


Exploring Neurobehaviour in Zebrafish Embryos as a Screening Model for Addictiveness of Substances

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Housing and Maintenance
2.3. Zebrafish Embryo Toxicity Test
2.4. Chemical Analysis
2.5. Locomotion Behavior in the Zebrabox
2.6. Data Analysis and Statistics
3. Results
3.1. Embryotoxicity of Nicotine (ZFET)
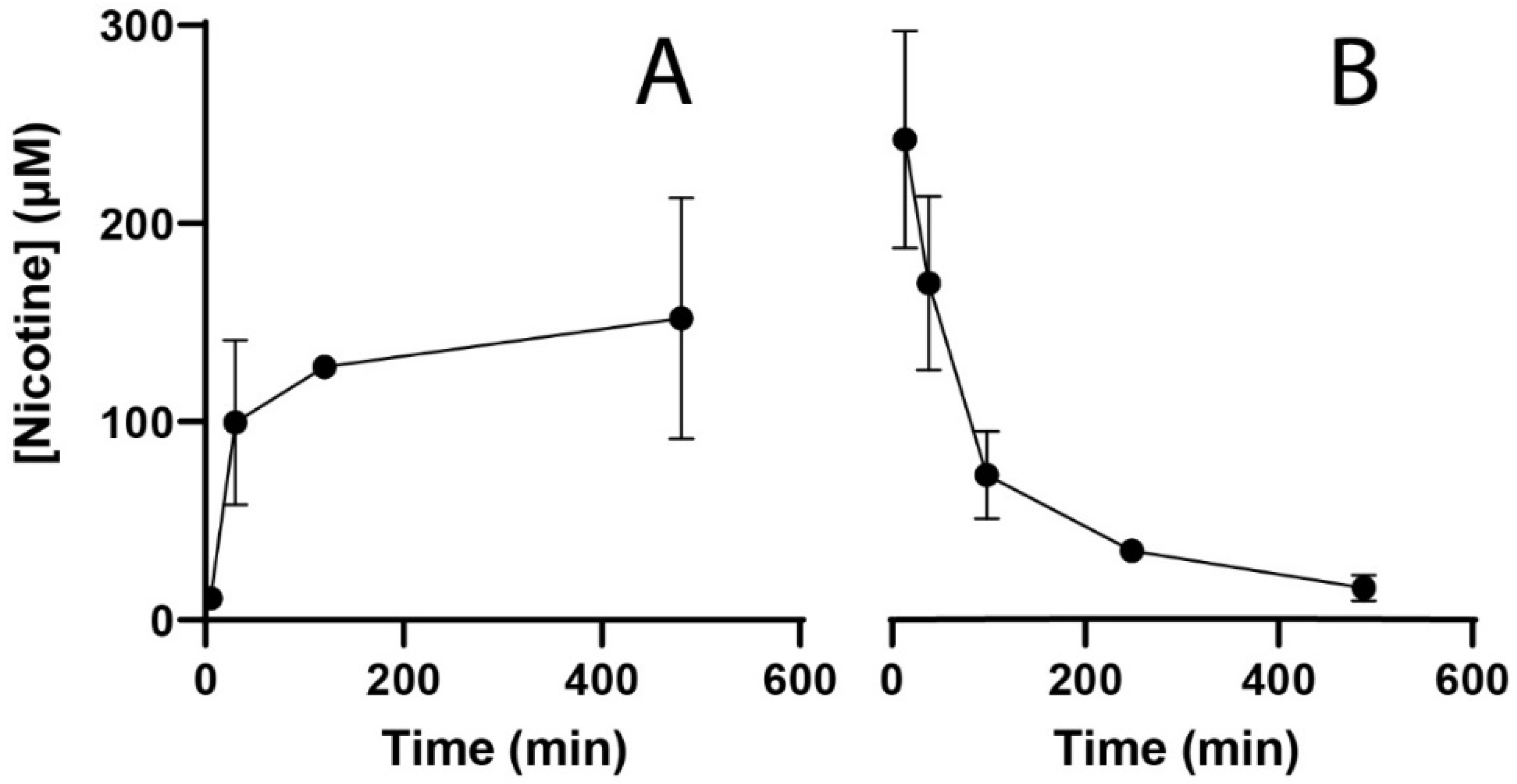
3.2. Kinetics of Nicotine in Zebrafish Embryos
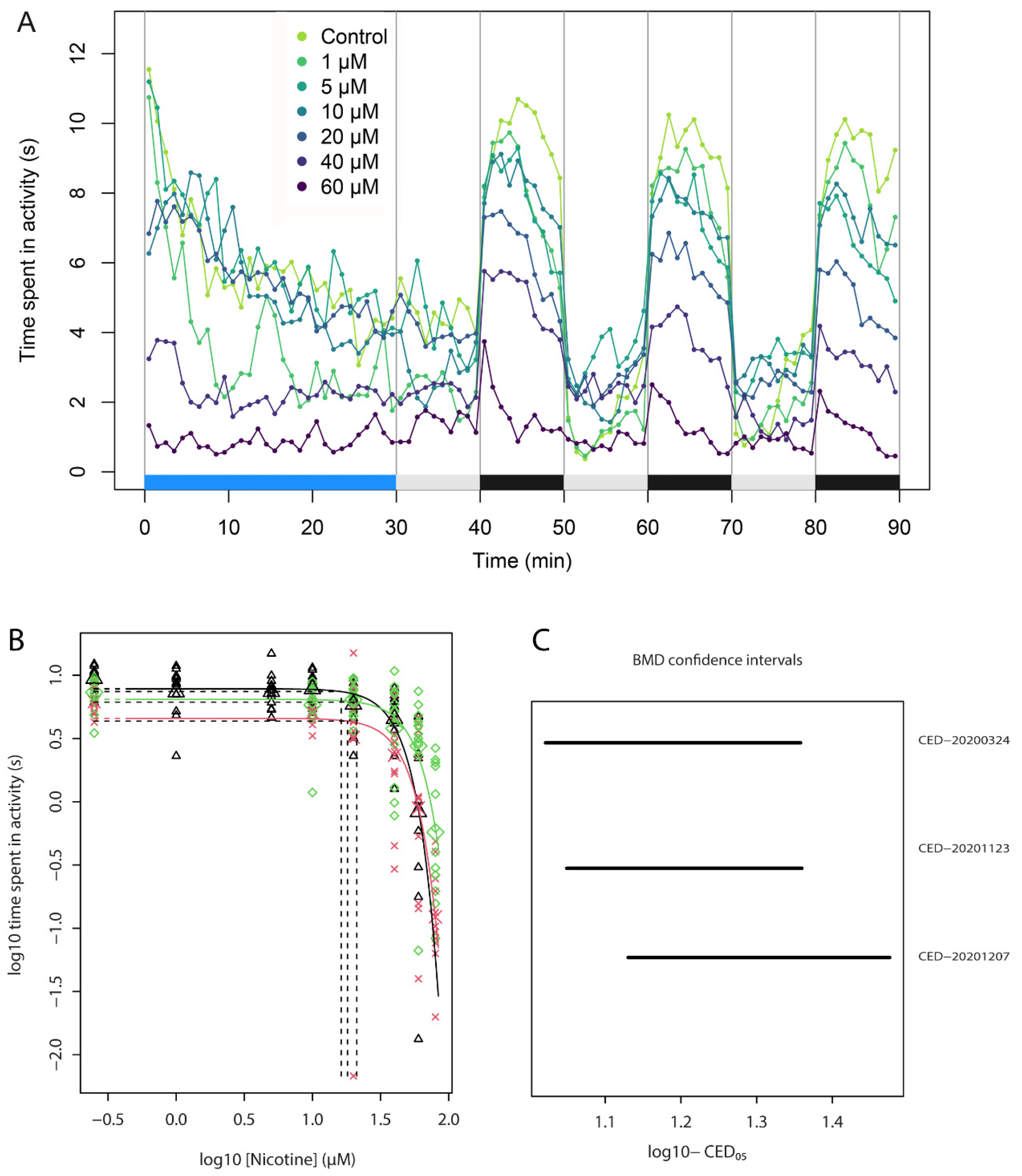
3.3. Locomotion in the Zebrabox
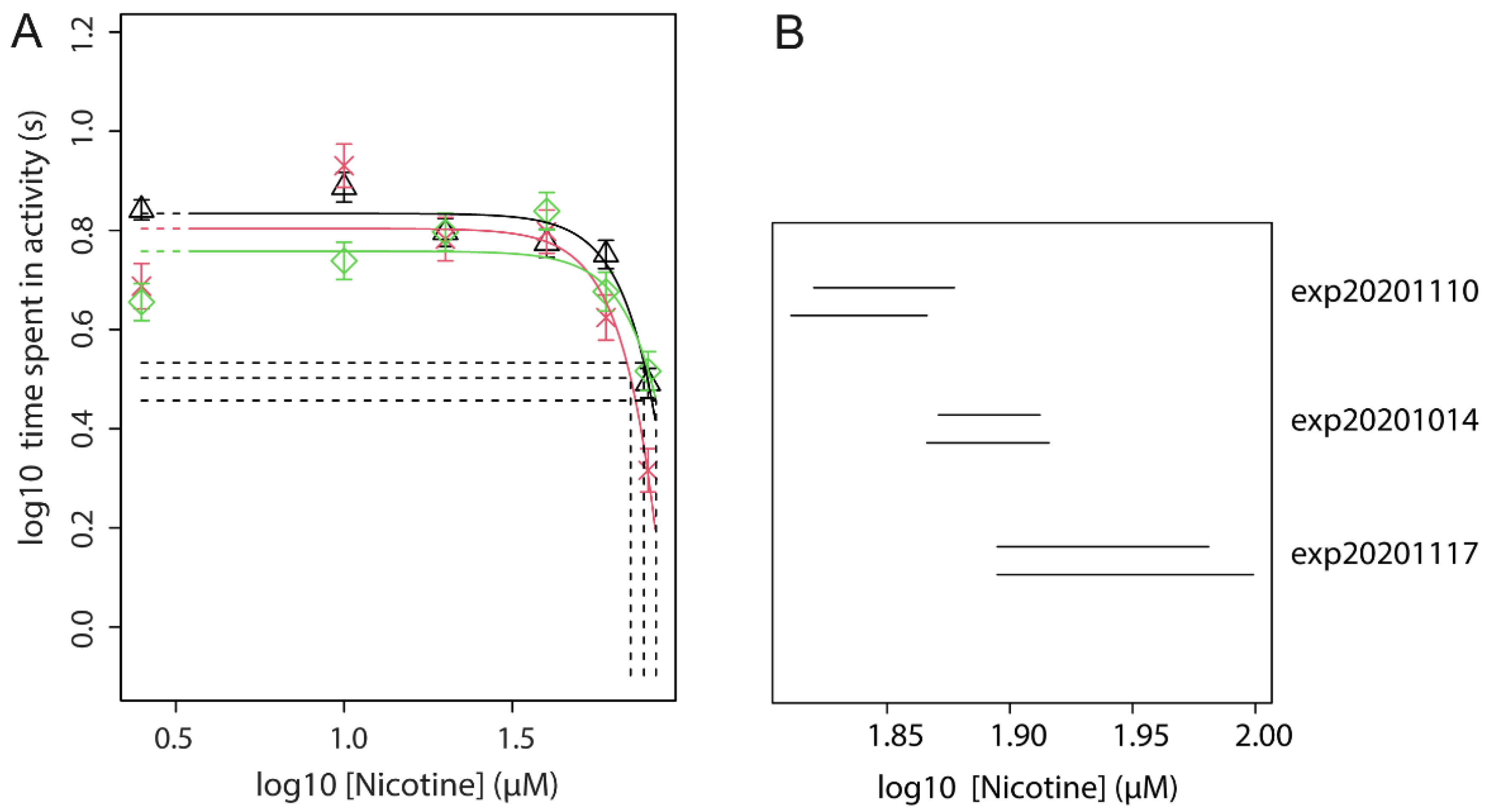
3.4. Concentration-Response of Acute Nicotine
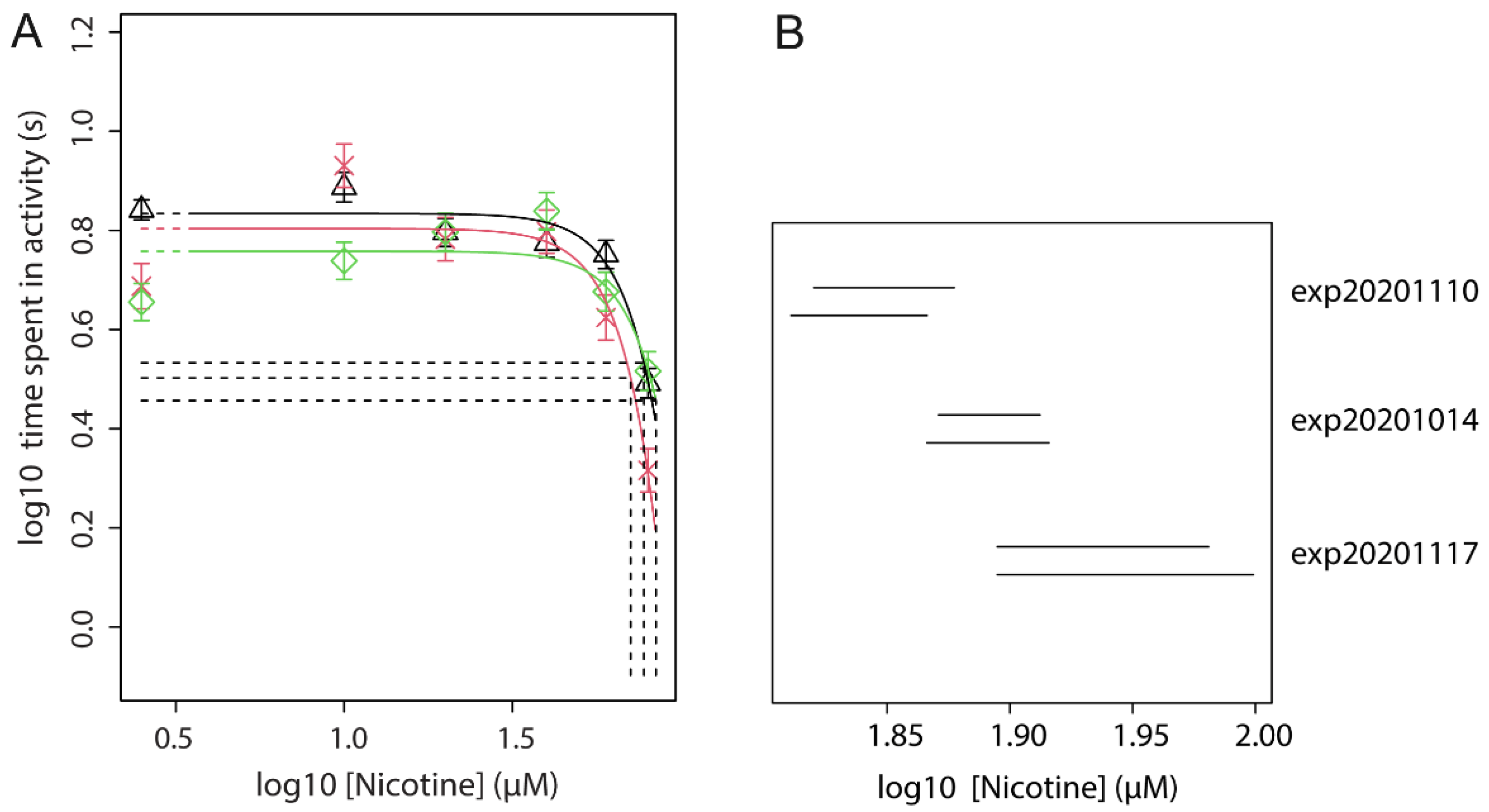
3.5. Effect of Nicotine Pre-Treatment (Concentration Range) on Acute Nicotine Exposure (Fixed Concentration)
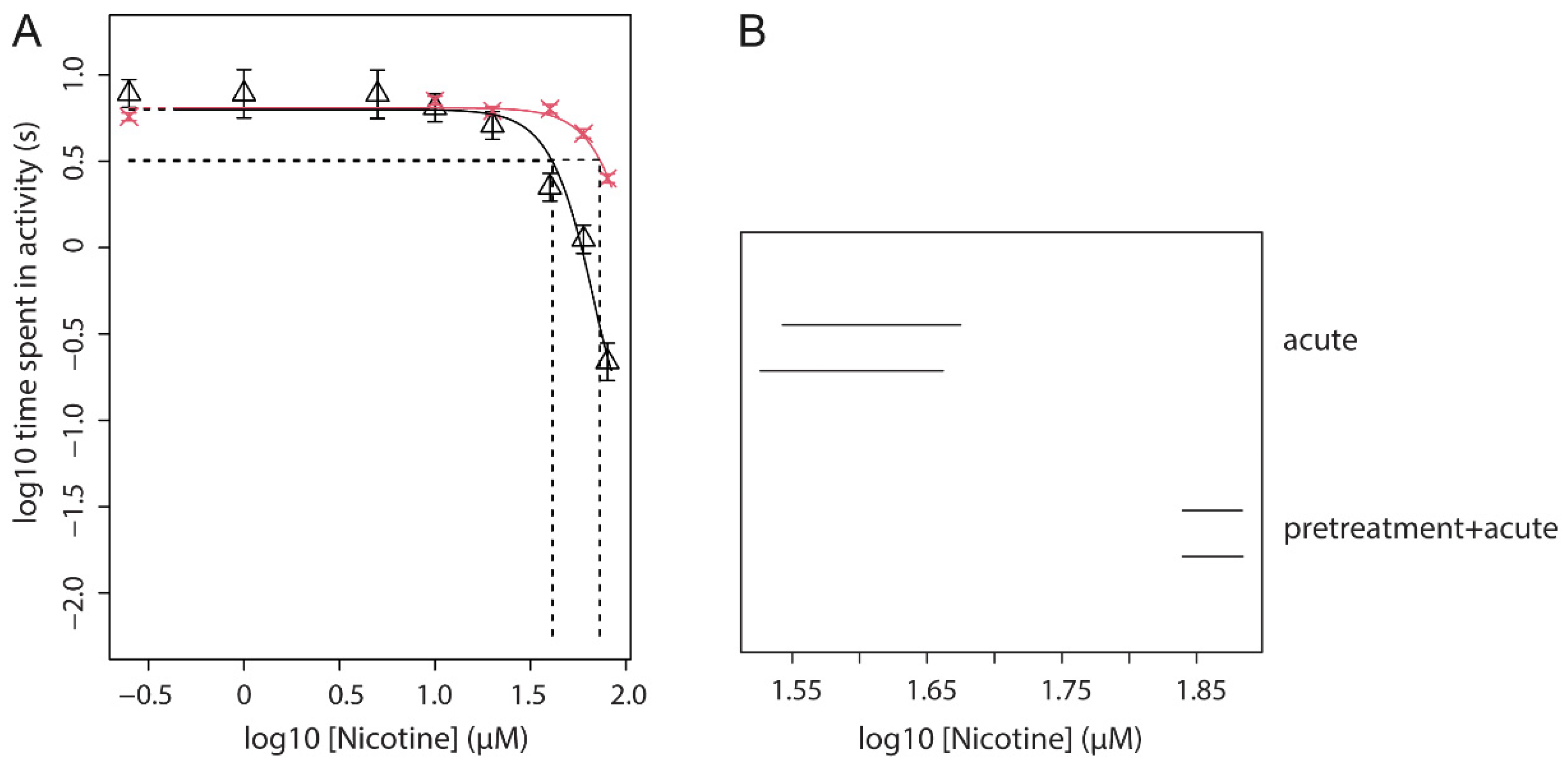
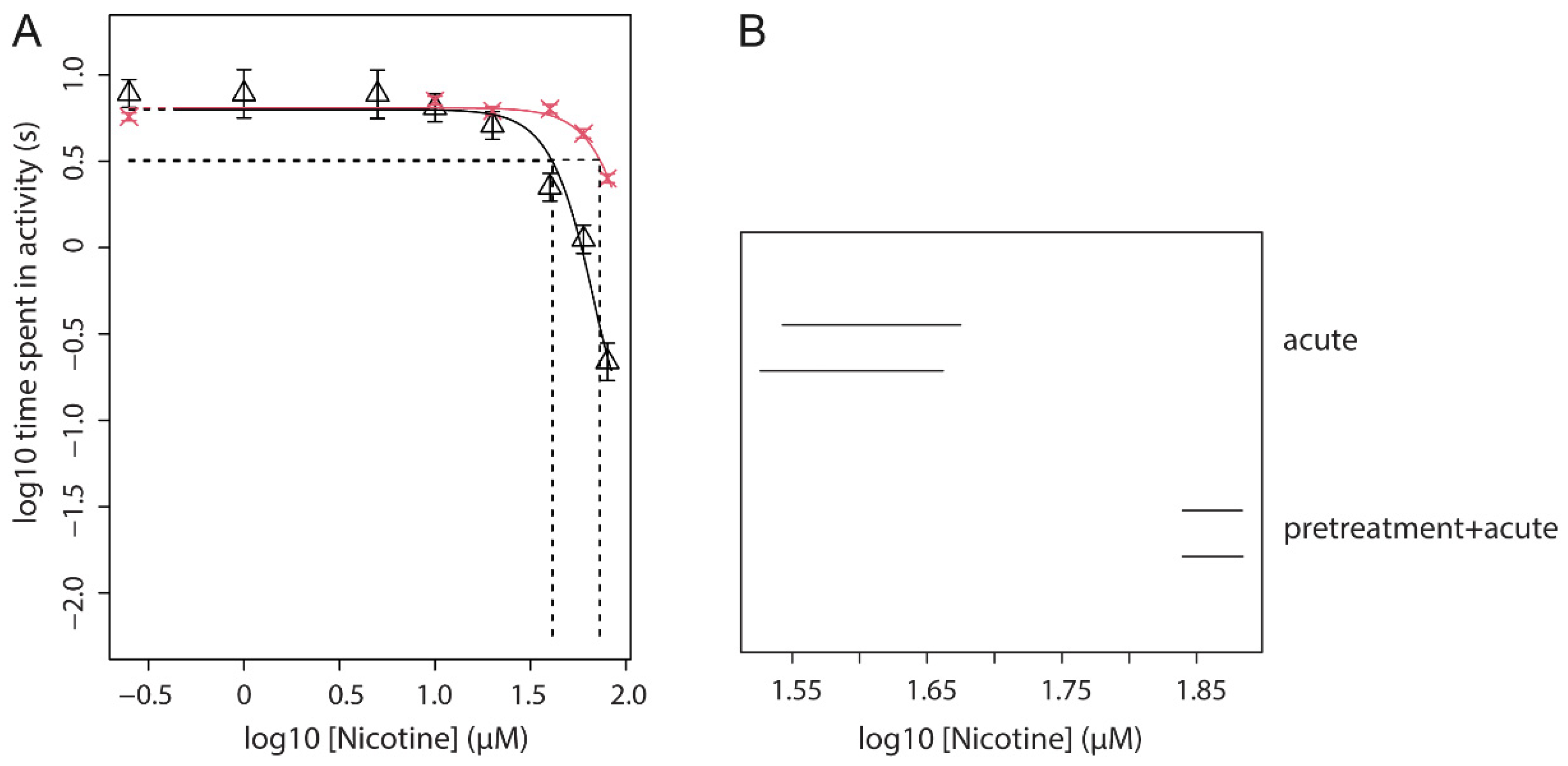
3.6. Effect of Nicotine Pre-Treatment (Fixed Dose) on Acute Nicotine Exposure (Dose Range)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- U.S. Department of Health and Human Services. The Health Consequences of Smoking: 50 Years of Progress. A Report of the Surgeon General; U.S. Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Office on Smoking and Health: Atlanta, GA, USA, 2014.
- World Health Organization (WHO). WHO Report on the Global Tobacco Epidemic: Addressing New and Emerging Products; World Health Organization (WHO): Geneva, Switzerland, 2021. [Google Scholar]
- Hughes, J.R.; Keely, J.; Naud, S. Shape of the relapse curve and long-term abstinence among untreated smokers. Addiction 2004, 99, 29–38. [Google Scholar] [CrossRef]
- Ursprung, W.W.S.A.; DiFranza, J.R. The loss of autonomy over smoking in relation to lifetime cigarette consumption. Addict. Behav. 2010, 35, 14–18. [Google Scholar] [CrossRef] [PubMed]
- van de Nobelen, S.; Kienhuis, A.S.; Talhout, R. An Inventory of Methods for the Assessment of Additive Increased Addictiveness of Tobacco Products: Table 1. Nicotine Tob. Res. 2016, 18, 1546–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinoff, M.; Caskey, N.; Rissling, A.; Park, C. Pharmacological and Chemical Effects of Cigarette Additives. Am. J. Public Health 2007, 97, 1981–1991. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, A.; Evans, S. Abuse potential of non-nicotine tobacco smoke components: Acetaldehyde, nornicotine, cotinine and anabasine. Nicotine Toabcco Res. 2013, 15, 622–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talhout, R.; Opperhuizen, A.; van Amsterdam, J.G. Sugars as tobacco ingredient: Effects on mainstream smoke composition. Food Chem. Toxicol. 2006, 44, 1789–1798. [Google Scholar] [CrossRef]
- The European Parliament and the Council of the European Union. Tobacco Product Directive 2014/40/EU of the European Parliament and of the Council; The European Union: Brussels, Belgium, 2014; Volume L127, pp. 1–38. [Google Scholar]
- Benowitz, N.L.; Henningfield, J.E. Nicotine Reduction Strategy: State of the science and challenges to tobacco control policy and FDA tobacco product regulation. Prev. Med. 2018, 117, 5–7. [Google Scholar] [CrossRef]
- Corrigall, W.A. Nicotine self-administration in animals as a dependence model. Nicotine Tob. Res. 1999, 1, 11–20. [Google Scholar] [CrossRef]
- O’Dell, L.E.; Khroyan, T.V. Rodent models of nicotine reward: What do they tell us about tobacco abuse in humans? Pharmacol. Biochem. Behav. 2009, 91, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Scientific Committee on Emerging and Newly Identified Health Risks (SCENIHR). Opinion on Additives Used in Tobacco Products (Opinion 1), Tobacco Additives I; The European Union: Brussels, Belgium, 2016. [Google Scholar]
- Nederlandse Wet op de Dierproeven (WoD). Available online: https://wetten.overheid.nl/BWBR0003081/2021-07-01 (accessed on 25 March 2021).
- Selderslaghs, I.W.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicology Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef]
- Tran, S.; Gerlai, R. Recent advances with a novel model organism: Alcohol tolerance and sensitization in zebrafish (Danio rerio). Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irons, T.; Kelly, P.; Hunter, D.; MacPhail, R.; Padilla, S. Acute administration of dopaminergic drugs has differential effects on locomotion in larval zebrafish. Pharmacol. Biochem. Behav. 2013, 103, 792–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irons, T.; MacPhail, R.; Hunter, D.; Padilla, S. Acute neuroactive drug exposures alter locomotor activity in larval zebrafish. Neurotoxicology Teratol. 2010, 32, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Basnet, R.M.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish Larvae as a Behavioral Model in Neuropharmacology. Biomedicines 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Parliament and the Council of the European Union. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; The European Union: Brussels, Belgium, 2010; Volume L276, pp. 33–79. [Google Scholar]
- Hubrecht, R.C.; Carter, E. The 3Rs and Humane Experimental Technique: Implementing Change. Animals 2019, 9, 754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horzmann, K.A.; Freeman, J.L. Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity. Toxics 2016, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kily, L.J.M.; Cowe, Y.C.M.; Hussain, O.; Patel, S.; McElwaine, S.; Cotter, F.E.; Brennan, C.H. Gene expression changes in a zebrafish model of drug dependency suggest conservation of neuro-adaptation pathways. J. Exp. Biol. 2008, 211, 1623–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benowitz, M.D. Neurobiology of Nicotine Addiction: Implications for Smoking Cessation Treatment. Am. J. Med. 2008, 121, S3–S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dani, J.A. Neuronal Nicotinic Acetylcholine Receptor Structure and Function and Response to Nicotine. Int. Rev. Neurobiol. 2015, 124, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, H.; Granato, M. Modulation of locomotor activity in larval zebrafish during light adaptation. J. Exp. Biol. 2007, 210, 2526–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atzei, A.; Jense, I.; Zwart, E.; Legradi, J.; Venhuis, B.; van der Ven, L.; Heusinkveld, H.; Hessel, E. Developmental Neurotoxicity of Environmentally Relevant Pharmaceuticals and Mixtures Thereof in a Zebrafish Embryo Behavioural Test. Int. J. Environ. Res. Public Health 2021, 18, 6717. [Google Scholar] [CrossRef] [PubMed]
- Vossen, L.E.; Cerveny, D.; Österkrans, M.; Thörnqvist, P.-O.; Jutfelt, F.; Fick, J.; Brodin, T.; Winberg, S. Chronic Exposure to Oxazepam Pollution Produces Tolerance to Anxiolytic Effects in Zebrafish (Danio rerio). Environ. Sci. Technol. 2020, 54, 1760–1769. [Google Scholar] [CrossRef] [PubMed]
- Hermsen, S.A.; Brandhof, E.-J.V.D.; van der Ven, L.T.; Piersma, A.H. Relative embryotoxicity of two classes of chemicals in a modified zebrafish embryotoxicity test and comparison with their in vivo potencies. Toxicol. Vitr. 2011, 25, 745–753. [Google Scholar] [CrossRef] [Green Version]
- Brox, S.; Seiwert, B.; Küster, E.; Reemtsma, T. Toxicokinetics of Polar Chemicals in Zebrafish Embryo (Danio rerio): Influence of Physicochemical Properties and of Biological Processes. Environ. Sci. Technol. 2016, 50, 10264–10272. [Google Scholar] [CrossRef]
- Slob, W. Dose-Response Modeling of Continuous Endpoints. Toxicol. Sci. 2002, 66, 298–312. [Google Scholar] [CrossRef]
- Mora-Zamorano, F.X.; Svoboda, K.R.; Carvan, M.J. The Nicotine-Evoked Locomotor Response: A Behavioral Paradigm for Toxicity Screening in Zebrafish (Danio rerio) Embryos and Eleutheroembryos Exposed to Methylmercury. PLoS ONE 2016, 11, e0154570. [Google Scholar] [CrossRef] [Green Version]
- Domino, E.F. Nicotine induced behavioral locomotor sensitization. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2001, 25, 59–71. [Google Scholar] [CrossRef]
- Adriani, W.; Macrì, S.; Pacifici, R.; Laviola, G. Peculiar Vulnerability to Nicotine Oral Self-administration in Mice during Early Adolescence. Neuropsychopharmacology 2002, 27, 212–224. [Google Scholar] [CrossRef]
- Rosecrans, J.A. The psychopharmacological basis of nicotine’s differential effects on behavior: Individual subject variability in the rat. Behav. Genet. 1995, 25, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Simpson, D.; White, N.; Randall, C. Age- and sex-related differences in alcohol and nicotine effects in C57BL/6J mice. Addict. Biol. 2003, 8, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.; Caruso, M.; Kamens, H. Early-adolescent male C57BL/6J and DBA/2J mice display reduced sensitivity to acute nicotine administration. Neurosci. Lett. 2019, 690, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Addy, N.A.; Mineur, Y.S.; Brunzell, D.H. It is not “either/or”: Activation and desensitization of nicotinic acetylcholine receptors both contribute to behaviors related to nicotine addiction and mood. Prog. Neurobiol. 2008, 84, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Ortells, M.O.; Arias, H.R. Neuronal networks of nicotine addiction. Int. J. Biochem. Cell Biol. 2010, 42, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, T.A.; Tate, C.A.; Cousins, M.M.; Seidler, F.J. Prenatal Nicotine Exposure Alters the Responses to Subsequent Nicotine Administration and Withdrawal in Adolescence: Serotonin Receptors and Cell Signaling. Neuropsychopharmacology 2006, 31, 2462–2475. [Google Scholar] [CrossRef] [Green Version]
- Shacka, J.J.; Fennell, O.B.; Robinson, S.E. Prenatal Nicotine Sex-Dependently Alters Agonist-Induced Locomotion and Stereotypy. Neurotoxicology Teratol. 1997, 19, 467–476. [Google Scholar] [CrossRef]
- Wang, T.; Chen, M.; Yan, Y.-E.; Xiao, F.-Q.; Pan, X.-L.; Wang, H. Growth retardation of fetal rats exposed to nicotinein utero: Possible involvement of CYP1A1, CYP2E1, and P-glycoprotein. Environ. Toxicol. 2009, 24, 33–42. [Google Scholar] [CrossRef]
- Holbrook, B.D. The effects of nicotine on human fetal development. Birth Defects Res. Part C Embryo Today Rev. 2016, 108, 181–192. [Google Scholar] [CrossRef]
- Svoboda, K.R.; Vijayaraghavan, S.; Tanguay, R.L. Nicotinic Receptors Mediate Changes in Spinal Motoneuron Development and Axonal Pathfinding in Embryonic Zebrafish Exposed to Nicotine. J. Neurosci. 2002, 22, 10731–10741. [Google Scholar] [CrossRef]
- Ellis, L.D.; Soo, E.C.; Achenbach, J.C.; Morash, M.G.; Soanes, K.H. Use of the Zebrafish Larvae as a Model to Study Cigarette Smoke Condensate Toxicity. PLoS ONE 2014, 9, e115305. [Google Scholar] [CrossRef]
- Papke, R.L.; Ono, F.; Stokes, C.; Urban, J.M.; Boyd, R.T. The nicotinic acetylcholine receptors of zebrafish and an evaluation of pharmacological tools used for their study. Biochem. Pharmacol. 2012, 84, 352–365. [Google Scholar] [CrossRef] [Green Version]
- Higa, K.K.; Grim, A.; Kamenski, M.E.; van Enkhuizen, J.; Zhou, X.; Li, K.; Naviaux, J.C.; Wang, L.; Naviaux, R.K.; Geyer, M.A.; et al. Nicotine withdrawal-induced inattention is absent in alpha7 nAChR knockout mice. Psychopharmacology 2017, 234, 1573–1586. [Google Scholar] [CrossRef] [Green Version]
- Salas, R.; Main, A.; Gangitano, D.; De Biasi, M. Decreased withdrawal symptoms but normal tolerance to nicotine in mice null for the α7 nicotinic acetylcholine receptor subunit. Neuropharmacology 2007, 53, 863–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotti, C.; Zoli, M.; Clementi, F. Brain nicotinic acetylcholine receptors: Native subtypes and their relevance. Trends Pharmacol. Sci. 2006, 27, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Tapper, A.R.; McKinney, S.L.; Nashmi, R.; Schwarz, J.; Deshpande, P.; Labarca, C.; Whiteaker, P.; Marks, M.J.; Collins, A.C.; Lester, H.A. Nicotine Activation of 4* Receptors: Sufficient for Reward, Tolerance, and Sensitization. Science 2004, 306, 1029–1032. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.B.; Kraus, G.P. Physiology, Cholinergic Receptors; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2021. [Google Scholar]
- Zouridakis, M.; Zisimopoulou, P.; Poulas, K.; Tzartos, S.J. Recent advances in understanding the structure of nicotinic acetylcholine receptors. IUBMB Life 2009, 61, 407–423. [Google Scholar] [CrossRef] [PubMed]
- Kalamida, D.; Poulas, K.; Avramopoulou, V.; Fostieri, E.; Lagoumintzis, G.; Lazaridis, K.; Sideri, A.; Zouridakis, M.; Tzartos, S.J. Muscle and neuronal nicotinic acetylcholine receptors. FEBS J. 2007, 274, 3799–3845. [Google Scholar] [CrossRef] [PubMed]
- Mathis, V.; Kenny, P.J. From controlled to compulsive drug-taking: The role of the habenula in addiction. Neurosci. Biobehav. Rev. 2019, 106, 102–111. [Google Scholar] [CrossRef]
- Ponzoni, L.; Melzi, G.; Marabini, L.; Martini, A.; Petrillo, G.; Teh, M.-T.; Torres-Perez, J.V.; Morara, S.; Gotti, C.; Braida, D.; et al. Conservation of mechanisms regulating emotional-like responses on spontaneous nicotine withdrawal in zebrafish and mammals. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 111, 110334. [Google Scholar] [CrossRef]
- Miller, N.; Greene, K.; Dydinski, A.; Gerlai, R. Effects of nicotine and alcohol on zebrafish (Danio rerio) shoaling. Behav. Brain Res. 2013, 240, 192–196. [Google Scholar] [CrossRef]
- Stewart, A.M.; Grossman, L.; Collier, A.D.; Echevarria, D.J.; Kalueff, A.V. Anxiogenic-like effects of chronic nicotine exposure in zebrafish. Pharmacol. Biochem. Behav. 2015, 139, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.D.; Bencan, Z.; Cerutti, D.T. Anxiolytic effects of nicotine in zebrafish. Physiol. Behav. 2007, 90, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.D. Zebrafish assessment of cognitive improvement and anxiolysis: Filling the gap between in vitro and rodent models for drug development. Rev. Neurosci. 2011, 22, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Ziani, P.R.; Müller, T.E.; Stefanello, F.V.; Fontana, B.D.; Duarte, T.; Canzian, J.; Rosemberg, D.B. Nicotine increases fear responses and brain acetylcholinesterase activity in a context-dependent manner in zebrafish. Pharmacol. Biochem. Behav. 2018, 170, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.D.; Limpuangthip, J.; Rachakonda, T.; Peterson, M. Timing of nicotine effects on learning in zebrafish. Psychopharmacology 2006, 184, 547–552. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Experiment | Endpoint | Purpose | Result | |
|---|---|---|---|---|
| 0 | Toxicity testing (ZFET) | embryonal development | Find sub-toxic dose for further testing | Figure S1 |
| 1 | Dose-range testing of acute nicotine | locomotion | Find potency of acute nicotine and an effective dose for exp. 2 | Figure 2 |
| 2 | Dose-range of nicotine pre-treatment with a fixed effective dose of acute nicotine | locomotion | Find an effective pre-treatment dose for exp. 3 | Figure 3 |
| 3 | Fixed dose of nicotine pre-treatment with a dose-range of acute nicotine | locomotion | Find an effect of pre-treatment on the potency of acute nicotine | Figures 4 and 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havermans, A.; Zwart, E.P.; Cremers, H.W.J.M.; van Schijndel, M.D.M.; Constant, R.S.; Mešković, M.; Worutowicz, L.X.; Pennings, J.L.A.; Talhout, R.; van der Ven, L.T.M.; et al. Exploring Neurobehaviour in Zebrafish Embryos as a Screening Model for Addictiveness of Substances. Toxics 2021, 9, 250. https://doi.org/10.3390/toxics9100250
Havermans A, Zwart EP, Cremers HWJM, van Schijndel MDM, Constant RS, Mešković M, Worutowicz LX, Pennings JLA, Talhout R, van der Ven LTM, et al. Exploring Neurobehaviour in Zebrafish Embryos as a Screening Model for Addictiveness of Substances. Toxics. 2021; 9(10):250. https://doi.org/10.3390/toxics9100250
Chicago/Turabian StyleHavermans, Anne, Edwin P. Zwart, Hans W. J. M. Cremers, Maarten D. M. van Schijndel, Romy S. Constant, Maja Mešković, Laura X. Worutowicz, Jeroen L. A. Pennings, Reinskje Talhout, Leo T. M. van der Ven, and et al. 2021. "Exploring Neurobehaviour in Zebrafish Embryos as a Screening Model for Addictiveness of Substances" Toxics 9, no. 10: 250. https://doi.org/10.3390/toxics9100250
APA StyleHavermans, A., Zwart, E. P., Cremers, H. W. J. M., van Schijndel, M. D. M., Constant, R. S., Mešković, M., Worutowicz, L. X., Pennings, J. L. A., Talhout, R., van der Ven, L. T. M., & Heusinkveld, H. J. (2021). Exploring Neurobehaviour in Zebrafish Embryos as a Screening Model for Addictiveness of Substances. Toxics, 9(10), 250. https://doi.org/10.3390/toxics9100250