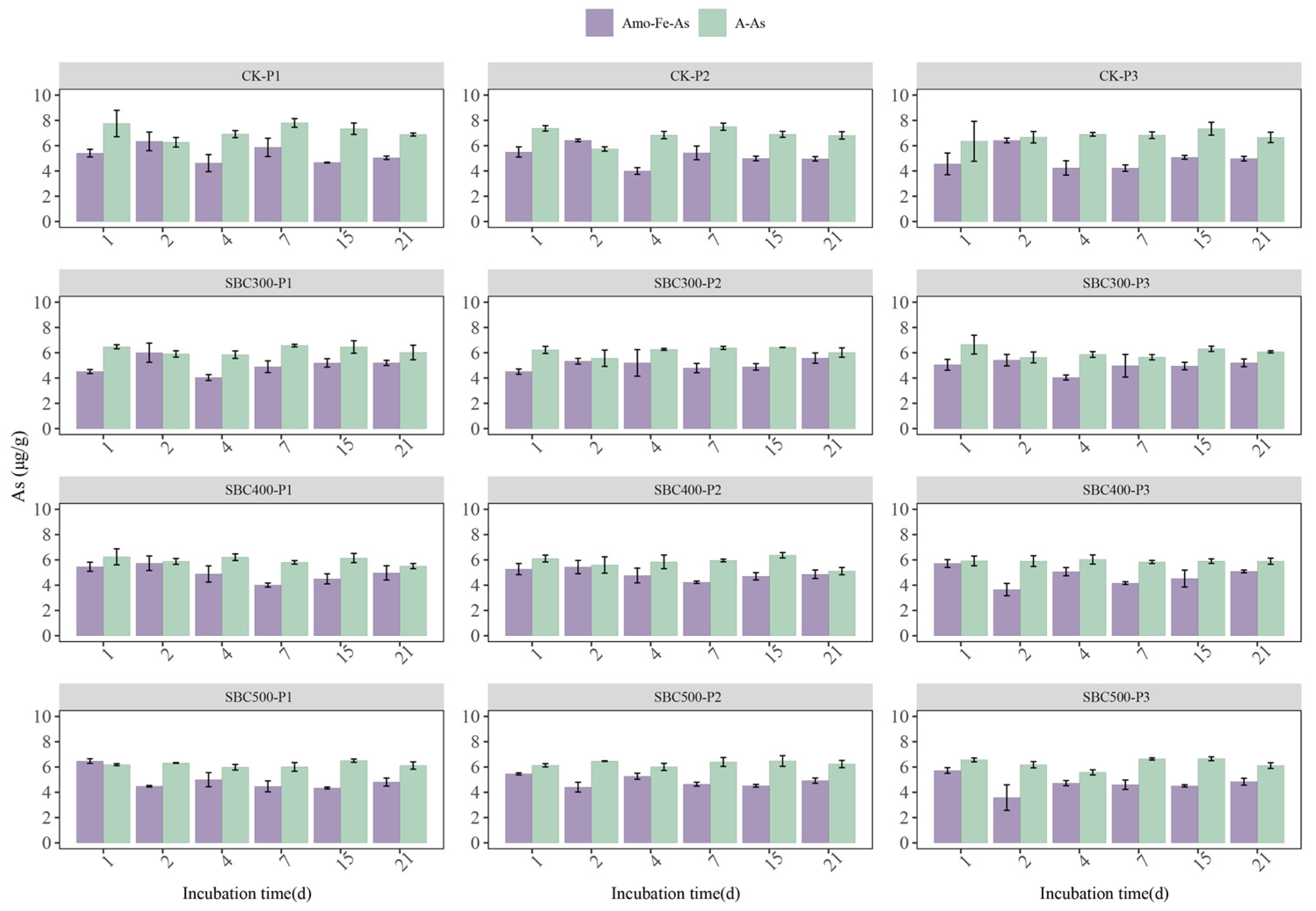
3.2. Fe and As Dynamics
3.2.1. Fe and As Dynamics in Supernatant
3.2.2. Fe and As Dynamics in Soil
3.3. Dynamics of Bacterial Diversity and Composition
3.3.1. Bacterial Composition Dynamics at the Phylum Level
3.3.2. Bacterial Composition Dynamics at the Genera Level
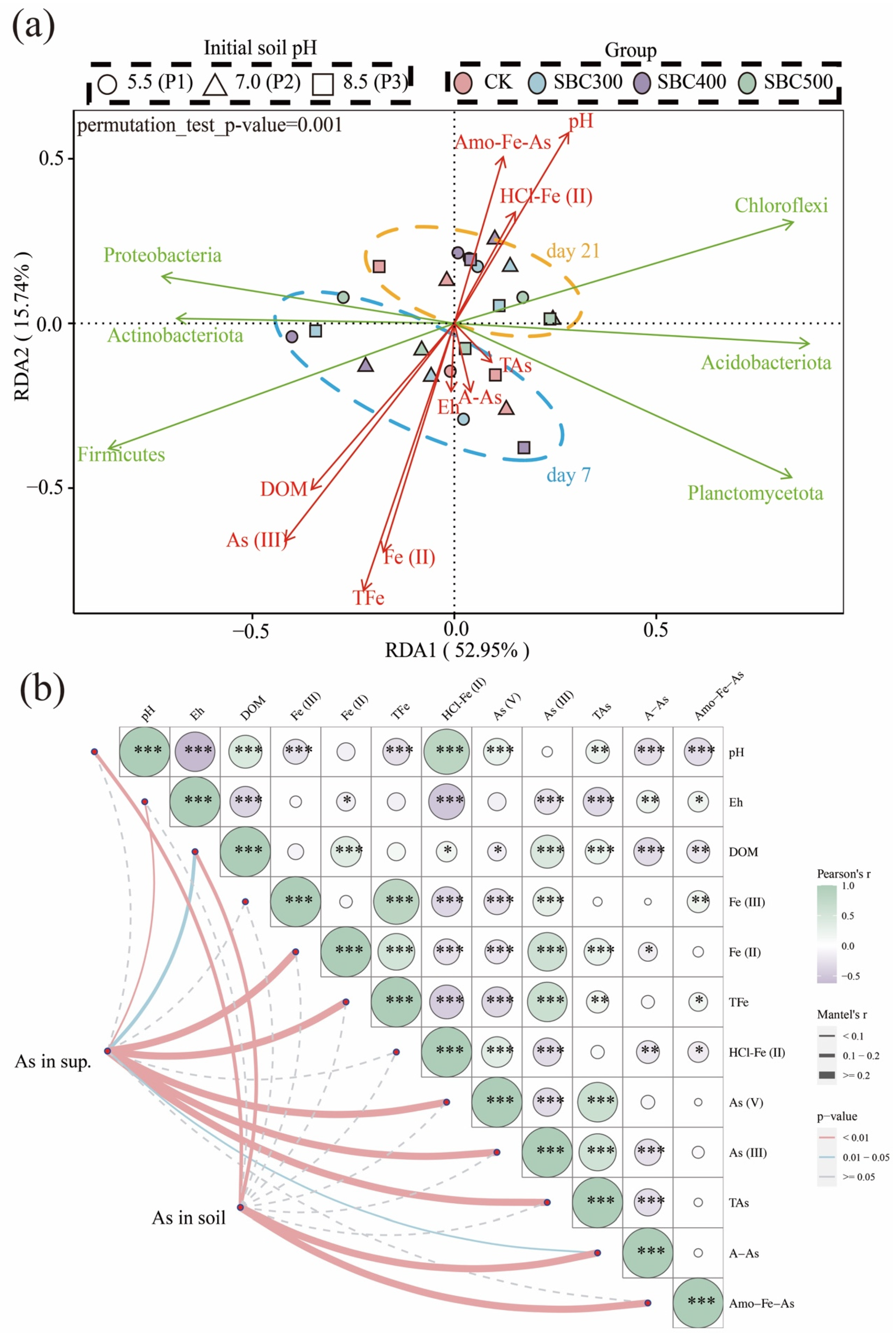
3.3.3. The Results of Cluster Analysis and RDA
3.4. The Results of the Mantel Test and Pearson Correlation Analysis
4. Discussion
4.1. Effect of Pyrolysis Temperature on Soil Component Dynamics
4.2. Possible Mechanisms for Biochar’s Effects on As Dynamics
4.3. Effect of Pyrolysis Temperature on As Transformation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Panthri, M.; Gupta, M. An Insight into the Act of Iron to Impede Arsenic Toxicity in Paddy Agro-System. J. Environ. Manag. 2022, 316, 115289. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, Q.; Zhang, L.; Huang, Z.; Zhao, Z.; Zhao, H.; Du, J.; Zhou, J. Toxic Metals in a Paddy Field System: A Review. Toxics 2022, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Chatterjee, S.; Moogouei, R.; Gupta, D. Arsenic Accumulation in Rice and Probable Mitigation Approaches: A Review. Agronomy 2017, 7, 67. [Google Scholar] [CrossRef]
- Stroud, J.L.; Norton, G.J.; Islam, M.R.; Dasgupta, T.; White, R.P.; Price, A.H.; Meharg, A.A.; McGrath, S.P.; Zhao, F.J. The Dynamics of Arsenic in Four Paddy Fields in the Bengal Delta. Environ. Pollut. 2011, 159, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tang, Z.; Chen, X.-P.; Wang, X.; Zhou, W.-X.; Tang, Z.; Zhang, J.; Zhao, F.-J. Water Management Impacts the Soil Microbial Communities and Total Arsenic and Methylated Arsenicals in Rice Grains. Environ. Pollut. 2019, 247, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.B.; Duckworth, O.W.; Stýblo, M.; Cable, P.H.; Alleoni, L.R.F. Pyrolysis Temperature and Biochar Redox Activity on Arsenic Availability and Speciation in a Sediment. J. Hazard. Mater. 2023, 460, 132308. [Google Scholar] [CrossRef]
- Sun, S.-K.; Chen, Y.; Che, J.; Konishi, N.; Tang, Z.; Miller, A.J.; Ma, J.F.; Zhao, F.-J. Decreasing Arsenic Accumulation in Rice by Overexpressing OsNIP1;1 and OsNIP3;3 through Disrupting Arsenite Radial Transport in Roots. New Phytol. 2018, 219, 641–653. [Google Scholar] [CrossRef]
- Kim, H.-B.; Kim, J.-G.; Kim, T.; Alessi, D.S.; Baek, K. Interaction of Biochar Stability and Abiotic Aging: Influences of Pyrolysis Reaction Medium and Temperature. Chem. Eng. J. 2021, 411, 128441. [Google Scholar] [CrossRef]
- Yuan, P.; Wang, J.; Pan, Y.; Shen, B.; Wu, C. Review of Biochar for the Management of Contaminated Soil: Preparation, Application and Prospect. Sci. Total Environ. 2019, 659, 473–490. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, W.; Duan, A.; Zeng, G.; Huang, D.; Lai, C.; Tan, X.; Cheng, M.; Wang, R.; Zhou, C.; et al. Adsorption Behavior of Engineered Carbons and Carbon Nanomaterials for Metal Endocrine Disruptors: Experiments and Theoretical Calculation. Chemosphere 2019, 222, 184–194. [Google Scholar] [CrossRef]
- Hassan, M.; Liu, Y.; Naidu, R.; Parikh, S.J.; Du, J.; Qi, F.; Willett, I.R. Influences of Feedstock Sources and Pyrolysis Temperature on the Properties of Biochar and Functionality as Adsorbents: A Meta-Analysis. Sci. Total Environ. 2020, 744, 140714. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhu, Y.; Cheng, L.; Andserson, B.; Zhao, X.; Wang, D.; Ding, A. Review on Utilization of Biochar for Metal-Contaminated Soil and Sediment Remediation. J. Environ. Sci. 2018, 63, 156–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kwak, J.-H.; Islam, M.S.; Naeth, M.A.; Gamal El-Din, M.; Chang, S.X. Biochar Surface Complexation and Ni(II), Cu(II), and Cd(II) Adsorption in Aqueous Solutions Depend on Feedstock Type. Sci. Total Environ. 2020, 712, 136538. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Ngo, H.H.; Guo, W.; Wen, H.; Zhang, D.; Li, C.; Qi, L. Characterization and Sulfonamide Antibiotics Adsorption Capacity of Spent Coffee Grounds Based Biochar and Hydrochar. Sci. Total Environ. 2020, 716, 137015. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-J.; Jiang, H.; Yu, H.-Q. Development of Biochar-Based Functional Materials: Toward a Sustainable Platform Carbon Material. Chem. Rev. 2015, 115, 12251–12285. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Ma, F.; Tankpa, V.; Bai, S.; Guo, X.; Wang, X. Mechanisms and Reutilization of Modified Biochar Used for Removal of Heavy Metals from Wastewater: A Review. Sci. Total Environ. 2019, 668, 1298–1309. [Google Scholar] [CrossRef]
- He, D.; Luo, Y.; Zhu, B. Feedstock and Pyrolysis Temperature Influence Biochar Properties and Its Interactions with Soil Substances: Insights from a DFT Calculation. Sci. Total Environ. 2024, 922, 171259. [Google Scholar] [CrossRef]
- Sugawara, K.; Ichio, K.; Ichikawa, Y.; Ogawa, H.; Suzuki, S. Effects of Pyrolysis Temperature and Chemical Modification on the Adsorption of Cd and As(V) by Biochar Derived from Pteris Vittata. Int. J. Environ. Res. Public Health 2022, 19, 5226. [Google Scholar] [CrossRef] [PubMed]
- Banik, C.; Lawrinenko, M.; Bakshi, S.; Laird, D.A. Impact of Pyrolysis Temperature and Feedstock on Surface Charge and Functional Group Chemistry of Biochars. J. Environ. Qual. 2018, 47, 452–461. [Google Scholar] [CrossRef]
- Amen, R.; Bashir, H.; Bibi, I.; Shaheen, S.M.; Niazi, N.K.; Shahid, M.; Hussain, M.M.; Antoniadis, V.; Shakoor, M.B.; Al-Solaimani, S.G.; et al. A Critical Review on Arsenic Removal from Water Using Biochar-Based Sorbents: The Significance of Modification and Redox Reactions. Chem. Eng. J. 2020, 396, 125195. [Google Scholar] [CrossRef]
- Niazi, N.K.; Bibi, I.; Shahid, M.; Ok, Y.S.; Burton, E.D.; Wang, H.; Shaheen, S.M.; Rinklebe, J.; Lüttge, A. Arsenic Removal by Perilla Leaf Biochar in Aqueous Solutions and Groundwater: An Integrated Spectroscopic and Microscopic Examination. Environ. Pollut. 2018, 232, 31–41. [Google Scholar] [CrossRef]
- Fidel, R.B.; Laird, D.A.; Thompson, M.L.; Lawrinenko, M. Characterization and Quantification of Biochar Alkalinity. Chemosphere 2017, 167, 367–373. [Google Scholar] [CrossRef]
- Singh, H.; Northup, B.K.; Rice, C.W.; Prasad, P.V.V. Biochar Applications Influence Soil Physical and Chemical Properties, Microbial Diversity, and Crop Productivity: A Meta-Analysis. Biochar 2022, 4, 8. [Google Scholar] [CrossRef]
- Qiu, J.; Fernandes de Souza, M.; Edayilam, N.; Yang, Y.; Ok, Y.S.; Ronsse, F.; Morabito, D.; Meers, E. Metal Behavior and Soil Quality Changes Induced by the Application of Tailor-Made Combined Biochar: An Investigation at Pore Water Scale. Sci. Total Environ. 2023, 898, 165552. [Google Scholar] [CrossRef]
- Bornø, M.L.; Müller-Stöver, D.S.; Liu, F. Biochar Properties and Soil Type Drive the Uptake of Macro- and Micronutrients in Maize (Zea Mays L.). J. Plant Nutr. Soil Sci. 2019, 182, 149–158. [Google Scholar] [CrossRef]
- Rasul, M.; Cho, J.; Shin, H.-S.; Hur, J. Biochar-Induced Priming Effects in Soil via Modifying the Status of Soil Organic Matter and Microflora: A Review. Sci. Total Environ. 2022, 805, 150304. [Google Scholar] [CrossRef]
- Liu, P.; Ptacek, C.J.; Blowes, D.W.; Berti, W.R.; Landis, R.C. Aqueous Leaching of Organic Acids and Dissolved Organic Carbon from Various Biochars Prepared at Different Temperatures. J. Environ. Qual. 2015, 44, 684–695. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Z.; Xiao, X.; Zhang, N.; Wang, X.; Yang, Z.; Xu, K.; Liang, Y. Structural Changes of Soil Organic Matter and the Linkage to Rhizosphere Bacterial Communities with Biochar Amendment in Manure Fertilized Soils. Sci. Total Environ. 2019, 692, 333–343. [Google Scholar] [CrossRef]
- Li, K.; Chen, J.; Sun, W.; Zhou, H.; Zhang, Y.; Yuan, H.; Hu, A.; Wang, D.; Zhang, W. Coupling Effect of DOM and Microbe on Arsenic Speciation and Bioavailability in Tailings Soil after the Addition of Different Biologically Stabilized Sludges. J. Hazard. Mater. 2023, 458, 132048. [Google Scholar] [CrossRef]
- Li, G.; Khan, S.; Ibrahim, M.; Sun, T.-R.; Tang, J.-F.; Cotner, J.B.; Xu, Y.-Y. Biochars Induced Modification of Dissolved Organic Matter (DOM) in Soil and Its Impact on Mobility and Bioaccumulation of Arsenic and Cadmium. J. Hazard. Mater. 2018, 348, 100–108. [Google Scholar] [CrossRef]
- Tang, L.; Xiong, L.; Zhang, H.; Joseph, A.; Wang, Y.; Li, J.; Yuan, X.; Rene, E.R.; Zhu, N. Reduced Arsenic Availability in Paddy Soil through Fe-Organic Ligand Complexation Mediated by Bamboo Biochar. Chemosphere 2023, 140790. [Google Scholar] [CrossRef]
- Xiang, Y.; Liu, Y.; Niazi, N.K.; Bolan, N.; Zhao, L.; Zhang, S.; Xue, J.; Yao, B.; Li, Y. Biochar Addition Increased Soil Bacterial Diversity and Richness: Large-Scale Evidence of Field Experiments. Sci. Total Environ. 2023, 893, 164961. [Google Scholar] [CrossRef]
- Majumder, S.; Banik, P. Geographical Variation of Arsenic Distribution in Paddy Soil, Rice and Rice-Based Products: A Meta-Analytic Approach and Implications to Human Health. J. Environ. Manag. 2019, 233, 184–199. [Google Scholar] [CrossRef]
- Smith, E.; Naidu, R.; Alston, A.M. Chemistry of Arsenic in Soils: I. Sorption of Arsenate and Arsenite by Four Australian Soils. J. Environ. Qual. 1999, 28, 1719–1726. [Google Scholar] [CrossRef]
- Feng, F.; Jiang, Y.; Jia, Y.; Lian, X.; Shang, C.; Zhao, M. Exogenous-Organic-Matter-Driven Mobilization of Groundwater Arsenic. Environ. Sci. Ecotechnology 2023, 15, 100243. [Google Scholar] [CrossRef]
- Mladenov, N.; Zheng, Y.; Miller, M.P.; Nemergut, D.R.; Legg, T.; Simone, B.; Hageman, C.; Rahman, M.M.; Ahmed, K.M.; McKnight, D.M. Dissolved Organic Matter Sources and Consequences for Iron and Arsenic Mobilization in Bangladesh Aquifers. Environ. Sci. Technol. 2010, 44, 123–128. [Google Scholar] [CrossRef]
- Kulkarni, H.V.; Mladenov, N.; McKnight, D.M.; Zheng, Y.; Kirk, M.F.; Nemergut, D.R. Dissolved Fulvic Acids from a High Arsenic Aquifer Shuttle Electrons to Enhance Microbial Iron Reduction. Sci. Total Environ. 2018, 615, 1390–1395. [Google Scholar] [CrossRef]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; de Hollander, M.; Soto, R.L.; Bouffaud, M.-L.; Buée, M.; Dimmers, W.; et al. Soil Networks Become More Connected and Take up More Carbon as Nature Restoration Progresses. Nat Commun 2017, 8, 14349. [Google Scholar] [CrossRef]
- Qiao, J.; Yu, H.; Wang, X.; Li, F.; Wang, Q.; Yuan, Y.; Liu, C. The Applicability of Biochar and Zero-Valent Iron for the Mitigation of Arsenic and Cadmium Contamination in an Alkaline Paddy Soil. Biochar 2019, 1, 203–212. [Google Scholar] [CrossRef]
- Qiao, J.; Li, X.; Hu, M.; Li, F.; Young, L.Y.; Sun, W.; Huang, W.; Cui, J. Transcriptional Activity of Arsenic-Reducing Bacteria and Genes Regulated by Lactate and Biochar during Arsenic Transformation in Flooded Paddy Soil. Environ. Sci. Technol. 2018, 52, 61–70. [Google Scholar] [CrossRef]
- Qiao, J.; Li, X.; Li, F. Roles of Different Active Metal-Reducing Bacteria in Arsenic Release from Arsenic-Contaminated Paddy Soil Amended with Biochar. J. Hazard. Mater. 2018, 344, 958–967. [Google Scholar] [CrossRef]
- Wei, S.; Zhu, M.; Fan, X.; Song, J.; Peng, P.; Li, K.; Jia, W.; Song, H. Influence of Pyrolysis Temperature and Feedstock on Carbon Fractions of Biochar Produced from Pyrolysis of Rice Straw, Pine Wood, Pig Manure and Sewage Sludge. Chemosphere 2019, 218, 624–631. [Google Scholar] [CrossRef]
- Gui, X.; Liu, C.; Li, F.; Wang, J. Effect of Pyrolysis Temperature on the Composition of DOM in Manure-Derived Biochar. Ecotoxicol. Environ. Saf. 2020, 197, 110597. [Google Scholar] [CrossRef]
- Uchimiya, M.; Ohno, T.; He, Z. Pyrolysis Temperature-Dependent Release of Dissolved Organic Carbon from Plant, Manure, and Biorefinery Wastes. J. Anal. Appl. Pyrolysis 2013, 104, 84–94. [Google Scholar] [CrossRef]
- Wei, J.; Tu, C.; Yuan, G.; Bi, D.; Wang, H.; Zhang, L.; Theng, B.K.G. Pyrolysis Temperature-Dependent Changes in the Characteristics of Biochar-Borne Dissolved Organic Matter and Its Copper Binding Properties. Bull Env. Contam Toxicol 2019, 103, 169–174. [Google Scholar] [CrossRef]
- Sun, T.; Sun, Y.; Huang, Q.; Xu, Y.; Jia, H. Sustainable Exploitation and Safe Utilization of Biochar: Multiphase Characterization and Potential Hazard Analysis. Bioresour. Technol. 2023, 383, 129241. [Google Scholar] [CrossRef]
- Jiang, S.; Dai, G.; Liu, Z.; He, T.; Zhong, J.; Ma, Y.; Shu, Y. Field-Scale Fluorescence Fingerprints of Biochar-Derived Dissolved Organic Matter (DOM) Provide an Effective Way to Trace Biochar Migration and the Downward Co-Migration of Pb, Cu and As in Soil. Chemosphere 2022, 301, 134738. [Google Scholar] [CrossRef]
- Wei, J.; Tu, C.; Yuan, G.; Zhou, Y.; Wang, H.; Lu, J. Limited Cu(II) Binding to Biochar DOM: Evidence from C K-Edge NEXAFS and EEM-PARAFAC Combined with Two-Dimensional Correlation Analysis. Sci. Total Environ. 2020, 701, 134919. [Google Scholar] [CrossRef]
- Wang, M.; Hu, R.; Zhao, J.; Kuzyakov, Y.; Liu, S. Iron Oxidation Affects Nitrous Oxide Emissions via Donating Electrons to Denitrification in Paddy Soils. Geoderma 2016, 271, 173–180. [Google Scholar] [CrossRef]
- Stookey, L.L. Ferrozine---a New Spectrophotometric Reagent for Iron. Anal. Chem. 1970, 42, 779–781. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Rapid Assay for Microbially Reducible Ferric Iron in Aquatic Sediments. Appl. Environ. Microbiol. 1987, 53, 1536–1540. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Y.; Liu, R.; Yin, P.; Li, D.; Shao, L. Speciation and Source Changes of Atmospheric Arsenic in Qingdao from 2016 to 2020—Response to Control Policies in China. Chemosphere 2023, 313, 137438. [Google Scholar] [CrossRef]
- Wenzel, W.W.; Kirchbaumer, N.; Prohaska, T.; Stingeder, G.; Lombi, E.; Adriano, D.C. Arsenic Fractionation in Soils Using an Improved Sequential Extraction Procedure. Anal. Chim. Acta 2001, 436, 309–323. [Google Scholar] [CrossRef]
- Sheng, Y.; Zhu, L. Biochar Alters Microbial Community and Carbon Sequestration Potential across Different Soil pH. Sci. Total Environ. 2018, 622–623, 1391–1399. [Google Scholar] [CrossRef]
- Jenkins, J.R.; Viger, M.; Arnold, E.C.; Harris, Z.M.; Ventura, M.; Miglietta, F.; Girardin, C.; Edwards, R.J.; Rumpel, C.; Fornasier, F.; et al. Biochar Alters the Soil Microbiome and Soil Function: Results of Next-generation Amplicon Sequencing across Europe. GCB Bioenergy 2017, 9, 591–612. [Google Scholar] [CrossRef]
- Cibati, A.; Foereid, B.; Bissessur, A.; Hapca, S. Assessment of Miscanthus × Giganteus Derived Biochar as Copper and Zinc Adsorbent: Study of the Effect of Pyrolysis Temperature, pH and Hydrogen Peroxide Modification. J. Clean. Prod. 2017, 162, 1285–1296. [Google Scholar] [CrossRef]
- Yuan, J.-H.; Xu, R.-K.; Zhang, H. The Forms of Alkalis in the Biochar Produced from Crop Residues at Different Temperatures. Bioresour. Technol. 2011, 102, 3488–3497. [Google Scholar] [CrossRef]
- Kluepfel, L.; Keiluweit, M.; Kleber, M.; Sander, M. Redox Properties of Plant Biomass-Derived Black Carbon (Biochar). Environ. Sci. Technol. 2014, 48, 5601–5611. [Google Scholar] [CrossRef]
- Abbas, G.; Murtaza, B.; Bibi, I.; Shahid, M.; Niazi, N.K.; Khan, M.I.; Amjad, M.; Hussain, M.; Natasha. Arsenic Uptake, Toxicity, Detoxification, and Speciation in Plants: Physiological, Biochemical, and Molecular Aspects. Int. J. Environ. Res. Public Health 2018, 15, 59. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, W.; Yao, D.; Wang, N.; Liu, M.; Wang, L.; Tian, W.; Yan, P.; Huang, Z.; Wang, H. Arsenic Pollution from Human Activities Drives Changes in Soil Microbial Community Characteristics. Environ. Microbiol. 2023, 25, 2592–2603. [Google Scholar] [CrossRef]
- Xie, X.; Gu, S.; Hao, L.; Zhang, T.; Guo, Z. Rhizosphere Microbial Communities and Geochemical Constraining Mechanism of Antimony Mine Waste-Adapted Plants in Southwestern China. Microorganisms 2022, 10, 1507. [Google Scholar] [CrossRef]
- Handley, K.M.; Héry, M.; Lloyd, J.R. Redox Cycling of Arsenic by the Hydrothermal Marine Bacterium Marinobacter Santoriniensis. Environ. Microbiol. 2009, 11, 1601–1611. [Google Scholar] [CrossRef]
- Luo, J.; Bai, Y.; Liang, J.; Qu, J. Metagenomic Approach Reveals Variation of Microbes with Arsenic and Antimony Metabolism Genes from Highly Contaminated Soil. PLOS ONE 2014, 9, e108185. [Google Scholar] [CrossRef]
- Engel, A.S.; Johnson, L.R.; Porter, M.L. Arsenite Oxidase Gene Diversity among Chloroflexi and Proteobacteria from El Tatio Geyser Field, Chile. FEMS Microbiol. Ecol. 2013, 83, 745–756. [Google Scholar] [CrossRef]
- Li, S.; Fan, S.; Peng, X.; Zheng, D.; Li, D. Using Ferrous-Oxidizing Bacteria to Enhance the Performance of a pH Neutral All-Iron Flow Battery. iScience 2024, 27, 108595. [Google Scholar] [CrossRef]
- Kappler, A.; Bryce, C.; Mansor, M.; Lueder, U.; Byrne, J.M.; Swanner, E.D. An Evolving View on Biogeochemical Cycling of Iron. Nat. Rev. Microbiol. 2021, 19, 360–374. [Google Scholar] [CrossRef]
- Dequiedt, S.; Saby, N.P.A.; Lelievre, M.; Jolivet, C.; Thioulouse, J.; Toutain, B.; Arrouays, D.; Bispo, A.; Lemanceau, P.; Ranjard, L. Biogeographical Patterns of Soil Molecular Microbial Biomass as Influenced by Soil Characteristics and Management. Glob. Ecol. Biogeogr. 2011, 20, 641–652. [Google Scholar] [CrossRef]
- Terrat, S.; Horrigue, W.; Dequietd, S.; Saby, N.P.A.; Lelièvre, M.; Nowak, V.; Tripied, J.; Régnier, T.; Jolivet, C.; Arrouays, D.; et al. Mapping and Predictive Variations of Soil Bacterial Richness across France. PLOS ONE 2017, 12, e0186766. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Wong, J.T.F.; Hashimoto, Y.; Huang, L.; Rinklebe, J.; Chang, S.X.; Bolan, N.; Wang, H.; Ok, Y.S. Response of Microbial Communities to Biochar-Amended Soils: A Critical Review. Biochar 2019, 1, 3–22. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Kiikkilä, O.; Fritze, H. Charcoal as a Habitat for Microbes and Its Effect on the Microbial Community of the Underlying Humus. Oikos 2000, 89, 231–242. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Y.; Xia, D.; Jiang, X.; Fu, D.; Shen, L.; Wang, H.; Li, Q.B. Enhanced Bioreduction of Iron and Arsenic in Sediment by Biochar Amendment Influencing Microbial Community Composition and Dissolved Organic Matter Content and Composition. J. Hazard. Mater. 2016, 311, 20–29. [Google Scholar] [CrossRef]
- Xu, M.; Gao, P.; Wu, J.; Ma, J.; Zhang, X.; Yang, G.; Long, L.; Chen, C.; Song, C.; Xiao, Y. Biochar Promotes Arsenic Sequestration on Iron Plaques and Cell Walls in Rice Roots. Chemosphere 2022, 288, 132422. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.M.; Bibi, I.; Niazi, N.K.; Shahid, M.; Iqbal, J.; Shakoor, M.B.; Ahmad, A.; Shah, N.S.; Bhattacharya, P.; Mao, K.; et al. Arsenic Biogeochemical Cycling in Paddy Soil-Rice System: Interaction with Various Factors, Amendments and Mineral Nutrients. Sci. Total Environ. 2021, 773, 145040. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Samanta, S.; Sarkar, S.; Mukherjee, A.; Pattnaik, B.K.; Saha, S.; Awasthi, J.P.; Bhowmick, S.; Ghosh, D.; Samal, A.C.; et al. Arsenic Contamination, Impact and Mitigation Strategies in Rice Agro-Environment: An Inclusive Insight. Sci. Total Environ. 2021, 800, 149477. [Google Scholar] [CrossRef]
- Yanan, Z.; Li, H.; Kai, Y.; Yiqun, G. The Role of Dissolved Organic Matter in the Competitive Adsorption to Goethite, during Arsenic Mobilization. Procedia Earth Planet. Sci. 2017, 17, 424–427. [Google Scholar] [CrossRef]
- Verbeeck, M.; Thiry, Y.; Smolders, E. Soil Organic Matter Affects Arsenic and Antimony Sorption in Anaerobic Soils. Environ. Pollut. 2020, 257, 113566. [Google Scholar] [CrossRef]
- Majumder, S.; Nath, B.; Sarkar, S.; Chatterjee, D.; Roman-Ross, G.; Hidalgo, M. Size-Fractionation of Groundwater Arsenic in Alluvial Aquifers of West Bengal, India: The Role of Organic and Inorganic Colloids. Sci. Total Environ. 2014, 468–469, 804–812. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, H.-M.; Yao, B.-M.; Chen, S.-C.; Sun, G.-X.; Zhu, Y.-G. Bioavailable Arsenic and Amorphous Iron Oxides Provide Reliable Predictions for Arsenic Transfer in Soil-Wheat System. J. Hazard. Mater. 2020, 383, 121160. [Google Scholar] [CrossRef]
- Al-Abed, S.R.; Jegadeesan, G.; Purandare, J.; Allen, D. Arsenic Release from Iron Rich Mineral Processing Waste: Influence of pH and Redox Potential. Chemosphere 2007, 66, 775–782. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Nakamura, T.; Dong, D.; Takahashi, Y.; Amachi, S.; Makino, T. Arsenic Release from Flooded Paddy Soils Is Influenced by Speciation, Eh, pH, and Iron Dissolution. Chemosphere 2011, 83, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Ascar, L.; Ahumada, I.; Richter, P. Influence of Redox Potential (Eh) on the Availability of Arsenic Species in Soils and Soils Amended with Biosolid. Chemosphere 2008, 72, 1548–1552. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.S.; Shafiquzzaman, M.; Haider, H. Arsenic Release Dynamics of Paddy Field Soil during Groundwater Irrigation and Natural Flooding. J. Environ. Manag. 2023, 343, 118204. [Google Scholar] [CrossRef] [PubMed]
- Rinklebe, J.; Shaheen, S.M.; Yu, K. Release of As, Ba, Cd, Cu, Pb, and Sr under Pre-Definite Redox Conditions in Different Rice Paddy Soils Originating from the U.S.A. and Asia. Geoderma 2016, 270, 21–32. [Google Scholar] [CrossRef]
- Aftabtalab, A.; Rinklebe, J.; Shaheen, S.M.; Niazi, N.K.; Moreno-Jiménez, E.; Schaller, J.; Knorr, K.-H. Review on the Interactions of Arsenic, Iron (Oxy)(Hydr)Oxides, and Dissolved Organic Matter in Soils, Sediments, and Groundwater in a Ternary System. Chemosphere 2022, 286, 131790. [Google Scholar] [CrossRef]
- Aftabtalab, A.; Moreno-Jiménez, E.; Henschel, J.; Nowak, S.; Schaller, J.; Knorr, K.-H. The Impact of Dissolved Organic Matter on Arsenic Mobilization from Goethite in the Presence of Silicic Acid and Phosphate under Reducing Conditions. Water 2022, 14, 2975. [Google Scholar] [CrossRef]
- Buschmann, J.; Kappeler, A.; Lindauer, U.; Kistler, D.; Berg, M.; Sigg, L. Arsenite and Arsenate Binding to Dissolved Humic Acids: Influence of pH, Type of Humic Acid, and Aluminum. Environ. Sci. Technol. 2006, 40, 6015–6020. [Google Scholar] [CrossRef]
- Fan, Y.; Zheng, C.; Liu, H.; He, C.; Shen, Z.; Zhang, T.C. Effect of pH on the Adsorption of Arsenic(V) and Antimony(V) by the Black Soil in Three Systems: Performance and Mechanism. Ecotoxicol. Environ. Saf. 2020, 191, 110145. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Sarkar, B.; Barman, A.; Datta, S.C.; Manjaiah, K.M. Arsenic Adsorption on Modified Clay Minerals in Contaminated Soil and Water: Impact of pH and Competitive Anions. CLEAN—Soil Air Water 2021, 49, 2000259. [Google Scholar] [CrossRef]
- Bai, Y.; Ding, A.; Zhang, S. Characterizing chromophoric dissolved organic matter in Guanting Reservoir Beijing using excitation-emission matrix fluorescence and parallel factor analysis (No. EGU2020-17510). In Proceedings of the EGU2020, Copernicus Meetings, Online, 4–8 May 2020. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, Y.; Wang, H.; Lu, W.; Zhou, Z.; Zhang, Y.; Ren, L. Influence of pyrolysis temperature on characteristics and heavy metal adsorptive performance of biochar derived from municipal sewage sludge. Bioresour. Technol. 2014, 164, 47–54. [Google Scholar] [CrossRef]
- Ding, W.; Dong, X.; Ime, I.M.; Gao, B.; Ma, L.Q. Pyrolytic temperatures impact lead sorption mechanisms by bagasse biochars. Chemosphere 2014, 105, 68–74. [Google Scholar] [CrossRef]
- Huo, P.; Zhang, W.; Jia, H.; Chen, J.; Gao, P. Characteristics of optical properties of DOM and nutrients in rainwater of different ecological areas of a large reservoir in China. Chemosphere 2023, 342, 140091. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Ok, Y.S.; Hur, J. Dynamic variations in dissolved organic matter and the precursors of disinfection by-products leached from biochars: Leaching experiments simulating intermittent rain events. Environ. Pollut. 2018, 242, 1912–1920. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Wang, F.; Wang, L.; Liu, J.; Hashimoto, Y.; Hosomi, M. Arsenic immobilization and removal in contaminated soil using zero-valent iron or magnetic biochar amendment followed by dry magnetic separation. Sci. Total Environ. 2021, 768, 144521. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-P.; Liu, Y.-H.; Ren, D.; Wang, J.-J. Characteristics and chlorine reactivity of biochar-derived dissolved organic matter: Effects of feedstock type and pyrolysis temperature. Water Res. 2022, 211, 118044. [Google Scholar] [CrossRef]
- Ronsse, F.; van Hecke, S.; Dickinson, D.; Prins, W. Production and characterization of slow pyrolysis biochar: Influence of feedstock type and pyrolysis conditions. GCB Bioenergy 2013, 5, 104–115. [Google Scholar] [CrossRef]
- Spokas, K.A.; Cantrell, K.B.; Novak, J.M.; Archer, D.W.; Ippolito, J.A.; Collins, H.P.; Boateng, A.A.; Lima, I.M.; Lamb, M.C.; McAloon, A.J.; et al. Biochar: A Synthesis of Its Agronomic Impact beyond Carbon Sequestration. J. Environ. Qual. 2012, 41, 973–989. [Google Scholar] [CrossRef]
- Tang, J.; Li, X.; Luo, Y.; Li, G.; Khan, S. Spectroscopic characterization of dissolved organic matter derived from different biochars and their polycylic aromatic hydrocarbons (PAHs) binding affinity. Chemosphere 2016, 152, 399–406. [Google Scholar] [CrossRef]
- Tomczyk, A.; Sokołowska, Z.; Boguta, P. Biochar physicochemical properties: Pyrolysis temperature and feedstock kind effects. Rev. Environ. Sci. Biotechnol. 2020, 19, 191–215. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Shen, Z.; Feng, C.; Chen, J. Revealing Sources and Distribution Changes of Dissolved Organic Matter (DOM) in Pore Water of Sediment from the Yangtze Estuary. PLoS ONE 2013, 8, e76633. [Google Scholar] [CrossRef]
- Wen, Y.; Xiao, M.; Chen, Z.; Zhang, W.; Yue, F. Seasonal Variations of Dissolved Organic Matter in Urban Rivers of Northern China. Land 2023, 12, 273. [Google Scholar] [CrossRef]
- Wu, H.; Qi, Y.; Dong, L.; Zhao, X.; Liu, H. Revealing the impact of pyrolysis temperature on dissolved organic matter released from the biochar prepared from Typha orientalis. Chemosphere 2019, 228, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, P.; Yuan, X.; Li, Y.; Han, L. Effect of pyrolysis temperature and correlation analysis on the yield and physicochemical properties of crop residue biochar. Bioresour. Technol. 2020, 296, 122318. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.-X.; Ta, N.; Wang, X.-D. Effect of Temperature on the Structural and Physicochemical Properties of Biochar with Apple Tree Branches as Feedstock Material. Energies 2017, 10, 1293. [Google Scholar] [CrossRef]
- Zoroufchi Benis, K.; Motalebi Damuchali, A.; Soltan, J.; McPhedran, K.N. Treatment of aqueous arsenic—A review of biochar modification methods. Sci. Total Environ. 2020, 739, 139750. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Liao, M.; Duan, T.; Hu, Y.; Sun, J. The Mechanism of Arsenic Release in Contaminated Paddy Soil with Added Biochar: The Role of Dissolved Organic Matter, Fe, and Bacteria. Toxics 2024, 12, 661. https://doi.org/10.3390/toxics12090661
Fan J, Liao M, Duan T, Hu Y, Sun J. The Mechanism of Arsenic Release in Contaminated Paddy Soil with Added Biochar: The Role of Dissolved Organic Matter, Fe, and Bacteria. Toxics. 2024; 12(9):661. https://doi.org/10.3390/toxics12090661
Chicago/Turabian StyleFan, Jianxin, Maoyu Liao, Ting Duan, Ying Hu, and Jiaoxia Sun. 2024. "The Mechanism of Arsenic Release in Contaminated Paddy Soil with Added Biochar: The Role of Dissolved Organic Matter, Fe, and Bacteria" Toxics 12, no. 9: 661. https://doi.org/10.3390/toxics12090661
APA StyleFan, J., Liao, M., Duan, T., Hu, Y., & Sun, J. (2024). The Mechanism of Arsenic Release in Contaminated Paddy Soil with Added Biochar: The Role of Dissolved Organic Matter, Fe, and Bacteria. Toxics, 12(9), 661. https://doi.org/10.3390/toxics12090661