

Bark Morphology and Nutrient Flux in Urban Trees: Investigating Water Absorption and Ion Concentration Dynamics



Abstract
1. Introduction
2. Materials and Methods
2.1. Hydrological Processes and Ion Concentrations
2.2. Nutrients Input
2.3. Bark Properties and Morphology
2.4. Statistical Analyses
3. Results
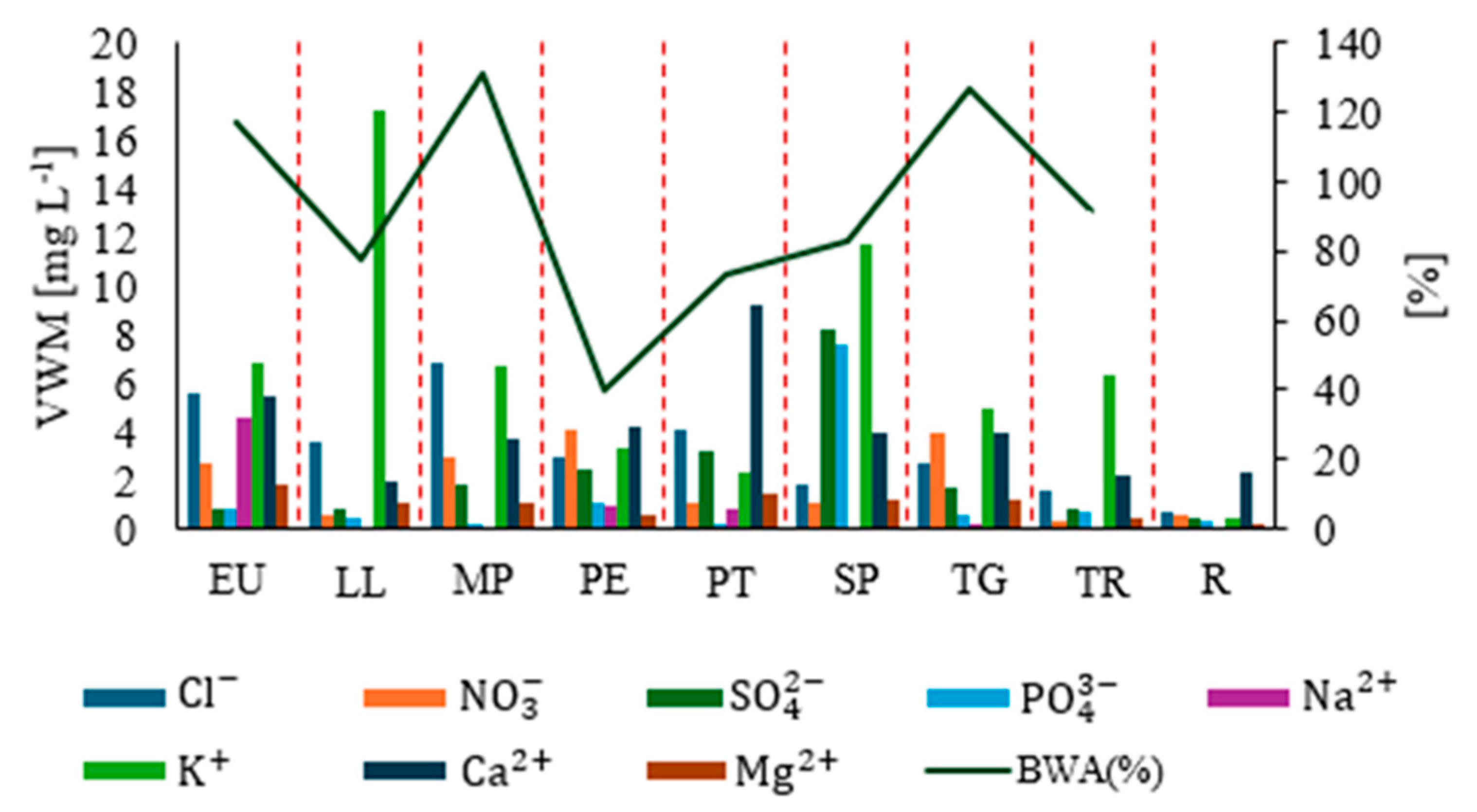
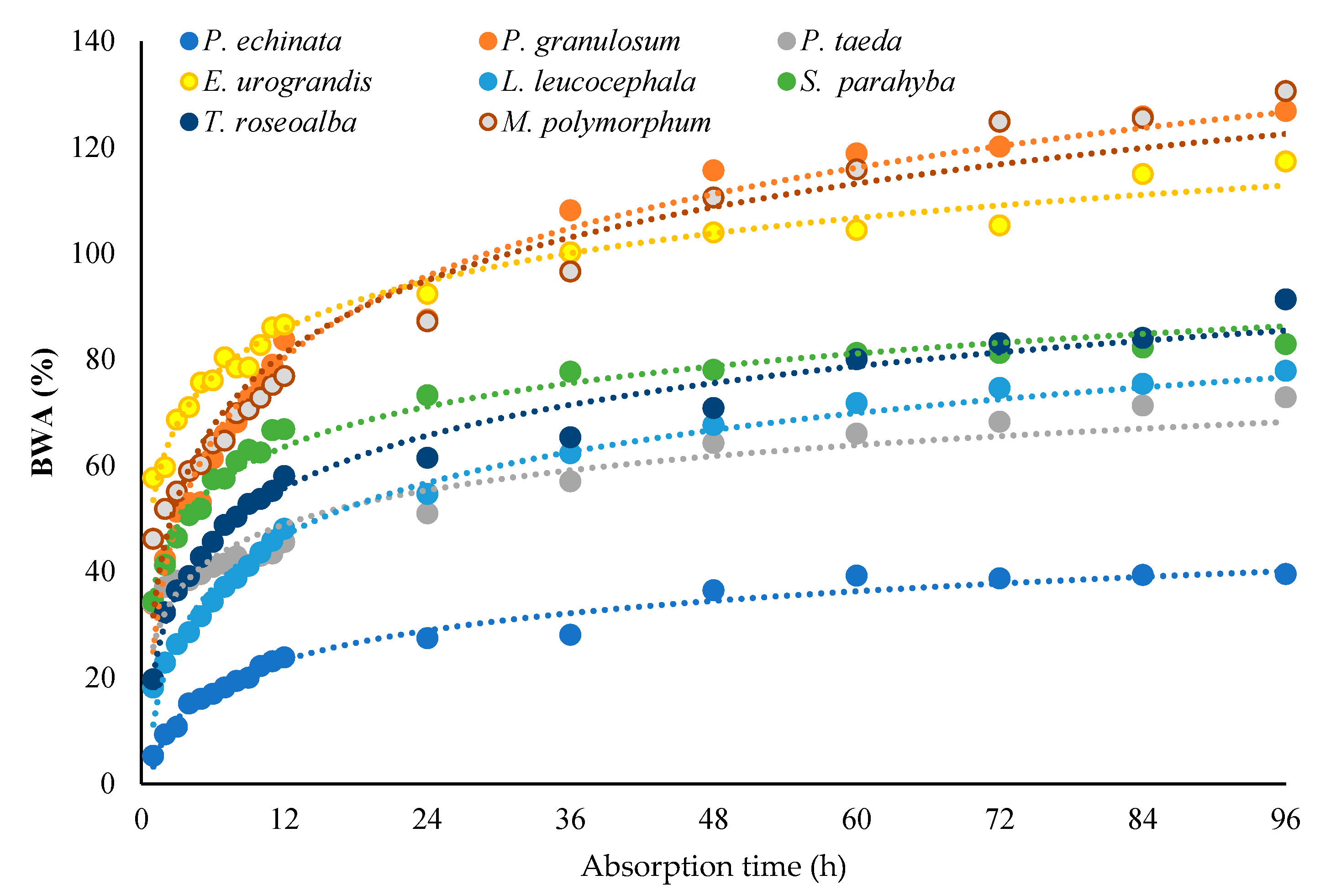
3.1. Nutrients Input and Bark Absorbability
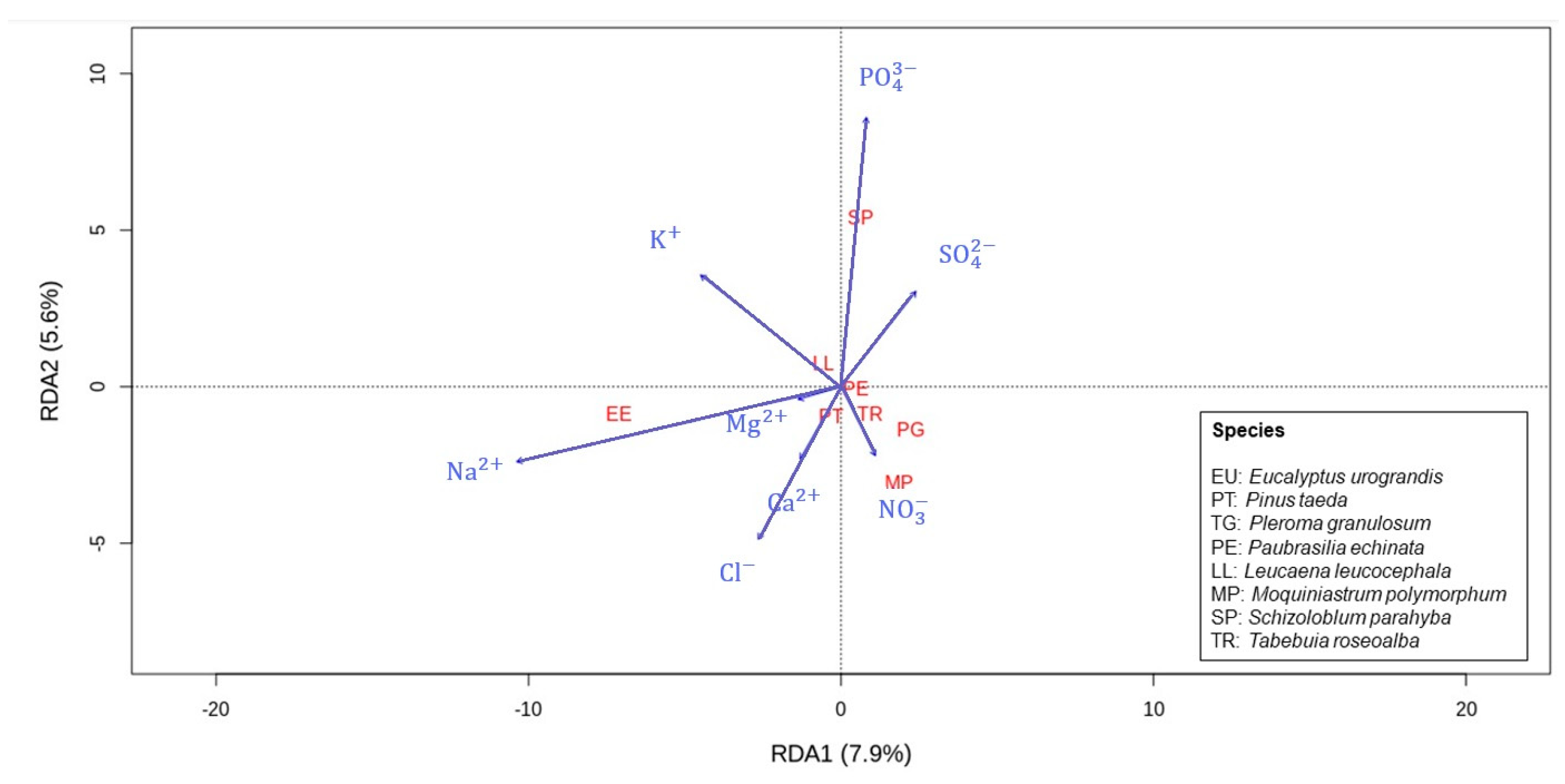
3.2. Ion Concentrations, Morphology, and Bark Absorption Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Momolli, D.R.; Schumacher, M.V.; Viera, M.; Ludvichak, A.A.; Guimarães, C.D.C.; Souza, H.P.D. Incident Precipitation Partitioning: The Canopy Interactions Enrich Water Solution with Nutrients in Throughfall and Stemflow. J. Agric. Sci. 2019, 11, 351. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Z.; Cao, Y.; Liu, L.; Hu, F.; Lu, X. Canopy Modification of Base Cations Deposition in a Subtropical Broadleaved Forest: Spatial Characteristics, Canopy Budgets and Acid Neutralizing Capacity. Ecol. Manag. 2021, 482, 118863. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Dymond, S.F.; Klamerus-Iwan, A. Bark-Water Interactions Across Ecosystem States and Fluxes. Front. For. Glob. Chang. 2021, 4, 660662. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Allen, S.T. What We Know About Stemflow’s Infiltration Area. Front. For. Glob. Chang. 2020, 3, 61. [Google Scholar] [CrossRef]
- Carlyle-Moses, D.E.; Iida, S.; Germer, S.; Llorens, P.; Michalzik, B.; Nanko, K.; Tischer, A.; Levia, D.F. Expressing Stemflow Commensurate with Its Ecohydrological Importance. Adv. Water Resour. 2018, 121, 472–479. [Google Scholar] [CrossRef]
- Astuti, H.P.; Suryatmojo, H. Water in the forest: Rain-vegetation interaction to estimate canopy interception in a tropical borneo rainforest. IOP Conf. Ser. Earth Environ. Sci. 2019, 361, 012035. [Google Scholar] [CrossRef]
- Schooling, J.T.; Levia, D.F.; Carlyle-Moses, D.E.; Dowtin, A.L.; Brewer, S.E.; Donkor, K.K.; Borden, S.A.; Grzybowski, A.A. Stemflow Chemistry in Relation to Tree Size: A Preliminary Investigation of Eleven Urban Park Trees in British Columbia, Canada. Urban. Urban. Green. 2017, 21, 129–133. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Wang, X.P.; Pan, Y.X.; Hu, R. Variations of Nutrients in Gross Rainfall, Stemflow, and Throughfall Within Revegetated Desert Ecosystems. Water Air Soil. Pollut. 2016, 227, 183. [Google Scholar] [CrossRef]
- Tonello, K.C.; Rosa, A.G.; Pereira, L.C.; Matus, G.N.; Guandique, M.E.G.; Navarrete, A.A. Rainfall Partitioning in the Cerrado and Its Influence on Net Rainfall Nutrient Fluxes. Agric. Meteorol. 2021, 303, 108372. [Google Scholar] [CrossRef]
- Aubrey, D.P.; Van Stan, J.T.; Gutmann, E.; Friesen, J. Relevance of Precipitation Partitioning to the Tree Water and Nutrient Balance Precipitation Partitioning by Vegetation: A Global Synthesis; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Germer, S.; Zimmermann, A.; Neill, C.; Krusche, A.V.; Elsenbeer, H. Disproportionate Single-Species Contribution to Canopy-Soil Nutrient Flux in an Amazonian Rainforest. Ecol. Manag. 2012, 267, 40–49. [Google Scholar] [CrossRef]
- Levia, D.F.; Frost, E.E. A Review and Evaluation of Stemflow Literature in the Hydrologic and Biogeochemical Cycles of Forested and Agricultural Ecosystems. J. Hydrol. 2003, 274, 1–29. [Google Scholar] [CrossRef]
- Levia, D.F.; Herwitz, S.R. Interspecific Variation of Bark Water Storage Capacity of Three Deciduous Tree Species in Relation to Stemflow Yield and Solute Flux to Forest Soils. Catena 2005, 64, 117–137. [Google Scholar] [CrossRef]
- Chuyong, G.B.; Newbery, D.M.; Songwe, N.C. Rainfall Input, Throughfall and Stemflow of Nutrients in a Central African Rain Forest Dominated by Ectomycorrhizal Trees. Biogeochemistry 2004, 67, 73–91. [Google Scholar] [CrossRef]
- Oka, A.; Takahashi, J.; Endoh, Y.; Seino, T. Bark Effects on Stemflow Chemistry in a Japanese Temperate Forest I. The Role of Bark Surface Morphology. Front. For. Glob. Chang. 2021, 4, 654375. [Google Scholar] [CrossRef]
- Song, Q.N.; Lu, H.; Liu, J.; Yang, J.; Yang, G.Y.; Yang, Q.P. Accessing the Impacts of Bamboo Expansion on NPP and N Cycling in Evergreen Broadleaved Forest in Subtropical China. Sci. Rep. 2017, 7, 40383. [Google Scholar] [CrossRef] [PubMed]
- Van Stan, J.T.; Levia, D.F. Inter- and Intraspecific Variation of Stemflow Production from Fagus Grandifolia Ehrh. (American Beech) and Liriodendron Tulipifera L. (Yellow Poplar) in Relation to Bark Microrelief in the Eastern United States. Ecohydrology 2010, 3, 11–19. [Google Scholar] [CrossRef]
- Tonello, K.C.; Campos, S.D.; Menezes, A.J.; Bramorski, J.; Mathias, S.L.; Lima, M.T. How Is Bark Absorbability and Wettability Related to Stemflow Yield? Observations From Isolated Trees in the Brazilian Cerrado. Front. For. Glob. Chang. 2021, 4, 650665. [Google Scholar] [CrossRef]
- Tobon Marin, C.; Bouten, I.W.; Dekker, S. Forest Foor Water Dynamics and Root Water Uptake in Four Forest Ecosystems in Northwest Amazonia. J. Hydrol. 2000, 237, 169–183. [Google Scholar] [CrossRef]
- Shearman, T.M.; Varner, J.M. Variation in Bark Allocation and Rugosity Across Seven Co-Occurring Southeastern US Tree Species. Front. For. Glob. Chang. 2021, 4, 731020. [Google Scholar] [CrossRef]
- Crockford, R.H.; Richardson, D.P. Partitioning of Rainfall into Throughfall, Stemfow and Interception: Effect of Forest Type, Ground Cover. and Climate. Hydrol. Process. 2000, 14, 2903–2920. [Google Scholar] [CrossRef]
- Siegert, C.; Ilek, A.; Wade, A.; Schweitzer, C. Changes in Bark Properties and Hydrology Following Prescribed Fire in Pinus Taeda and Quercus Montana. Hydrol. Process 2023, 37, e14799. [Google Scholar] [CrossRef]
- Dubreuil, V.; Fante, K.P.; Planchon, O.; Sant’Anna Neto, J.L. Climate Change Evidence in Brazil from Köppen’s Climate Annual Types Frequency. Int. J. Climatol. 2019, 39, 1446–1456. [Google Scholar] [CrossRef]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araujo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos; Embrapa Solos: Brasilia, Brazil, 2018. [Google Scholar]
- André, F.; Jonard, M.; Ponette, Q. Effects of Biological and Meteorological Factors on Stemflow Chemistry within a Temperate Mixed Oak-Beech Stand. Sci. Total Environ. 2008, 393, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Hofhansl, F.; Wanek, W.; Drage, S.; Huber, W.; Weissenhofer, A.; Richter, A. Controls of Hydrochemical Fluxes via Stemflow in Tropical Lowland Rainforests: Effects of Meteorology and Vegetation Characteristics. J. Hydrol. 2012, 452–453, 247–258. [Google Scholar] [CrossRef]
- Klamerus-Iwan, A.; Lasota, J.; Błońska, E. Interspecific Variability of Water Storage Capacity and Absorbability of Deadwood. Forests 2020, 11, 575. [Google Scholar] [CrossRef]
- Ilek, A.M.; Kucza, J.; Morkisz, K. Hydrological Properties of Bark of Selected Forest Tree Species. Part 2: Interspecific Variability of Bark Water Storage Capacity. Folia For. Pol. Ser. A 2017, 59, 110–122. [Google Scholar] [CrossRef][Green Version]
- Leps, J. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Oka, A.; Takahashi, J.; Endoh, Y.; Seino, T. Bark Effects on Stemflow Chemistry in a Japanese Temperate Forest II. The Role of Bark Anatomical Features. Front. For. Glob. Chang. 2021, 4, 657850. [Google Scholar] [CrossRef]
- Legrand, I.; Asta, J.; Goudard, Y. Variations in Bark Acidity and Conductivity over the Trunk Length of Silver Fir and Norway Spruce. Trees 1996, 11, 54–58. [Google Scholar] [CrossRef]
- Silva, M.A.; dos Santos, M.V.F.; Lira, M.A.; Dubeux Júnior, J.C.B.; de Andrade Silva, D.K.; Santoro, K.R.; de Arruda Leite, P.M.B.; de Freitas, E.V. Qualitative and Anatomical Characteristics of Tree-Shrub Legumes in the Forest Zone in Pernambuco State, Brazil. Rev. Bras. Zootec. 2012, 41, 2396–2404. [Google Scholar] [CrossRef][Green Version]
- Rodrigo, A.; Ávila, A.; Rodà, F. The Chemistry of Precipitation, Throughfall and Stemflow in two Holm Oak (Quercus ilex L.) Forests under a Contrasted Environment in NE Spain. Sci. Total Environ. 2003, 305, 195–205. [Google Scholar] [CrossRef]
- Tóbon, C.; Sevink, J.; Verstraten, J.M. Solute Fluxes in Throughfall and Stemflow in Four Forest Ecosystems in Northwest. Biogeochemistry 2004, 70, 1–25. [Google Scholar] [CrossRef]
- García Hortal, J.A. Fibras Papeleras; Edicions PC: Barcelona, Spain, 2007; 243p. [Google Scholar]
- Su, L.; Zhao, C.; Xu, W.; Xie, Z. Hydrochemical Fluxes in Bulk Precipitation, Throughfall, and Stemflow in a Mixed Evergreen and Deciduous Broadleaved Forest. Forests 2019, 10, 507. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | ID | DBH [cm] |
|---|---|---|
| Eucalyptus urograndis W. Hill | EU | 23.6 (2.9) |
| Leucaena leucocephala (Lam.) de Wit | LL | 19.4 (3.2) |
| Moquiniastrum polymorphum (Less.) G. Sancho | MP | 20.2 (1.4) |
| Paubrasilia echinata (Lam.) Gagnon, H. C. Lima & GPLewis | PE | 18.1 (1.3) |
| Pinus taeda L. | PT | 27.2 (3.7) |
| Schizoloblum parahyba (Vell.) Blake | SP | 21.1 (0.8) |
| Pleroma granulosum (Desr.) Cogn | TG | 21.9 (2.5) |
| Tabebuia roseoalba (Ridl.) Sandwith | TR | 20.0 (2.9) |
| Classes | DF | NF |
|---|---|---|
| 1 | ||
| 2 | ||
| 3 |
| Species | BWAmax(%) 1 | BWArate |
|---|---|---|
| E. urograndis | 117.35 | 0.62 |
| L. leucocephala | 77.85 | 0.62 |
| M. polymorphum | 130.62 | 0.88 |
| P. echinata | 39.52 | 0.36 |
| P. taeda | 72.88 | 0.40 |
| P. granulosum | 126.86 | 0.97 |
| T. roseoalba | 91.32 | 0.75 |
| S. parahyba | 82.92 | 0.51 |
| Class | Species 1 | Anions | Cations | BWArate | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bark texture | ||||||||||
| Furrowed | MP | 6.84 a | 2.93 a | 1.88 | 0.19 a | 0.04 a | 6.67 ab | 3.64 a | 1.10 a | 0.88 a |
| Scaly/ Exfoliating | EU | 3.35 a (0.69) | 2.41 a (0.75) | 1.82 a (0.47) | 0.68 a (0.13) | 1.30 a (0.83) | 4.75 b (0.87) | 5.01 a (0.16) | 1.11 a (0.26) | 0.62 a |
| PE | ||||||||||
| PT | ||||||||||
| PG | ||||||||||
| TR | ||||||||||
| Smooth | LL | 2.70 a (0.86) | 0.85 a (0.24) | 4.53 a (3.72) | 3.98 a (3.58) | 0.07 a (0.03) | 14.50 a (2.77) | 2.95 a (1.03) | 1.10 a (0.04) | 0.57 a |
| SP | ||||||||||
| Depth of Furrows in the bark | ||||||||||
| <0.02 | LL | 2.70 a (0.86) | 0.85 a (0.24) | 4.53 a (3.72) | 3.98 a (3.58) | 0.07 a (0.03) | 14.50 a (2.77) | 2.95 a (1.03) | 1.10 a (0.04) | 0.57 a |
| SP | ||||||||||
| ≥0.02 <0.10 | EU | 4.26 a (1.35) | 3.35 a (0.67) | 1.65 a (0.82) | 0.92 a (0.12) | 2.75 a (1.80) | 5.06 b (1.76) | 4.89 a (0.62) | 1.21 a (0.63) | 0.49 a |
| PE | ||||||||||
| ≥0.10 | MP | 3.76 a (1.02) | 2.07 a (0.75) | 1.92 a (0.44) | 0.44 a (0.11) | 0.26 a (0.16 | 5.07 b (0.89) | 4.73 a (1.36) | 1.04 a (0.19) | 0.75 a |
| PT | ||||||||||
| PG | ||||||||||
| TR | ||||||||||
| Number of furrows in the bark | ||||||||||
| <10 | LL | 2.70 a (0.86) | 0.85 a (0.24) | 4.53 a (3.72) | 3.98 a (3.58) | 0.07 a (0.03) | 14.50 a (2.77) | 2.95 a (1.03) | 1.11 a (0.04) | 0.57 ab |
| SP | ||||||||||
| ≥10.12 <50.28 | PE | 3.47 a (0.55) | 2.53 a (1.49) | 2.87 a (0.37) | 0.65 a (0.39) | 0.86 a (0.08) | 2.80 b (0.50) | 6.71 a (2.44) | 0.99 a (0.42) | 0.38 b |
| PT | ||||||||||
| ≥50.28 | EU | 4.16 a (0.11) | 2.48 a (0.69) | 1.31 a (0.26) | 0.58 a (0.12) | 1.20 a (1.00) | 6.20 a (0.39) | 3.82 a (0.61) | 1.15 a (0.26) | 0.81 a |
| MP | ||||||||||
| PG | ||||||||||
| TR | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, M.T.; Guandique, M.E.G.; Tonello, K.C. Bark Morphology and Nutrient Flux in Urban Trees: Investigating Water Absorption and Ion Concentration Dynamics. Hydrology 2024, 11, 56. https://doi.org/10.3390/hydrology11040056
Lima MT, Guandique MEG, Tonello KC. Bark Morphology and Nutrient Flux in Urban Trees: Investigating Water Absorption and Ion Concentration Dynamics. Hydrology. 2024; 11(4):56. https://doi.org/10.3390/hydrology11040056
Chicago/Turabian StyleLima, Marcelle Teodoro, Manuel Enrique Gamero Guandique, and Kelly Cristina Tonello. 2024. "Bark Morphology and Nutrient Flux in Urban Trees: Investigating Water Absorption and Ion Concentration Dynamics" Hydrology 11, no. 4: 56. https://doi.org/10.3390/hydrology11040056
APA StyleLima, M. T., Guandique, M. E. G., & Tonello, K. C. (2024). Bark Morphology and Nutrient Flux in Urban Trees: Investigating Water Absorption and Ion Concentration Dynamics. Hydrology, 11(4), 56. https://doi.org/10.3390/hydrology11040056