
Wine Fermentation Performance of Indigenous Saccharomyces cerevisiae and Saccharomyces paradoxus Strains Isolated in a Piedmont Vineyard

 , , , ,
, , , ,
Abstract
:
1. Introduction
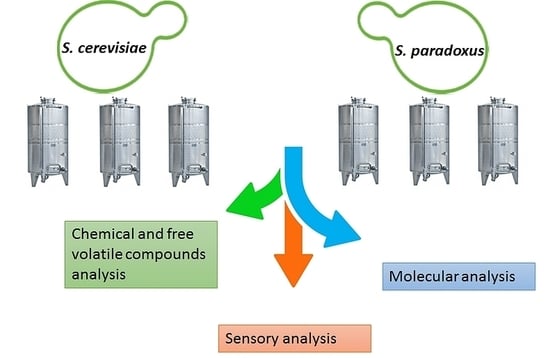
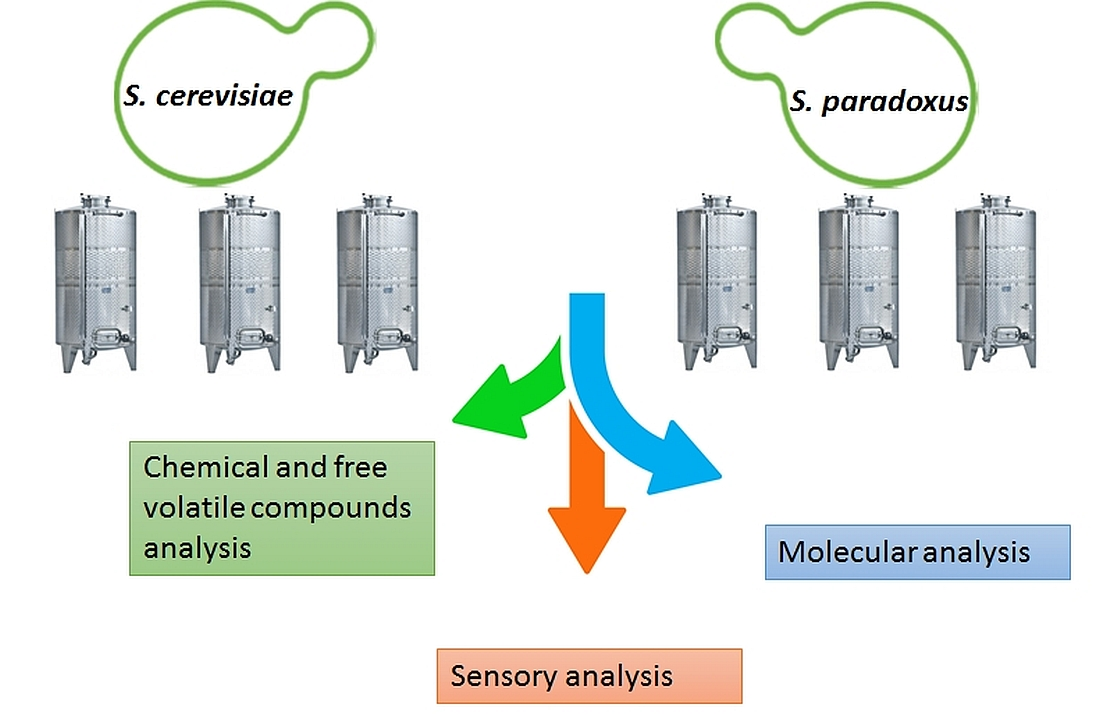
2. Materials and Methods
2.1. Yeast Strains
2.2. Vinification
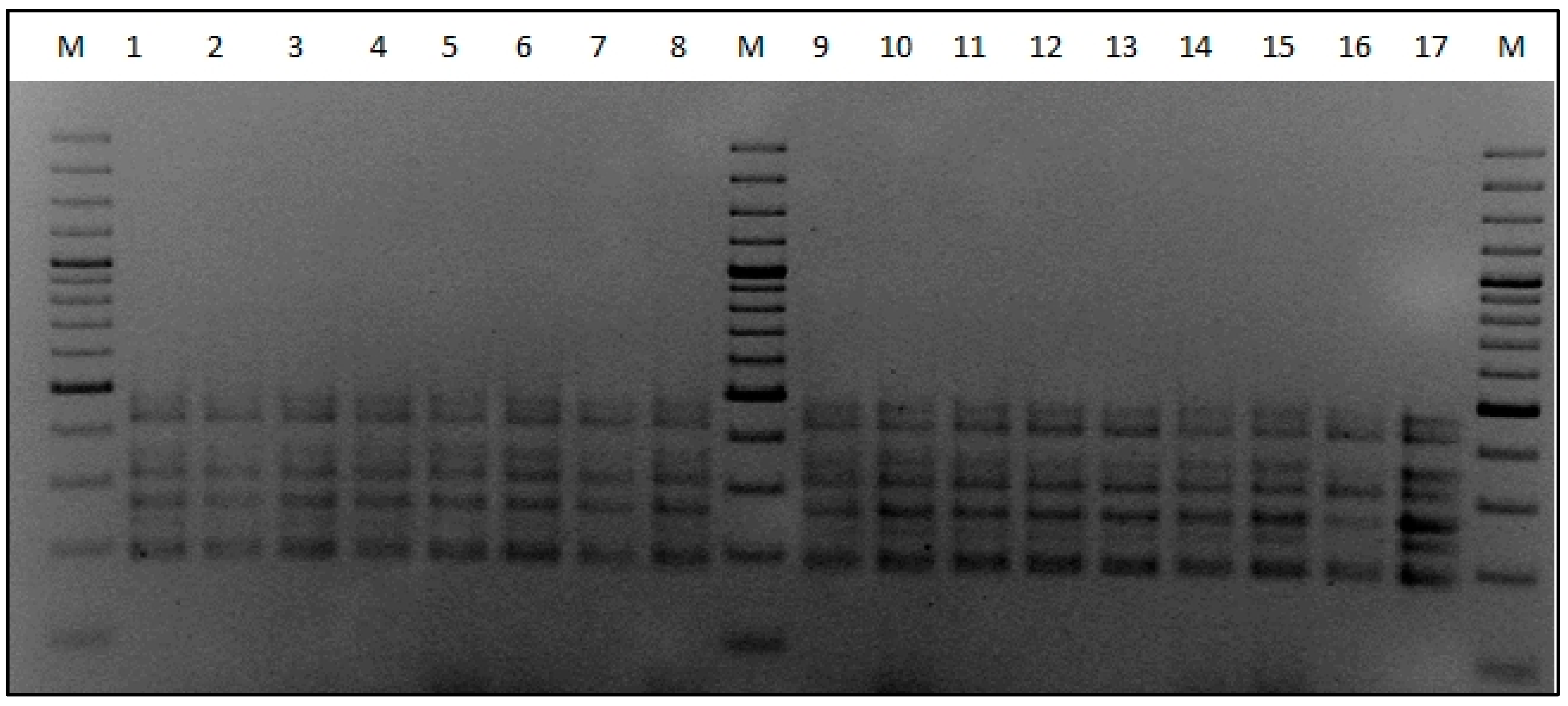
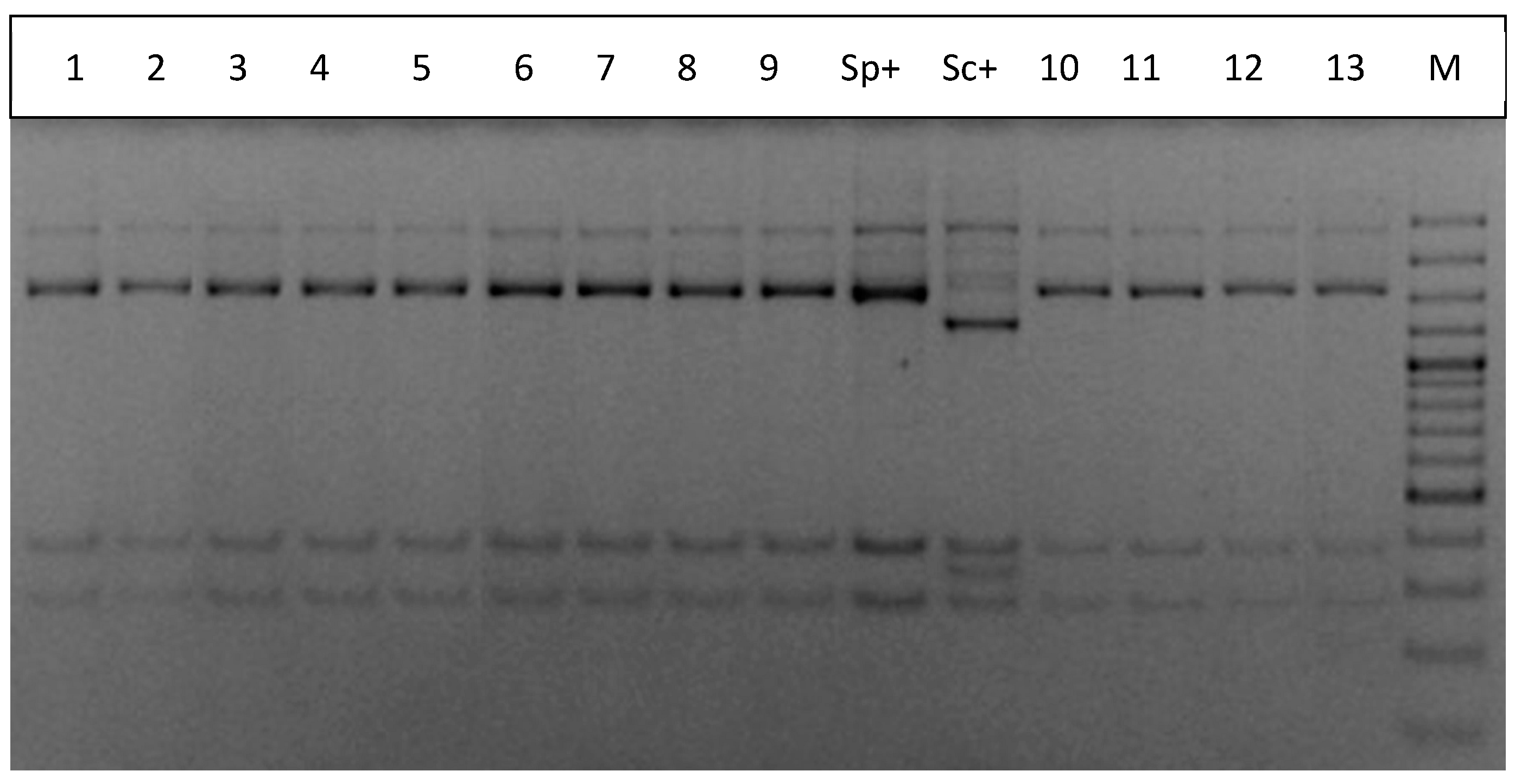
2.3. Yeast Dominance Analysis
2.4. Chemical Analysis
2.5. Sensory Analysis
2.6. Free Volatile Compounds Analysis
2.7. Statistical Analysis
3. Results and Discussion
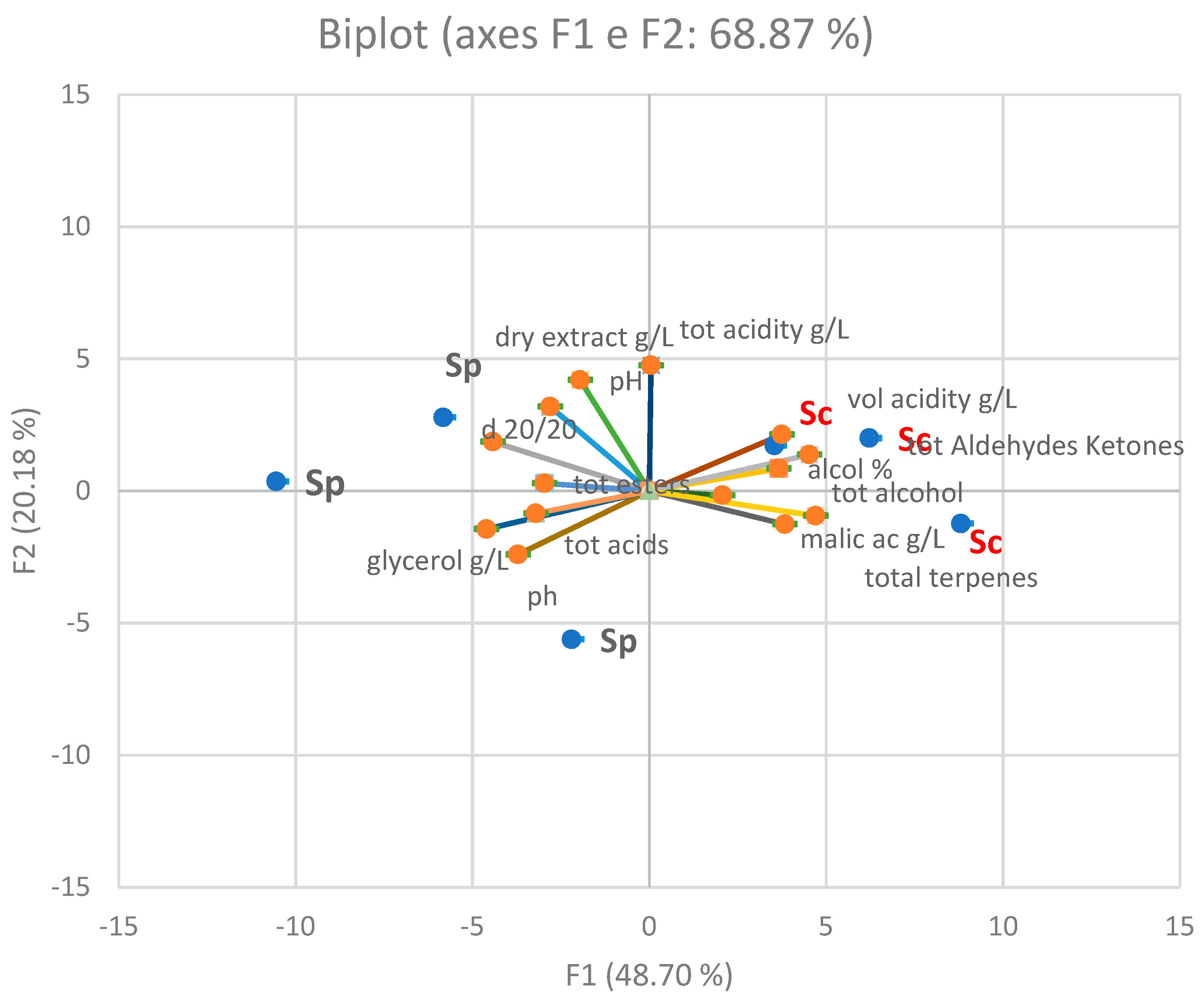
3.1. Enological Characteristics of S. paradoxus and S. cerevisiae Strains
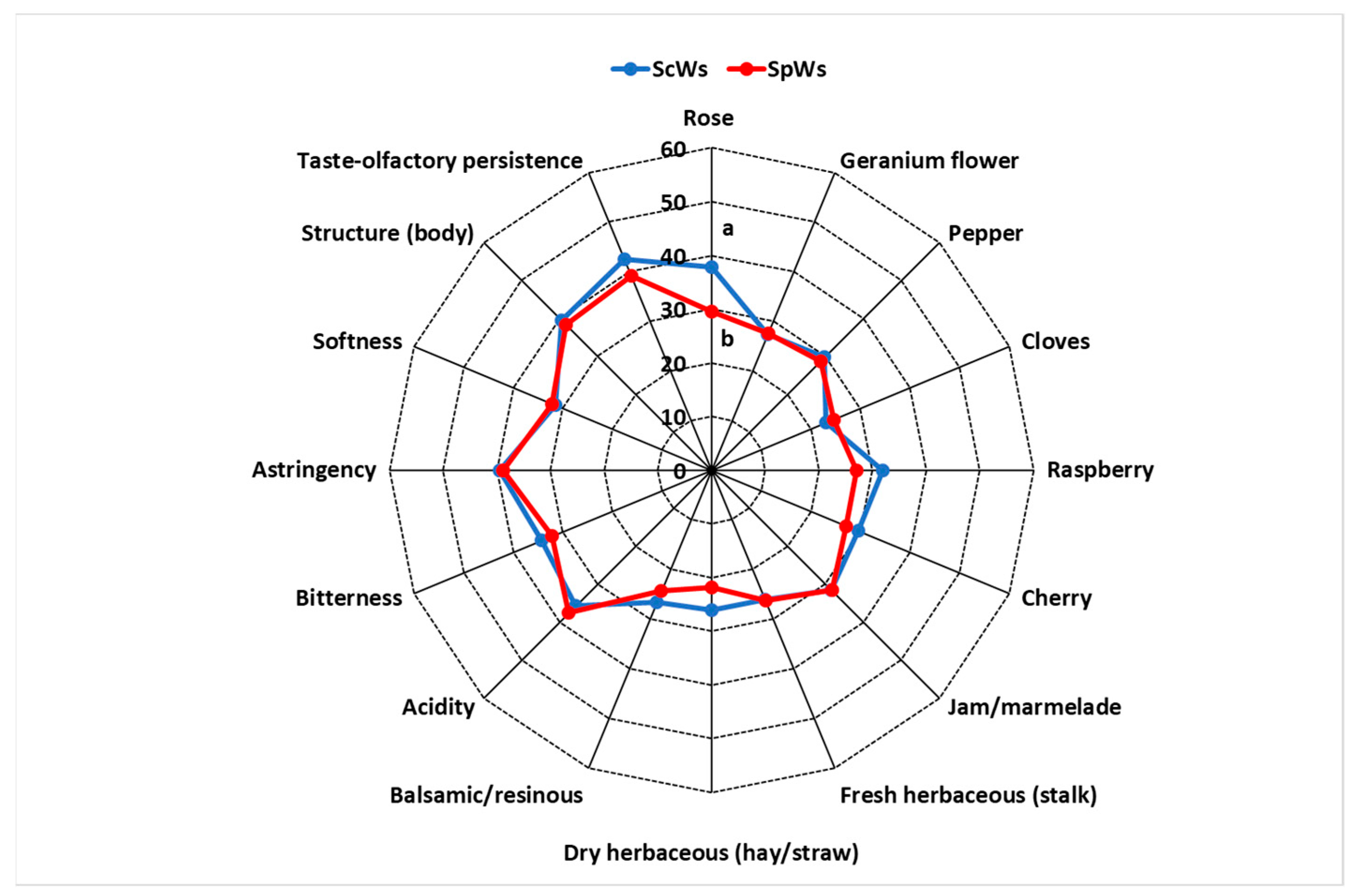
3.2. Free Volatile Compounds and Sensory Profile of the Wines
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Rousseaux, S.; Tourdot-Maréchal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine microbiome: A dynamic world of microbial interactions. Crit. Rev. Food Sci. Nutr. 2017, 57, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Tristezza, M.; Fantastico, L.; Vetrano, C.; Bleve, G.; Corallo, D.; Grieco, F.; Mita, G. Molecular and Technological Characterization of Saccharomyces cerevisiae Strains Isolated from Natural Fermentation of Susumaniello Grape Must in Apulia, Southern Italy. Int. J. Microbiol. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, F.; Veličkovska, S.K.; Dimovska, V.; Mirhosseini, H.; Spasov, H. Selection of 80 newly isolated autochthonous yeast strains from the Tikveš region of Macedonia and their impact on the quality of red wines produced from Vranec and Cabernet Sauvignon grape varieties. Food Chem. 2017, 216, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Vigentini, I.; Cardenas, S.B.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Foschino, R. Use of Native Yeast Strains for In-Bottle Fermentation to Face the Uniformity in Sparkling Wine Production. Front. Microbiol. 2017, 8, 1225. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J. Yeast systems biology: Model organism and cell factory. Biotechnol. J. 2019, 14, e1800421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redžepović, S.; Orlić, S.; Sikora, S.; Majdak, A.; Pretorius, I.S. Identification and characterization of Saccharomyces cerevisiae and Saccharomyces paradoxus strains isolated from Croatian vineyards. Lett. Appl. Microbiol. 2002, 35, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Sniegowski, P.D.; Dombrowski, P.G.; Fingerman, E. Saccharomyces cerevisiae and Saccharomyces paradoxus coexist in a natural woodland site in North America and display different levels of reproductive isolation from European conspecifics. FEMS Yeast Res. 2002, 1, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Yurkov, A.M. First Isolation of the Yeast Saccharomyces paradoxus in Western Siberia. Microbiology 2005, 74, 459–462. [Google Scholar] [CrossRef]
- Boynton, P.J.; Greig, D. The ecology and evolution of non-domesticated Saccharomyces species. Yeast 2014, 31, 449–462. [Google Scholar] [PubMed] [Green Version]
- Bleuven, C.; Dubé, A.K.; Nguyen, G.Q.; Gagnon-Arsenault, I.; Martin, H.; Landry, C.R. A collection of barcoded natural isolates of Saccharomyces paradoxus to study microbial evolutionary ecology. Microbiologyopen 2019, 8, e773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majdak, A.; Herjavec, S.; Orlić, S.; Redžepović, S.; Mirošević, N. Comparison of Wine Aroma Compounds Produced by Saccharomyces paradoxus and Saccharomyces cerevisiae Strains. Food Technol. Biotechnol. 2002, 40, 103–109. [Google Scholar]
- Orlic, S.; Redzepovic, S.; Jeromel, A.; Herjavec, S.; Iacumin, L. Influence of indigenous Saccharomyces paradoxus strains on Chardonnay wine fermentation aroma. Int. J. Food Sci. Technol. 2007, 42, 95–101. [Google Scholar] [CrossRef]
- Alonso-Del-Real, J.; Lairón-Peris, M.; Barrio, E.; Querol, A. Effect of Temperature on the Prevalence of Saccharomyces Non cerevisiae Species against a S. cerevisiae Wine Strain in Wine Fermentation: Competition, Physiological Fitness, and Influence in Final Wine Composition. Front. Microbiol. 2017, 8, 150. [Google Scholar] [CrossRef] [Green Version]
- González-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Wine Aroma Compounds in Grapes: A Critical Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 202–218. [Google Scholar]
- García, M.; Esteve-Zarzoso, B.; Crespo, J.; Cabellos, J.M.; Arroyo, T. Influence of Native Saccharomyces cerevisiae Strains from D.O. “Vinos de Madrid” in the Volatile Profile of White Wines. Fermentation 2019, 5, 94. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Vaudano, E.; Quinterno, G.; Costantini, A.; Pulcini, L.; Pessione, E.; Garcia-Moruno, E. Yeast distribution in Grignolino grapes growing in a new vineyard in Piedmont and the technological characterization of indigenous Saccharomyces spp. strains. Int. J. Food Microbiol. 2019, 289, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaudano, E.; Garcia-Moruno, E. Discrimination of Saccharomyces cerevisiae wine strains using microsatellite multiplex PCR and band pattern analysis. Food Microbiol. 2008, 25, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Cane, P. II controllo di qualità dei vini mediante HPLC: Determinazione di acidi organici. L’Enotecnico 1990, 26, 69–72. [Google Scholar]
- Zoecklein, B.; Fugelsang, K.C.; Gump, B.; Nury, F.S. Wine Analysis and Production; Springer: New York, NY, USA, 1999. [Google Scholar]
- Cravero, M.C.; Bonello, F.; Tsolakis, C.; Piano, F.; Borsa, D. Comparison between Nero d’Avola wines produced with grapes grown in Sicily and Tuscany. Ital. J. Food Sci. 2012, 24, 384–387. [Google Scholar]
- Campo, E.; Do, B.V.; Ferreira, V.; Valentin, D. Aroma properties of young Spanish monovarietal white wines: A study using sorting task, list of terms and frequency of citation. Aust. J. Grape Wine Res. 2008, 14, 104–115. [Google Scholar] [CrossRef]
- Wirth, J.; Morel-Salmi, C.; Souquet, J.M.; Dieval, J.B.; Aagaard, O.; Vidal, S.; Fulcrand, H.; Cheynier, V. The impact of oxygen exposure before and after bottling on the polyphenolic composition of red wines. Food Chem. 2010, 123, 107–116. [Google Scholar] [CrossRef]
- Guinard, J.X.; Noble, A.C. Proposition d’une terminologie pour une description analytique de l’arome des vins. Sc. Alim. 1986, 6, 657–662. [Google Scholar]
- Petrozziello, M.; Guaita, M.; Motta, S.; Panero, L.; Bosso, A. Analytical and Sensory Characterization of the Aroma of “Langhe D.O.C. Nebbiolo” Wines: Influence of the Prefermentative Cold Maceration with Dry Ice. J. Food Sci. 2011, 76, C525–C534. [Google Scholar] [CrossRef] [PubMed]
- Scanes, K.T.; Hohrnann, S.; Prior, B.A. Glycerol Production by the Yeast Saccharomyces cerevisiae and its Relevance to Wine: A Review. S. Afr. J. Enol. Vitic. 2017, 19, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Orlic, S.; Arroyo-Lopez, F.N.; Huic-Babic, K.; Lucilla, I.; Querol, A.; Barrio, E. A comparative study of the wine fermentation performance of Saccharomyces paradoxus under different nitrogen concentrations and glucose/fructose ratios. J. Appl. Microbiol. 2010, 108, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozturk, B.; Anli, E. Different techniques for reducing alcohol levels in wine: A review. In Proceedings of the BIO Web of Conferences, Mendoza, Argentina, 9–14 November 2014; Aurand, J.-M., Ed.; EDP Sciences: Les Ulis Cedex, France, 2014; Volume 3, p. 02012. [Google Scholar] [CrossRef]
- Caballero, A.; Segura, A. The quest for lower alcoholic wines. Microb. Biotechnol. 2017, 10, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Cravero, M.C.; Ubigli, M. L’analisi sensoriale e la tipicità del Grignolino. OICCE TIMES 2003, 4, 13–18. [Google Scholar]
- Carrau, F.M.; Medina, K.; Boido, E.; Farina, L.; Gaggero, C.; Dellacassa, E.; Versini, G.; Henschke, P.A. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 2005, 243, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Orte, P.; Cersosimo, M.; Loscos, N.; Cacho, J.; Garcia-Moruno, E.; Ferreira, V. Aroma development from non-floral grape precursors by wine lactic acid bacteria. Food Res. Int. 2009, 42, 773–781. [Google Scholar] [CrossRef]
- Ferreira, V. A base química do aroma do vinho: Moléculas e sensações olfactogustativas. Parte 1: Álcool e efeito do tampão aromático. Rev. Internet Vitic. Enol. 2009, 9, 1–7. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SpW | ScW | |
|---|---|---|
| Density 20/20 | 0.99456 | 0.99368 |
| Ethanol % v/v | 12.73 | 13.29 |
| Residual sugars g/L | ≤1 | ≤1 |
| Titratable acidity g/L | 6.32 | 6.35 |
| Volatile acidity g/L | 0.19 | 0.25 |
| Malic acid g/L | 2.50 a | 3.27 b |
| pH | 3.54 | 3.52 |
| Glycerol g/L | 8.62 a | 6.70 b |
| SpW | ScW | |
|---|---|---|
| Alcohols | ||
| Isoamylalcohol | 12,261 ± 1447 | 13,140 ± 2888 |
| Cis-3-hexenol | 73 ± 6 | 57 ± 21 |
| 1- hexanol | 1359 ± 66 | 1402 ± 298 |
| benzylalcohol | 7 ± 2 | 7 ± 1 |
| 2 phenylethanol | 14,337 ± 1000 | 17,465 ± 5000 |
| total alcohol | 28,037 | 32,071 |
| Esters | ||
| ethylhexanoate | 279 ± 60 | 185 ± 31 |
| isoamilacetatate | 406 ± 151 | 418 ± 27 |
| ethyllactate | 1284 ± 118 | 585 ± 42 |
| ethyloctanoate | 267 ± 62 | 177 ± 13 |
| ethylhexanoate | 271 ± 46 | 185 ± 31 |
| ethyldecanoate | 55 ± 20 | 37 ± 4 |
| diethylsuccinate | 1187 ± 360 | 1351 ± 293 |
| ethylpalmitate | 12 ± 1 | 16 ± 4 |
| total esters | 3761 | 2954 |
| Acids | ||
| isovaleric acid | 150 ± 11 | 99 ± 24 |
| octanoic acid | 1869 ± 242 | 1559 ± 342 |
| decanoic acid | 444 ± 172 | 377 ± 57 |
| lauric acid | 37 ± 13 | 29 ± 4 |
| total acids | 2500 | 2064 |
| Aldehydes ketones | ||
| benzaldehyde | 6 ± 2 | 8 ± 2 |
| butyrolactone | 36 ± 20 | 40 ± 27 |
| methoxyacetophenone | 0 | 248 ± 95 |
| vanillin | 0 | 14 ± 1 |
| β−damascenone | 11 ± 5 | 8 ± 1 |
| total Aldehydes Ketones | 53a | 318b |
| Terpenic compounds | ||
| linalool | 0 a | 16 ± 2 b |
| cis-linalooloxide | 4 ± 1 | 5 ± 1 |
| citronellol | 14 ± 0 | 24 ± 2 |
| HO trienol | 0 | 7 ± 5 |
| alpha terpineol | 10 ± 1 | 11 ± 2 |
| geranic acid | 40 ± 13 | 43 ± 4 |
| Total terpenes | 68a | 106b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costantini, A.; Cravero, M.C.; Panero, L.; Bonello, F.; Vaudano, E.; Pulcini, L.; Garcia-Moruno, E. Wine Fermentation Performance of Indigenous Saccharomyces cerevisiae and Saccharomyces paradoxus Strains Isolated in a Piedmont Vineyard. Beverages 2021, 7, 30. https://doi.org/10.3390/beverages7020030
Costantini A, Cravero MC, Panero L, Bonello F, Vaudano E, Pulcini L, Garcia-Moruno E. Wine Fermentation Performance of Indigenous Saccharomyces cerevisiae and Saccharomyces paradoxus Strains Isolated in a Piedmont Vineyard. Beverages. 2021; 7(2):30. https://doi.org/10.3390/beverages7020030
Chicago/Turabian StyleCostantini, Antonella, Maria Carla Cravero, Loretta Panero, Federica Bonello, Enrico Vaudano, Laura Pulcini, and Emilia Garcia-Moruno. 2021. "Wine Fermentation Performance of Indigenous Saccharomyces cerevisiae and Saccharomyces paradoxus Strains Isolated in a Piedmont Vineyard" Beverages 7, no. 2: 30. https://doi.org/10.3390/beverages7020030
APA StyleCostantini, A., Cravero, M. C., Panero, L., Bonello, F., Vaudano, E., Pulcini, L., & Garcia-Moruno, E. (2021). Wine Fermentation Performance of Indigenous Saccharomyces cerevisiae and Saccharomyces paradoxus Strains Isolated in a Piedmont Vineyard. Beverages, 7(2), 30. https://doi.org/10.3390/beverages7020030