

In Vitro Bioassay for Damage-Associated Molecular Patterns Arising from Injured Oral Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Lines
2.2. Cell Lysates
2.3. Reverse Transcription Quantitative Real-Time PCR (RT-qPCR)
2.4. Immunoassay
2.5. Statistical Analysis
3. Results
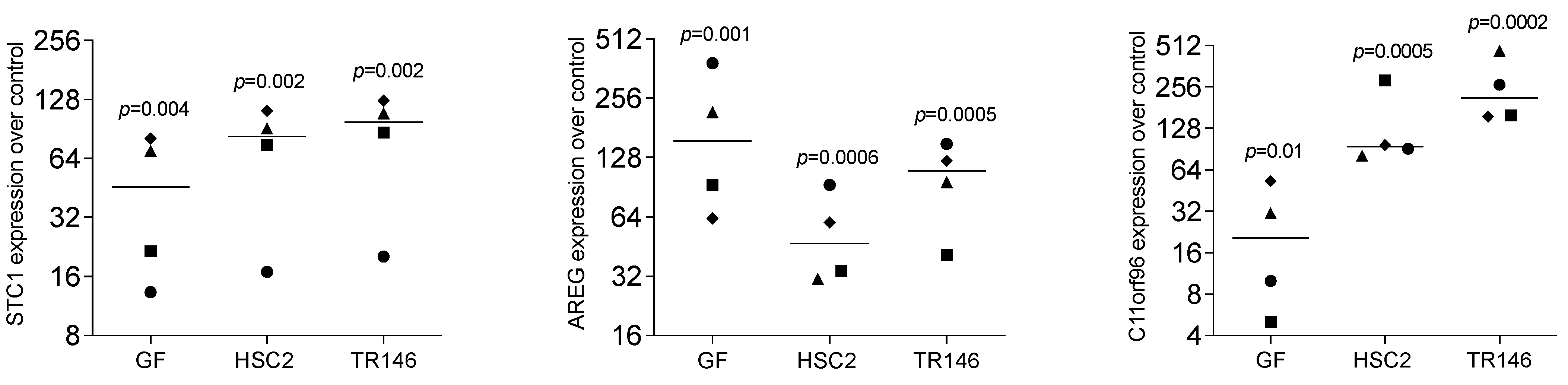
3.1. Necrotic Oral Cell Lysate Drives Gene Expression in Gingival Fibroblasts
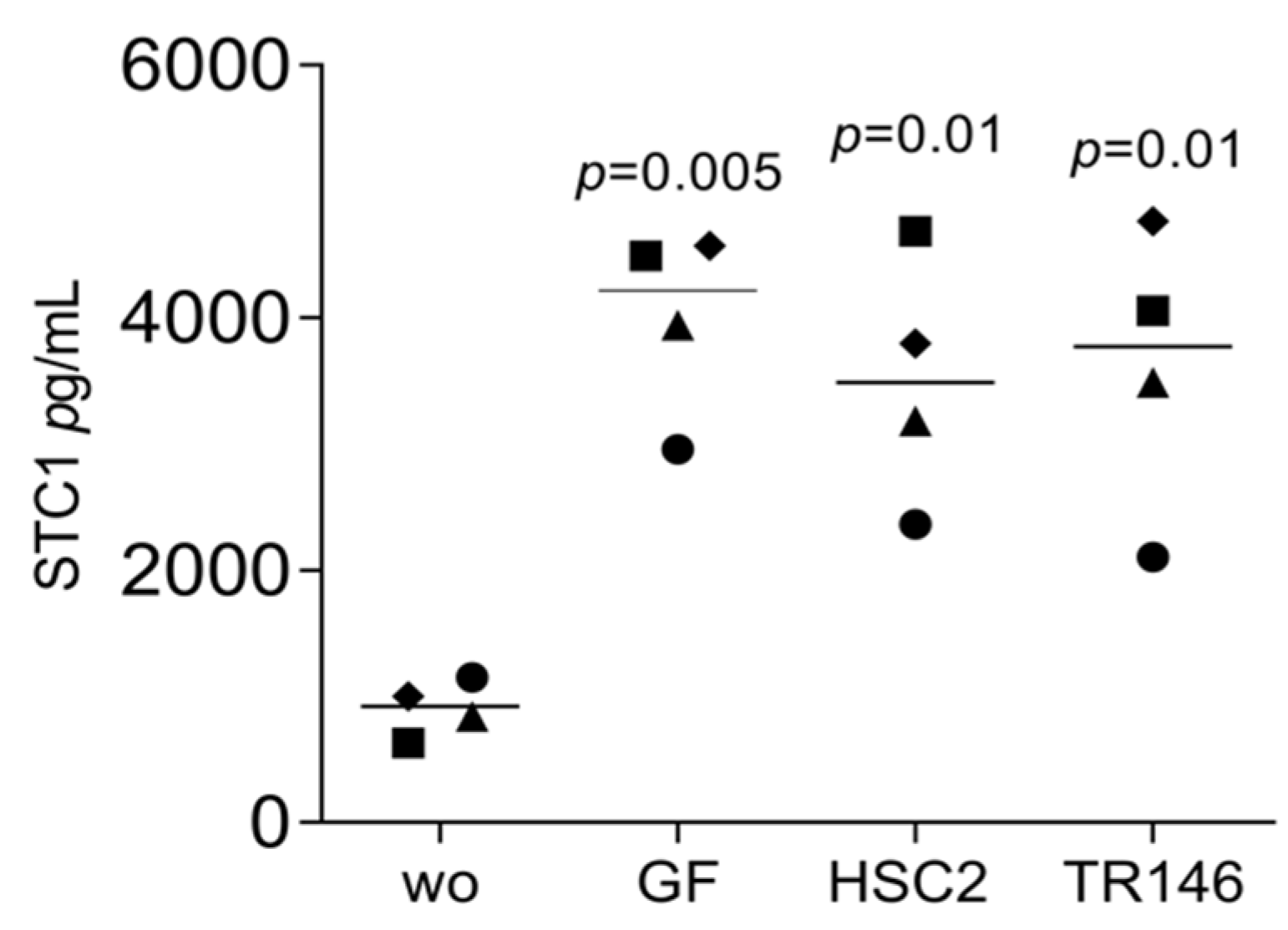
3.2. STC1, AREG, and C11orf96 Are Increased by Oral Cell Lysate Depending on SB431542
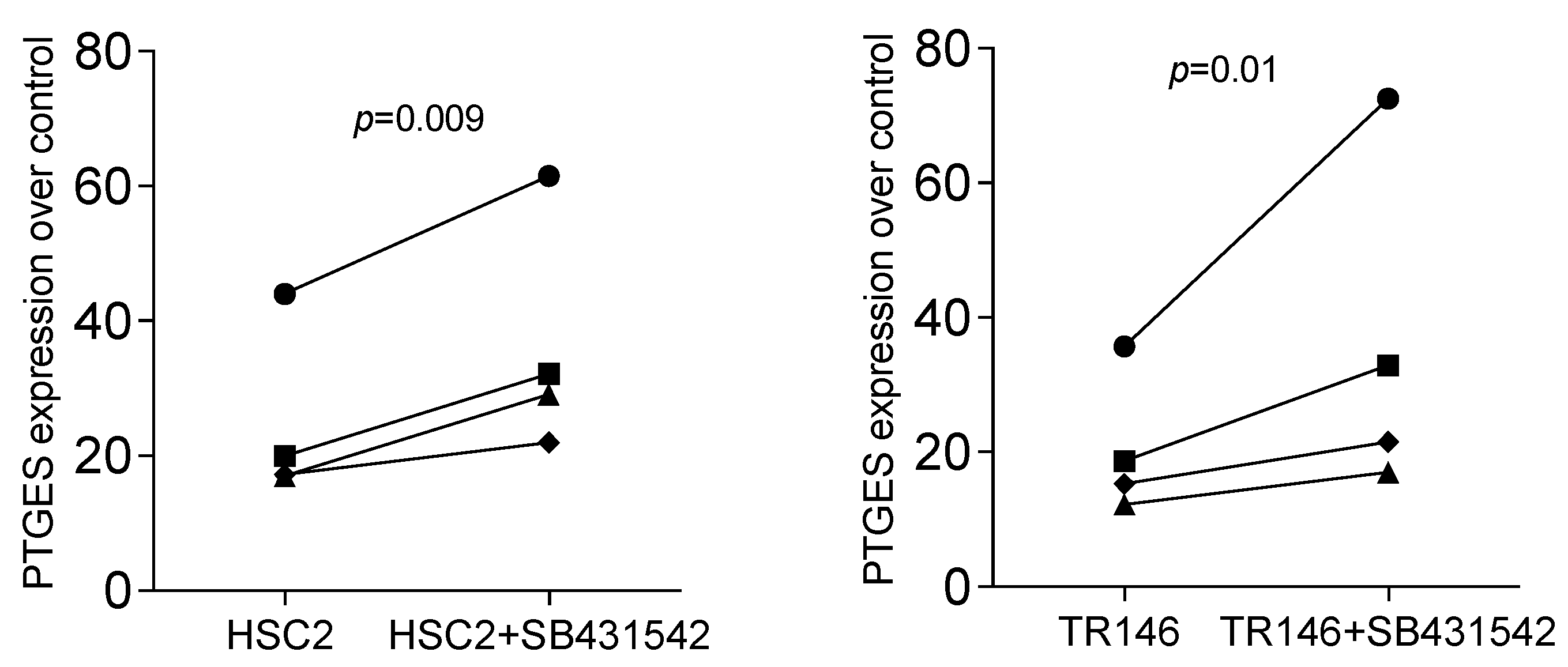
3.3. PTGES Is Regulated by Oral Cell Lysate but Enhanced by SB431542
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moutsopoulos, N.M.; Konkel, J.E. Tissue-Specific Immunity at the Oral Mucosal Barrier. Trends Immunol. 2018, 39, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Dutzan, N.; Abusleme, L.; Bridgeman, H.; Greenwell-Wild, T.; Zangerle-Murray, T.; Fife, M.E.; Bouladoux, N.; Linley, H.; Brenchley, L.; Wemyss, K.; et al. On-going Mechanical Damage from Mastication Drives Homeostatic Th17 Cell Responses at the Oral Barrier. Immunity 2017, 46, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Liu, B.; Cuevas, P.; Brunski, J.; Aellos, F.; Petersen, J.; Koehne, T.; Broer, S.; Gruber, R.; LeBlanc, A.; et al. Linking the Mechanics of Chewing to Biology of the Junctional Epithelium. J. Dent. Res. 2023, 102, 1252–1260. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jiang, W.; Zhou, R. DAMP sensing and sterile inflammation: Intracellular, intercellular and inter-organ pathways. Nat. Rev. Immunol. 2024, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Jiang, W.; Zhou, R. DAMPs and DAMP-sensing receptors in inflammation and diseases. Immunity 2024, 57, 752–771. [Google Scholar] [CrossRef] [PubMed]
- Deas, D.E.; Moritz, A.J.; Sagun, R.S., Jr.; Gruwell, S.F.; Powell, C.A. Scaling and root planing vs. conservative surgery in the treatment of chronic periodontitis. Periodontology 2000 2016, 71, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Coyac, B.R.; Arioka, M.; Leahy, B.; Tulu, U.S.; Aghvami, M.; Holst, S.; Hoffmann, W.; Quarry, A.; Bahat, O.; et al. A Novel Osteotomy Preparation Technique to Preserve Implant Site Viability and Enhance Osteogenesis. J. Clin. Med. 2019, 8, 170. [Google Scholar] [CrossRef]
- Aghvami, M.; Brunski, J.B.; Serdar Tulu, U.; Chen, C.H.; Helms, J.A. A Thermal and Biological Analysis of Bone Drilling. J. Biomech. Eng. 2018, 140, 1010101–1010108. [Google Scholar] [CrossRef]
- Kujan, O.; Azzeghaiby, S.N.; Tarakji, B.; Abuderman, A.; Sakka, S. Cryosurgery of the oral and peri-oral region: A literature review of the mechanism, tissue response, and clinical applications. J. Investig. Clin. Dent. 2013, 4, 71–77. [Google Scholar] [CrossRef]
- Demirbilek, N.; Evren, C. Is Piezoelectric Surgery Really Harmless to Soft Tissue? J. Craniofac. Surg. 2019, 30, 1966–1969. [Google Scholar] [CrossRef]
- Fathima, S.D.; Gururaj, N.; Sivapathasundharam, B.; Vennila, A.A.; Keerthik Lavanya, M.K.; Sarayushivani, U. Histopathological significance of necrosis in oral lesions: A review. J. Oral Maxillofac. Pathol. 2023, 27, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Tonnus, W.; Belavgeni, A.; Beuschlein, F.; Eisenhofer, G.; Fassnacht, M.; Kroiss, M.; Krone, N.P.; Reincke, M.; Bornstein, S.R.; Linkermann, A. The role of regulated necrosis in endocrine diseases. Nat. Rev. Endocrinol. 2021, 17, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.W.; Zhao, L.; Mei, Z.S.; Zhou, Y.H.; Yu, T. Association between periodontitis and uric acid levels in blood and oral fluids: A systematic review and meta-analysis. BMC Oral Health 2023, 23, 178. [Google Scholar] [CrossRef] [PubMed]
- Furuse, N.; Takai, H.; Ogata, Y. Effects of Initial Periodontal Therapy on Heat Shock Protein 70 Levels in Gingival Crevicular Fluid from Periodontitis Patients. J. Clin. Med. 2020, 9, 3072. [Google Scholar] [CrossRef] [PubMed]
- Eren, G.; Turkoglu, O.; Atmaca, H.; Atilla, G. Evaluation of gingival crevicular fluid cyclophilin a and extracellular matrix metalloproteinase inducer levels in different periodontal diseases. Arch. Oral Biol. 2016, 68, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Turer, C.C.; Balli, U.; Guven, B. Fetuin-A, serum amyloid A and tumor necrosis factor alpha levels in periodontal health and disease. Oral Dis. 2017, 23, 379–386. [Google Scholar] [CrossRef]
- Luo, L.; Xie, P.; Gong, P.; Tang, X.H.; Ding, Y.; Deng, L.X. Expression of HMGB1 and HMGN2 in gingival tissues, GCF and PICF of periodontitis patients and peri-implantitis. Arch. Oral Biol. 2011, 56, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Ebe, N.; Hara-Yokoyama, M.; Iwasaki, K.; Iseki, S.; Okuhara, S.; Podyma-Inoue, K.A.; Terasawa, K.; Watanabe, A.; Akizuki, T.; Watanabe, H.; et al. Pocket epithelium in the pathological setting for HMGB1 release. J. Dent. Res. 2011, 90, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, Y.; Kim, E.; Kwak, A.; Ryoo, S.; Bae, S.H.; Azam, T.; Kim, S.; Dinarello, C.A. The Interleukin-1alpha Precursor is Biologically Active and is Likely a Key Alarmin in the IL-1 Family of Cytokines. Front. Immunol. 2013, 4, 391. [Google Scholar] [CrossRef]
- Koli, K.; Saharinen, J.; Hyytiainen, M.; Penttinen, C.; Keski-Oja, J. Latency, activation, and binding proteins of TGF-beta. Microsc. Res. Tech. 2001, 52, 354–362. [Google Scholar] [CrossRef]
- Panahipour, L.; Omerbasic, A.; Nasirzade, J.; Gruber, R. TGF-beta Activity of a Demineralized Bone Matrix. Int. J. Mol. Sci. 2021, 22, 664. [Google Scholar] [CrossRef] [PubMed]
- Panahipour, L.; Abbasabadi, A.O.; Wagner, A.; Kratochwill, K.; Pichler, M.; Gruber, R. Bone Allograft Acid Lysates Change the Genetic Signature of Gingival Fibroblasts. Int. J. Mol. Sci. 2023, 24, 16181. [Google Scholar] [CrossRef] [PubMed]
- Panahipour, L.; Abbasabadi, A.O.; Gruber, R. Gingival Fibroblasts Are Sensitive to Oral Cell Lysates Indicated by Their IL11 Expression. Bioengineering 2023, 10, 1193. [Google Scholar] [CrossRef] [PubMed]
- Miranda, T.S.; Figueiredo, N.F.; Figueiredo, L.C.; Silva, H.; Rocha, F.R.G.; Duarte, P.M. Cytokine profiles of healthy and diseased sites in individuals with periodontitis. Arch. Oral Biol. 2020, 120, 104957. [Google Scholar] [CrossRef] [PubMed]
- Gruskin, E.; Doll, B.A.; Futrell, F.W.; Schmitz, J.P.; Hollinger, J.O. Demineralized bone matrix in bone repair: History and use. Adv. Drug Deliv. Rev. 2012, 64, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.H.; Kao, S.Y.; Chen, C.L.; Yuliani, F.S.; Lin, L.Y.; Lin, C.H.; Chen, B.C. Amphiregulin induces CCN2 and fibronectin expression by TGF-beta through EGFR-dependent pathway in lung epithelial cells. Respir. Res. 2022, 23, 381. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, M.; Lehner, D.; Handschuh, S.; Jay, F.F.; Erben, R.G.; Schneider, M.R. Osteoblast-specific overexpression of amphiregulin leads to transient increase in femoral cancellous bone mass in mice. Bone 2015, 81, 36–46. [Google Scholar] [CrossRef]
- Jay, F.F.; Vaidya, M.; Porada, S.M.; Andrukhova, O.; Schneider, M.R.; Erben, R.G. Amphiregulin lacks an essential role for the bone anabolic action of parathyroid hormone. Mol. Cell Endocrinol. 2015, 417, 158–165. [Google Scholar] [CrossRef]
- Shao, J.; Sheng, H. Amphiregulin promotes intestinal epithelial regeneration: Roles of intestinal subepithelial myofibroblasts. Endocrinology 2010, 151, 3728–3737. [Google Scholar] [CrossRef]
- Nevo, S.; Frenkel, N.; Kadouri, N.; Gome, T.; Rosenthal, N.; Givony, T.; Avin, A.; Peligero Cruz, C.; Kedmi, M.; Lindzen, M.; et al. Tuft cells and fibroblasts promote thymus regeneration through ILC2-mediated type 2 immune response. Sci. Immunol. 2024, 9, eabq6930. [Google Scholar] [CrossRef]
- Jia, X.; Wang, G.; Wu, L.; Pan, H.; Ling, L.; Zhang, J.; Wen, Q.; Cui, J.; He, Z.; Qi, B.; et al. XBP1-elicited environment by chemotherapy potentiates repopulation of tongue cancer cells by enhancing miR-22/lncRNA/KAT6B-dependent NF-kappaB signalling. Clin. Transl. Med. 2023, 13, e1166. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.Y.; Cho, K.H.; Jeong, K.J.; Cho, S.J.; Won, M.; Kim, S.H.; Cho, N.H.; Hur, G.M.; Yoon, S.H.; Park, H.W.; et al. Lysophosphatidic acid-induced amphiregulin secretion by cancer-associated fibroblasts augments cancer cell invasion. Cancer Lett. 2022, 551, 215946. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.; Ramos-Valdes, Y.; Stanton, L.A.; Ladhani, S.; Beier, F.; Dimattia, G.E. Human stanniocalcin-1 or -2 expressed in mice reduces bone size and severely inhibits cranial intramembranous bone growth. Transgenic. Res. 2010, 19, 1017–1039. [Google Scholar] [CrossRef] [PubMed]
- Kamata, T.; So, T.Y.; Ahmed, Q.; Giblett, S.; Patel, B.; Luo, J.; Reddel, R.; Pritchard, C. Fibroblast-Derived STC-1 Modulates Tumor-Associated Macrophages and Lung Adenocarcinoma Development. Cell Rep. 2020, 31, 107802. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Yuan, Y.; Liu, J.; Chen, Y.; Li, L.; Liu, S.; An, X.; Luo, R.; Long, D.; Chen, B.; et al. MSCs protect endothelial cells from inflammatory injury partially by secreting STC1. Int. Immunopharmacol. 2018, 61, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhu, J.; Guo, H.; Tang, A.; Chen, S.; Zhang, D.; Yuan, L.; Liu, G. Molecular cloning, characterization, and functional analysis of the uncharacterized C11orf96 gene. BMC Vet. Res. 2022, 18, 170. [Google Scholar] [CrossRef] [PubMed]
- Trebino, C.E.; Stock, J.L.; Gibbons, C.P.; Naiman, B.M.; Wachtmann, T.S.; Umland, J.P.; Pandher, K.; Lapointe, J.M.; Saha, S.; Roach, M.L.; et al. Impaired inflammatory and pain responses in mice lacking an inducible prostaglandin E synthase. Proc. Natl. Acad. Sci. USA 2003, 100, 9044–9049. [Google Scholar] [CrossRef] [PubMed]
- Inada, M.; Matsumoto, C.; Uematsu, S.; Akira, S.; Miyaura, C. Membrane-bound prostaglandin E synthase-1-mediated prostaglandin E2 production by osteoblast plays a critical role in lipopolysaccharide-induced bone loss associated with inflammation. J. Immunol. 2006, 177, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Hauser, C.; Saurat, J.H.; Schmitt, A.; Jaunin, F.; Dayer, J.M. Interleukin 1 is present in normal human epidermis. J. Immunol. 1986, 136, 3317–3323. [Google Scholar] [CrossRef]
- Gahring, L.C.; Buckley, A.; Daynes, R.A. Presence of epidermal-derived thymocyte activating factor/interleukin 1 in normal human stratum corneum. J. Clin. Investig. 1985, 76, 1585–1591. [Google Scholar] [CrossRef]
- Sordi, M.B.; Panahipour, L.; Gruber, R. Oral squamous carcinoma cell lysates provoke exacerbated inflammatory response in gingival fibroblasts. Clin. Oral Investig. 2023, 27, 4785–4794. [Google Scholar] [CrossRef] [PubMed]
- Urcan, E.; Scherthan, H.; Styllou, M.; Haertel, U.; Hickel, R.; Reichl, F.X. Induction of DNA double-strand breaks in primary gingival fibroblasts by exposure to dental resin composites. Biomaterials 2010, 31, 2010–2014. [Google Scholar] [CrossRef] [PubMed]
- Schweikl, H.; Spagnuolo, G.; Schmalz, G. Genetic and cellular toxicology of dental resin monomers. J. Dent. Res. 2006, 85, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.W.; Greenwell-Wild, T.; Brenchley, L.; Dutzan, N.; Overmiller, A.; Sawaya, A.P.; Webb, S.; Martin, D.; Genomics, N.N.; Computational Biology, C.; et al. Human oral mucosa cell atlas reveals a stromal-neutrophil axis regulating tissue immunity. Cell 2021, 184, 4090–4104.e4015. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.B.; Huh, C.G.; Becker, D.; Geiser, A.; Lyght, M.; Flanders, K.C.; Roberts, A.B.; Sporn, M.B.; Ward, J.M.; Karlsson, S. Transforming growth factor beta 1 null mutation in mice causes excessive inflammatory response and early death. Proc. Natl. Acad. Sci. USA 1993, 90, 770–774. [Google Scholar] [CrossRef]
- Mohanty, A.; Nam, A.; Srivastava, S.; Jones, J.; Lomenick, B.; Singhal, S.S.; Guo, L.; Cho, H.; Li, A.; Behal, A.; et al. Acquired resistance to KRAS G12C small-molecule inhibitors via genetic/nongenetic mechanisms in lung cancer. Sci. Adv. 2023, 9, eade3816. [Google Scholar] [CrossRef]
- Choi, H.I.; An, G.Y.; Baek, M.; Yoo, E.; Chai, J.C.; Lee, Y.S.; Jung, K.H.; Chai, Y.G. BET inhibitor suppresses migration of human hepatocellular carcinoma by inhibiting SMARCA4. Sci. Rep. 2021, 11, 11799. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Z.; Zhu, Q.; Ren, S.; Xu, Y.; Wang, G.; Zhou, L. Comprehensive analysis and experimental verification of the mechanism of action of T cell-mediated tumor-killing related genes in Colon adenocarcinoma. Transl. Oncol. 2024, 43, 101918. [Google Scholar] [CrossRef]
- Ouellet, V.; Negrao, J.; Skibiel, A.L.; Lantigua, V.A.; Fabris, T.F.; Marrero, M.G.; Dado-Senn, B.; Laporta, J.; Dahl, G.E. Endocrine Signals Altered by Heat Stress Impact Dairy Cow Mammary Cellular Processes at Different Stages of the Dry Period. Animals 2021, 11, 563. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.C.T.; Wong, C.K.C. Characterization of stanniocalcin-1 expression in macrophage differentiation. Transl. Oncol. 2021, 14, 100881. [Google Scholar] [CrossRef]
- Liu, F.L.; Wu, C.C.; Chang, D.M. TACE-dependent amphiregulin release is induced by IL-1beta and promotes cell invasion in fibroblast-like synoviocytes in rheumatoid arthritis. Rheumatology 2014, 53, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Yeung, B.H.; Wong, C.K. Stanniocalcin-1 regulates re-epithelialization in human keratinocytes. PLoS ONE 2011, 6, e27094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Fang, S.; Gao, H.; Zhang, X.; Gu, D.; Liu, Y.; Wan, J.; Xie, J. A critical role of AREG for bleomycin-induced skin fibrosis. Cell Biosci. 2021, 11, 40. [Google Scholar] [CrossRef]
- Sculean, A.; Gruber, R.; Bosshardt, D.D. Soft tissue wound healing around teeth and dental implants. J. Clin. Periodontol. 2014, 41 (Suppl. S15), S6–S22. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.M.; Manigrasso, M.B.; O’Connor, J.P. Cyclo-oxygenase 2 function is essential for bone fracture healing. J. Bone Miner. Res. 2002, 17, 963–976. [Google Scholar] [CrossRef]
- Robertson, G.; Xie, C.; Chen, D.; Awad, H.; Schwarz, E.M.; O’Keefe, R.J.; Guldberg, R.E.; Zhang, X. Alteration of femoral bone morphology and density in COX-2-/- mice. Bone 2006, 39, 767–772. [Google Scholar] [CrossRef]
- Chikazu, D.; Fujikawa, Y.; Fujihara, H.; Suenaga, H.; Saijo, H.; Ohkubo, K.; Ogasawara, T.; Mori, Y.; Iino, M.; Takato, T. Cyclooxygenase-2 activity is important in craniofacial fracture repair. Int. J. Oral Maxillofac. Surg. 2011, 40, 322–326. [Google Scholar] [CrossRef]
- Yamakawa, K.; Kamekura, S.; Kawamura, N.; Saegusa, M.; Kamei, D.; Murakami, M.; Kudo, I.; Uematsu, S.; Akira, S.; Chung, U.I.; et al. Association of microsomal prostaglandin E synthase 1 deficiency with impaired fracture healing, but not with bone loss or osteoarthritis, in mouse models of skeletal disorders. Arthritis Rheum. 2008, 58, 172–183. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panahipour, L.; Micucci, C.; Gelmetti, B.; Gruber, R. In Vitro Bioassay for Damage-Associated Molecular Patterns Arising from Injured Oral Cells. Bioengineering 2024, 11, 687. https://doi.org/10.3390/bioengineering11070687
Panahipour L, Micucci C, Gelmetti B, Gruber R. In Vitro Bioassay for Damage-Associated Molecular Patterns Arising from Injured Oral Cells. Bioengineering. 2024; 11(7):687. https://doi.org/10.3390/bioengineering11070687
Chicago/Turabian StylePanahipour, Layla, Chiara Micucci, Benedetta Gelmetti, and Reinhard Gruber. 2024. "In Vitro Bioassay for Damage-Associated Molecular Patterns Arising from Injured Oral Cells" Bioengineering 11, no. 7: 687. https://doi.org/10.3390/bioengineering11070687
APA StylePanahipour, L., Micucci, C., Gelmetti, B., & Gruber, R. (2024). In Vitro Bioassay for Damage-Associated Molecular Patterns Arising from Injured Oral Cells. Bioengineering, 11(7), 687. https://doi.org/10.3390/bioengineering11070687