
Land-Based Impact of Nutrient Loads and Eutrophication on an Ancient Mediterranean Natural Lake

 , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling Strategy and Analyses
2.3. Nutrient Load Calculations
2.3.1. Point Sources
2.3.2. Non-Point Sources

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Group 1 | Description | References | Units | Export Coefficients | |
|---|---|---|---|---|---|---|
| TP | TN | |||||
| Point | UW | Permanent residents | [42,43] | kg × inhabitant y−1 | 0.33 | 2.25 |
| Point | UW | Floating population | 3/12 (3 months per year) the coefficient of permanent residents | kg × inhabitant y−1 | 0.0828 | 0.5544 |
| Point | IW | [33] | kg × employee y−1 | 0.45000 | 2.24840 | |
| Point | LS | Sheep | [44] adjusted as in [33] | kg × unit y−1 | 0.01000 | 0.06125 |
| Point | LS | Cows | kg × unit y−1 | 0.09250 | 0.68750 | |
| Point | LS | Horses | kg × unit y−1 | 0.10875 | 0.77500 | |
| Point | LS | Pigs | kg × unit y−1 | 0.04750 | 0.14125 | |
| Point | LS | Poultry | kg × unit y−1 | 0.00730 | 0.02555 | |
| Non-Point | UA | All areas identified in the preceding subdivision 1 (artificial surfaces) of the 4th-level CORINE LC | [43,45] | Kg × ha y−1 | 0.20 | 7.00 |
| Non-Point | AA | All areas identified in category 2 (used agricultural surfaces), except category 244 (agroforestry areas) of the 4th level CORINE LC | kg × ha y−1 | 0.60 | 16.00 | |
| Non-Point | NsNA | All areas identified in category 3 (forests and semi-natural areas) except the category 3315 (beds of streams wider than 25 m) + category 244 (agroforestry areas) of the 4th-level CORINE LC | kg × ha y−1 | 0.10 | 2.00 | |
2.4. Predictive Modeling
3. Results
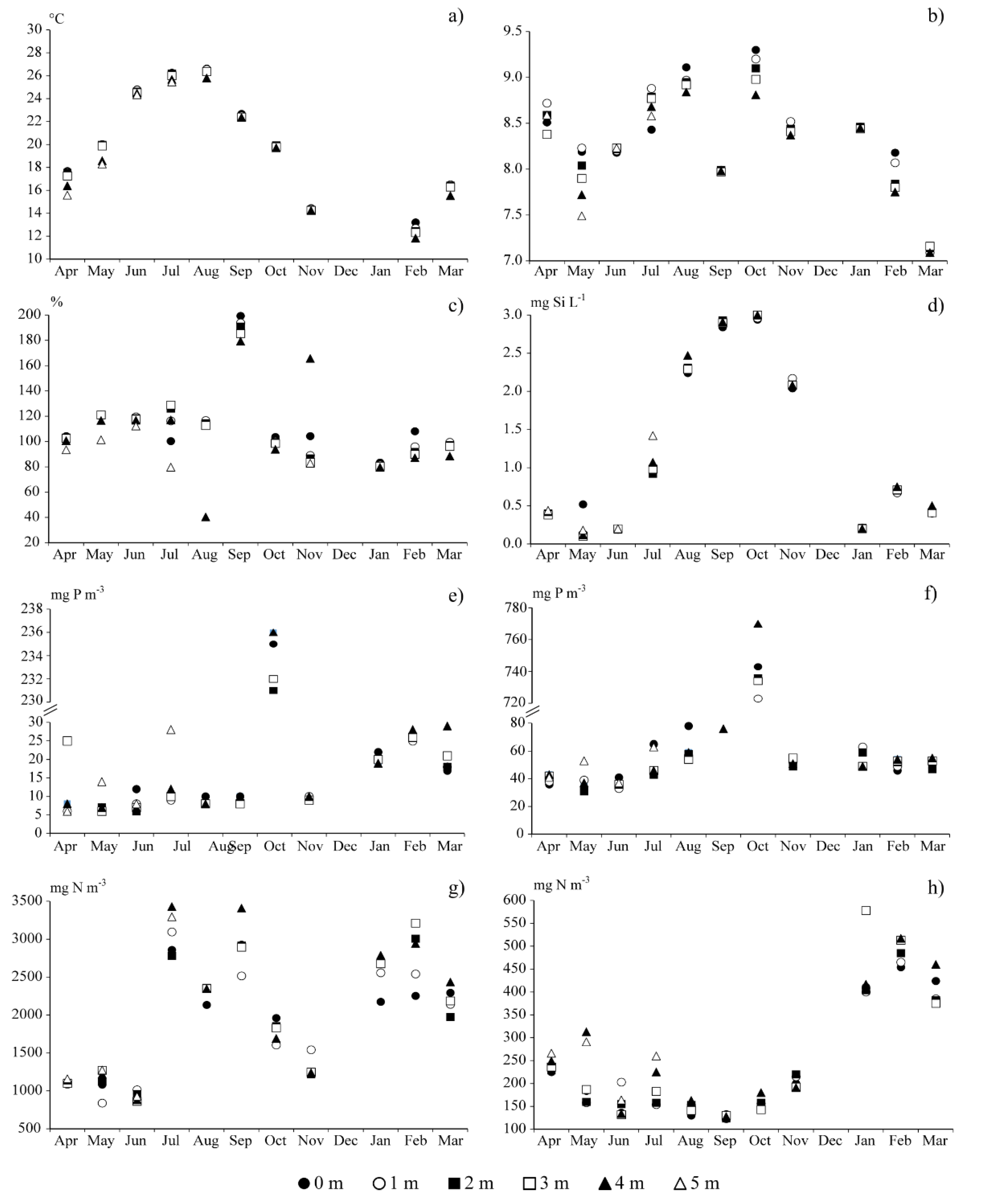
3.1. Environmental Conditions
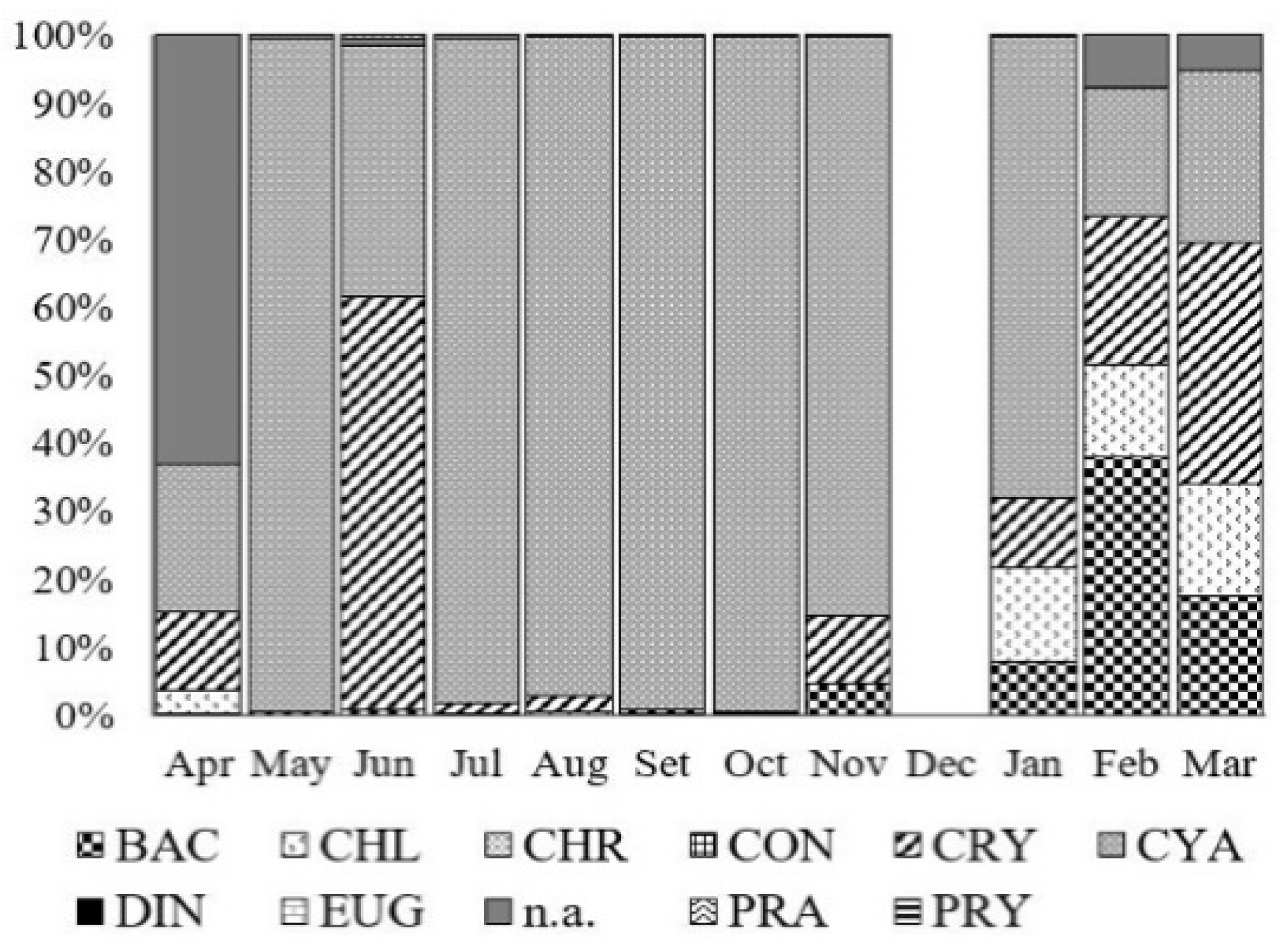
3.2. Chlorophyll and Phytoplankton
3.3. Trophic State
3.4. Nutrient Loads
3.5. Phosphorus Mass Balance and Predictive Modeling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Moal, M.; Gascuel-Odoux, C.; Ménesguen, A.; Souchon, Y.; Étrillard, C.; Levain, A.; Moatar, F.; Pannard, A.; Souchu, P.; Lefebvre, A.; et al. Eutrophication: A New Wine in an Old Bottle? Sci. Total Environ. 2019, 651, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of Excess Nutrient Inputs on Freshwater, Marine, and Terrestrial Ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef]
- Weigelhofer, G.; Hein, T.; Bondar-Kunze, E. Phosphorus and Nitrogen Dynamics in Riverine Systems: Human Impacts and Management Options. In Riverine Ecosystem Management. Aquatic Ecology Series; Schmutz, S., Sendzimir, J., Eds.; Springer: Cham, Switzerland, 2018; Volume 8, pp. 187–202. [Google Scholar] [CrossRef]
- Edwards, A.C.; Withers, P.J.A. Transport and Delivery of Suspended Solids, Nitrogen and Phosphorus from Various Sources to Freshwaters in the UK. J. Hydrol. 2008, 350, 144–153. [Google Scholar] [CrossRef]
- Bradford, A.; Denich, C. Rainwater Management to Mitigate the Effects of Development on the Urban Hydrologic Cycle. J. Green Build. 2007, 2, 37–52. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J.; Kozłowska, J.; Feniova, I.; Dzialowski, A.R. Zooplankton Community Responses to Oxygen Stress. Water 2020, 12, 706. [Google Scholar] [CrossRef] [Green Version]
- Fadda, A.; Marková, S.; Kotlík, P.; Lugliè, A.; Padedda, B.; Buscarinu, P.; Sechi, N.; Manca, M. First Record of Planktonic Crustaceans in Sardinian Reservoirs. Biologia 2011, 66, 856–865. [Google Scholar] [CrossRef]
- Mariani, M.A.; Padedda, B.M.; Kaštovský, J.; Buscarinu, P.; Sechi, N.; Virdis, T.; Lugliè, A. Effects of Trophic Status on Microcystin Production and the Dominance of Cyanobacteria in the Phytoplankton Assemblage of Mediterranean Reservoirs. Sci. Rep. 2015, 5, 17964. [Google Scholar] [CrossRef] [Green Version]
- Grizzetti, B.; Liquete, C.; Pistocchi, A.; Vigiak, O.; Zulian, G.; Bouraoui, F.; de Roo, A.; Cardoso, A.C. Relationship between Ecological Condition and Ecosystem Services in European Rivers, Lakes and Coastal Waters. Sci. Total Environ. 2019, 671, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.; Dong, J.; Yang, H.; van Zwieten, L.; Lu, H.; Alshameri, A.; Zhan, Z.; Chen, X.; Jiang, X.; et al. A Critical Review of Methods for Analyzing Freshwater Eutrophication. Water 2021, 13, 225. [Google Scholar] [CrossRef]
- Woelmer, W.M.; Kao, Y.-C.; Bunnell, D.B.; Deines, A.M.; Bennion, D.H.; Rogers, M.W.; Brooks, C.N.; Sayers, M.J.; Banach, D.M.; Grimm, A.G.; et al. Assessing the Influence of Watershed Characteristics on Chlorophyll a in Waterbodies at Global and Regional Scales. Inland Waters 2016, 6, 379–392. [Google Scholar] [CrossRef]
- Hasan, M.N.; Altaf, M.M.; Khan, N.A.; Khan, A.H.; Khan, A.A.; Ahmed, S.; Kumar, P.S.; Naushad, M.; Rajapaksha, A.U.; Iqbal, J.; et al. Recent Technologies for Nutrient Removal and Recovery from Wastewaters: A Review. Chemosphere 2021, 277, 130328. [Google Scholar] [CrossRef]
- Rout, P.R.; Shahid, M.K.; Dash, R.R.; Bhunia, P.; Liu, D.; Varjani, S.; Zhang, T.C.; Surampalli, R.Y. Nutrient Removal from Domestic Wastewater: A Comprehensive Review on Conventional and Advanced Technologies. J. Environ. Manag. 2021, 296, 113246. [Google Scholar] [CrossRef] [PubMed]
- Bol, R.; Gruau, G.; Mellander, P.-E.; Dupas, R.; Bechmann, M.; Skarbøvik, E.; Bieroza, M.; Djodjic, F.; Glendell, M.; Jordan, P.; et al. Challenges of Reducing Phosphorus Based Water Eutrophication in the Agricultural Landscapes of Northwest Europe. Front. Mar. Sci. 2018, 5, 276. [Google Scholar] [CrossRef] [Green Version]
- Meissner, R.; Seeger, J.; Rupp, H.; Balla, H. Assessing the Impact of Agricultural Land Use Changes on Water Quality. Water Sci. Technol. 1999, 40, 1–10. [Google Scholar] [CrossRef]
- Sharpley, A.N.; Gburek, W.J.; Folmar, G.; Pionke, H.B. Sources of Phosphorus Exported from an Agricultural Watershed in Pennsylvania. Agric. Water Manag. 1999, 41, 77–89. [Google Scholar] [CrossRef]
- Johnes, P.J. Evaluation and Management of the Impact of Land Use Change on the Nitrogen and Phosphorus Load Delivered to Surface Waters: The Export Coefficient Modelling Approach. J. Hydrol. 1996, 183, 323–349. [Google Scholar] [CrossRef]
- Caspers, H. OECD: Eutrophication of Waters. Monitoring, Assessment and Control.—154 Pp. Paris: Organisation for Economic Co-Operation and Development 1982. (Publié En Français Sous Le Titre »Eutrophication Des Eaux. Méthodes de Surveillance, d’Evaluation et de Lutte). Int. Rev. Gesamten Hydrobiol. Hydrogr. 1984, 69, 200. [Google Scholar] [CrossRef]
- Padedda, B.M.; Lugliè, A.; Ceccherelli, G.; Trebini, F.; Sechi, N. Nutrient-Flux Evaluation by the LOICZ Biogeochemical Model in Mediterranean Lagoons: The Case of Cabras Lagoon (Central-Western Sardinia). Chem. Ecol. 2010, 26, 147–162. [Google Scholar] [CrossRef]
- Jeppesen, E.; Moss, B.; Bennion, H.; Carvalho, L.; DeMeester, L.; Feuchtmayr, H.; Friberg, N.; Gessner, M.O.; Hefting, M.; Lauridsen, T.L.; et al. Interaction of Climate Change and Eutrophication. In Climate Change Impacts on Freshwater Ecosystems, 1st ed.; Kernan, M., Battarbee, R.W., Moss, B., Eds.; Blackwell Publishing Ltd.: Chichester, UK, 2010; pp. 119–151. [Google Scholar] [CrossRef] [Green Version]
- Dokulil, M.T.; Teubner, K. Eutrophication and Climate Change: Present Situation and Future Scenarios. In Eutrophication: Causes, Consequences and Control; Springer: Dordrecht, The Netherlands, 2010; pp. 1–16. [Google Scholar] [CrossRef]
- Scheffer, M. Ecology of Shallow Lakes; Springer: Dordrecht, The Netherlands, 2004. [Google Scholar] [CrossRef]
- Sechi, N.; Cossu, A. Note Preliminari Sull’ecologia Del Lago Baratz. In Proceedings of the Atti del 3° Congresso A.I.O.L., Sorrento, Italy, 1–3 December 1980. [Google Scholar]
- Lugliè, A. Indagine Preliminare Sulle Diatomee Sedimentarie Come Indicatrici Della Dinamica Ambientale Recente Del Lago Baratz (Sardegna Settentrionale). In Proceedings of the Atti del 7° Congresso A.I.O.L., Trieste, Italy, 11–14 June 1986; pp. 367–372. [Google Scholar]
- Sechi, N.; Lugliè, A. Phytoplankton in Sardinian Reservoirs. G. Bot. Ital. 1996, 130, 977–994. [Google Scholar] [CrossRef]
- Sollazzo, C.; Scanu, G.; Aste, F.; Belli, M.; Balzamo, S.; Martone, C.; Cadoni, F.; Bernabei, S.; Tartari, G.; Buraschi, E.; et al. Protocollo per Il Campionamento Dei Parametri Chimico-Fisici a Sostegno Degli Elementi Biologici in Ambiente Lacustre; APAT: Rome, Italy, 2008. [Google Scholar]
- American Water Works Association. Standard Methods for the Examination of Water and Wastewater; AWWA: Denver, CO, USA, 2017; pp. 185–192. [Google Scholar]
- Golterman, H.L.; Clymo, R.S.; Ohnstadt, M.A.M. Methods for Physical and Chemical Analysis of Freshwaters, 2nd ed.; Blackwell Scientific Publications: London, UK, 1978. [Google Scholar]
- Utermhöl, H. Zur Vervollkhung Der Quantitativen Phytoplanktonmethodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- Mariani, M.A.; Lai, G.G.; Padedda, B.M.; Pulina, S.; Sechi, N.; Virdis, T.; Lugliè, A. Long-Term Ecological Studies on Phytoplankton in Mediterranean Reservoirs: A Case Study from Sardinia (Italy). Inland Waters 2015, 5, 339–354. [Google Scholar] [CrossRef]
- Lai, G.G.; Padedda, B.M.; Wetzel, C.E.; Lugliè, A.; Sechi, N.; Ector, L. Epilithic Diatom Assemblages and Environmental Quality of the Su Gologone Karst Spring (Centraleastern Sardinia, Italy). Acta Bot. Croat. 2016, 75, 129–143. [Google Scholar] [CrossRef] [Green Version]
- Pulina, S.; Satta, C.T.; Padedda, B.M.; Sechi, N.; Lugliè, A. Seasonal Variations of Phytoplankton Size Structure in Relation to Environmental Variables in Three Mediterranean Shallow Coastal Lagoons. Estuar. Coast. Shelf Sci. 2018, 212, 95–104. [Google Scholar] [CrossRef]
- Sechi, N.; Botti, P.; Vacca, S.; Mosello, R. Deflusso e Rilascio Di Nutrienti Nel Bacino Del Rio Uvini (Sardegna Centro-Meridionale). In Proceedings of the Atti del III° Seminario Della Seconda Sezione Tecnica AIGR Insediamenti Zootecnici e Protezione dell’ambiente, Udine, Italy, 25–27 September 1991; pp. 609–616. [Google Scholar]
- Mosello, R.; Sechi, N.; Cossu, A. Il Carico Chimico Veicolato al Lago Oschiri. In Proceedings of the Qualità dell’acqua in Sardegna, Cagliari, Italy, 1989. [Google Scholar]
- Sechi, N.; Mosello, R. Nutrient Budget and Trophic Level of Lake Oschiri (Sardinia. Italy). Mem. Dell’istituto Ital. Di Idrobiol. 1985, 43, 55–75. [Google Scholar]
- Padedda, B.M.; Sechi, N.; Lai, G.G.; Mariani, M.A.; Pulina, S.; Satta, C.T.; Bazzoni, A.M.; Virdis, T.; Buscarinu, P.; Lugliè, A. A Fast-Response Methodological Approach to Assessing and Managing Nutrient Loads in Eutrophic Mediterranean Reservoirs. Ecol. Eng. 2015, 85, 47–55. [Google Scholar] [CrossRef]
- Istituto Nazionale di Statistica. Censimento Permanente Della Popolazione e Delle Abitazioni. Available online: http://dati-censimentopopolazione.istat.it/Index.aspx (accessed on 10 January 2020).
- Istituto Nazionale di Statistica. 9° Censimento Dell’industria e Dei Servizi e Censimento Delle Istituzioni Non Profit. Available online: http://dati.istat.it/Index.aspx?DataSetCode=DICA_ASIAULP (accessed on 10 January 2020).
- Banca Dati Nazionale Dell’Anagrafe Zootecnica. Available online: https://www.vetinfo.it/j6_statistiche/#/ (accessed on 1 May 2019).
- Sardegna Geoportale. Carta Dell’Uso Del Suolo in Scala 1:25,000—Edizione 2008 (Strati Poligonali). Regione Autonoma della Sardegna, Cagliari. 2008. Available online: http://dati.regione.sardegna.it/en/dataset/carta-delluso-del-suolo-in-scala-1-25-000-elementi-poligonali-2008 (accessed on 10 January 2020).
- CORINE Land Cover Technical Guide—Addendum 2000—European Environment Agency. Available online: https://www.eea.europa.eu/publications/tech40add (accessed on 8 October 2021).
- Tartari, G.; Marchetto, A.; Buzzi, F.; Pagnotta, R.; Barbiero, G. Notiziario dei Metodi Analitici; IRSA CNR: Roma, Italy, 2008. [Google Scholar]
- Marchetti, R.; Gerletti, M.; Clamari, D.; Chiaudani, G. Elementi e Criteri per La Definizione Del Livello Di Accettabilità Delle Acque Di Scarico. Quad. IRSA 1973, 24, 255. [Google Scholar]
- Chiaudani, G.; Gerletti, M.; Marchetti, R.; Provini, A.; Vighi, M. Il Problema Dell’Eutrofizzazione in Italia. Quad. IRSA 1978, 42, 93. [Google Scholar]
- Marchetti, R.; Verna, N. Quantification of the Phosphorus and Nitrogen Loads in the Minor Rivers of the Emilia-Romagna Coast (Italy). A Methodological Study on the Use of Theoretical Coefficients in Calculating the Loads. In Marine Coastal Eutrophication, Proceedings of the International Conference, Bologna, Italy, 21–24 March 1990; Elsevier: Amsterdam, The Netherlands; pp. 315–336. [CrossRef]
- Marchetti, R.; Barone, R.; Calvo, S.; Lugliè, A.; Naselli-Flores, L.; Sechi, N. Studies on Italian Reservoirs. Mem. Dell’istituto Ital. Di Idrobiol. 1992, 50, 337–363. [Google Scholar]
- Sechi, N. Le Problematiche Legate al Fitoplancton Nelle Acque Lacustri Destinate Ad Uso Potabile. G. Bot. Ital. 1992, 126, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Marchetto, A.; Padedda, B.M.; Mariani, M.A.; Lugliè, A.; Sechi, N. A Numerical Index for Evaluating Phytoplankton Response to Changes in Nutrient Levels in Deep Mediterranean Reservoirs. J. Limnol. 2009, 68, 106–121. [Google Scholar] [CrossRef] [Green Version]
- Padedda, B.M.; Sechi, N.; Lai, G.G.; Mariani, M.A.; Pulina, S.; Sarria, M.; Satta, C.T.; Virdis, T.; Buscarinu, P.; Lugliè, A. Consequences of Eutrophication in the Management of Water Resources in Mediterranean Reservoirs: A Case Study of Lake Cedrino (Sardinia, Italy). Glob. Ecol. Conserv. 2017, 12, 21–35. [Google Scholar] [CrossRef]
- Giadrossich, F.; Niedda, M.; Cohen, D.; Pirastru, M. Evaporation in a Mediterranean Environment by Energy Budget and Penman Methods, Lake Baratz, Sardinia, Italy. Hydrol. Earth Syst. Sci. 2015, 19, 2451–2468. [Google Scholar] [CrossRef] [Green Version]
- Vollenweider, R.A.; Kerekes, J. Eutrophication of Waters. Monitoring, Assessment and Control; OECD Cooperative Programme on Monitoring of Inland Waters (Eutrophication Control): Paris, France, 1982. [Google Scholar]
- Carlson, R.E. A Trophic State Index for Lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Padedda, B.M.; Sechi, N. Condizioni Del Lago Cedrino e Relazioni Con Il Suo Bacino Versante. Nat. Sard. 2008, 30, 42–49. [Google Scholar]
- Messineo, V.; Bogialli, S.; Melchiorre, S.; Sechi, N.; Lugliè, A.; Casiddu, P.; Mariani, M.A.; Padedda, B.M.; di Corcia, A.; Mazza, R.; et al. Cyanobacterial Toxins in Italian Freshwaters. Limnol. Ecol. Manag. Inland Waters 2009, 39, 95–106. [Google Scholar] [CrossRef]
- Wetzel, R.G. Microcommunities and Microgradients: Linking Nutrient Regeneration, Microbial Mutualism, and High Sustained Aquatic Primary Production. Neth. J. Aquat. Ecol. 1993, 27, 3–9. [Google Scholar] [CrossRef]
- Leip, A.; Billen, G.; Garnier, J.; Grizzetti, B.; Lassaletta, L.; Reis, S.; Simpson, D.; Sutton, M.A.; de Vries, W.; Weiss, F.; et al. Impacts of European Livestock Production: Nitrogen, Sulphur, Phosphorus and Greenhouse Gas Emissions, Land-Use, Water Eutrophication and Biodiversity. Environ. Res. Lett. 2015, 10, 115004. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Bennett, E.M. The Influence of Agricultural Trade and Livestock Production on the Global Phosphorus Cycle. Ecosystems 2011, 15, 256–268. [Google Scholar] [CrossRef]
- Nature-Based Solutions Improving Water Quality & Waterbody Conditions—Publications Office of the EU. Available online: https://op.europa.eu/it/publication-detail/-/publication/d6efaeeb-d530-11ea-adf7-01aa75ed71a1/language-en (accessed on 8 October 2021).
- Riggio, V.A.; Ruffino, B.; Campo, G.; Comino, E.; Comoglio, C.; Zanetti, M. Constructed Wetlands for the Reuse of Industrial Wastewater: A Case-Study. J. Clean. Prod. 2018, 171, 723–732. [Google Scholar] [CrossRef]
- Stefanakis, A.I. The Role of ConstructedWetlands as Green Infrastructure for Sustainable Urban Water Management. Sustainability 2019, 11, 6981. [Google Scholar] [CrossRef] [Green Version]
- Nivala, J.; Murphy, C.; Freeman, A. Recent Advances in the Application, Design, and Operations & Maintenance of Aerated Treatment Wetlands. Water 2020, 12, 1188. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, A.; Bresciani, R.; Martinuzzi, N.; Masi, F. French Reed Bed as a Solution to Minimize the Operational and Maintenance Costs of Wastewater Treatment from a Small Settlement: An Italian Example. Water 2018, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Virsta, A. Floodplain Revegetation and River Basin Restoration. Environ. Eng. Manag. J. 2007, 6, 275–280. [Google Scholar] [CrossRef]
- Carreira, J.A.; Viñegla, B.; García-Ruiz, R.; Ochoa, V.; Hinojosa, M.B. Recovery of Biochemical Functionality in Polluted Flood-Plain Soils: The Role of Microhabitat Differentiation through Revegetation and Rehabilitation of the River Dynamics. Soil Biol. Biochem. 2008, 40, 2088–2097. [Google Scholar] [CrossRef]
- Haddaway, N.R.; Brown, C.; Eales, J.; Eggers, S.; Josefsson, J.; Kronvang, B.; Randall, N.P.; Uusi-Kämppä, J. The Multifunctional Roles of Vegetated Strips around and within Agricultural Fields. Environ. Evid. 2018, 7, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Prosser, R.S.; Hoekstra, P.F.; Gene, S.; Truman, C.; White, M.; Hanson, M.L. A Review of the Effectiveness of Vegetated Buffers to Mitigate Pesticide and Nutrient Transport into Surface Waters from Agricultural Areas. J. Environ. Manag. 2020, 261, 110210. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Barreira, A.P.; Loures, L.; Antunes, D.; Panagopoulos, T. Stakeholders’ Engagement on Nature-Based Solutions: A Systematic Literature Review. Sustainability 2020, 12, 640. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F.; Lionello, P. Climate Change Projections for the Mediterranean Region. Glob. Planet. Change 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Gao, X.; Giorgi, F. Increased Aridity in the Mediterranean Region under Greenhouse Gas Forcing Estimated from High Resolution Simulations with a Regional Climate Model. Glob. Planet. Change 2008, 62, 195–209. [Google Scholar] [CrossRef]
- Naselli-Flores, L. Man-Made Lakes in Mediterranean Semi-Arid Climate: The Strange Case of Dr Deep Lake and Mr Shallow Lake. Hydrobiologia 2003, 506–509, 13–21. [Google Scholar] [CrossRef]
- Padedda, B.M.; Pulina, S.; Magni, P.; Sechi, N.; Lugliè, A. Phytoplankton Dynamics in Relation to Environmental Changes in a Phytoplankton-Dominated Mediterranean Lagoon (Cabras Lagoon, Italy). Ital. Assoc. Oceanogr. Limnol. 2012, 3, 147–169. [Google Scholar] [CrossRef]




| OECD Model | Trophic Classes | Med TP | Med CHLa | Max CHLa | SD | Mean |
| 118 mg P m−3 | 6 µg L−1 | 21 µg L−1 | 3.4 m | |||
| hyper-eutrophy | 60% | 1% | 10% | 8% | 20% | |
| eutrophic | 37% | 24% | 35% | 44% | 35% | |
| mesotrophic | 3% | 61% | 54% | 42% | 40% | |
| oligotrophic | 0% | 14% | 1% | 6% | 5% | |
| ultra-oligotrophic | 0% | 0% | 0% | 0% | 0% | |
| TSI | 73 | 47 | - | 42 | 54 |
| Type | Group | P (t y−1) | N (t y−1) | P (%) | N (%) |
|---|---|---|---|---|---|
| Point | UW | 0.005 | 0.034 | 1.1 | 0.3 |
| LS | 0.033 | 0.185 | 7.2 | 1.7 | |
| IW | 0.000 | 0.000 | 0.0 | 0.0 | |
| subtotal | 0.038 | 0.219 | 8.4 | 2.0 | |
| Non-point | UA | 0.002 | 0.060 | 0.4 | 0,5 |
| AA | 0.356 | 9.527 | 78.4 | 86,7 | |
| NsNA | 0.058 | 1.186 | 12.8 | 10,8 | |
| subtotal | 0.417 | 10.773 | 91.6 | 98.0 | |
| Total | 0.455 | 10.992 | 100 | 100 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padedda, B.M.; Lugliè, A.; Lai, G.G.; Giadrossich, F.; Satta, C.T.; Pulina, S. Land-Based Impact of Nutrient Loads and Eutrophication on an Ancient Mediterranean Natural Lake. Hydrology 2022, 9, 7. https://doi.org/10.3390/hydrology9010007
Padedda BM, Lugliè A, Lai GG, Giadrossich F, Satta CT, Pulina S. Land-Based Impact of Nutrient Loads and Eutrophication on an Ancient Mediterranean Natural Lake. Hydrology. 2022; 9(1):7. https://doi.org/10.3390/hydrology9010007
Chicago/Turabian StylePadedda, Bachisio Mario, Antonella Lugliè, Giuseppina Grazia Lai, Filippo Giadrossich, Cecilia Teodora Satta, and Silvia Pulina. 2022. "Land-Based Impact of Nutrient Loads and Eutrophication on an Ancient Mediterranean Natural Lake" Hydrology 9, no. 1: 7. https://doi.org/10.3390/hydrology9010007
APA StylePadedda, B. M., Lugliè, A., Lai, G. G., Giadrossich, F., Satta, C. T., & Pulina, S. (2022). Land-Based Impact of Nutrient Loads and Eutrophication on an Ancient Mediterranean Natural Lake. Hydrology, 9(1), 7. https://doi.org/10.3390/hydrology9010007