
Cultivation of Protozoa Parasites In Vitro: Growth Potential in Conventional Culture Media versus RPMI-PY Medium

 , , , , ,
, , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Media Preparation
2.1.1. Evans’s Modified Tobie Medium
2.1.2. Novy–MacNeal–Nicolle Medium
2.1.3. RPMI-PY Medium
2.2. Protozoa Parasites
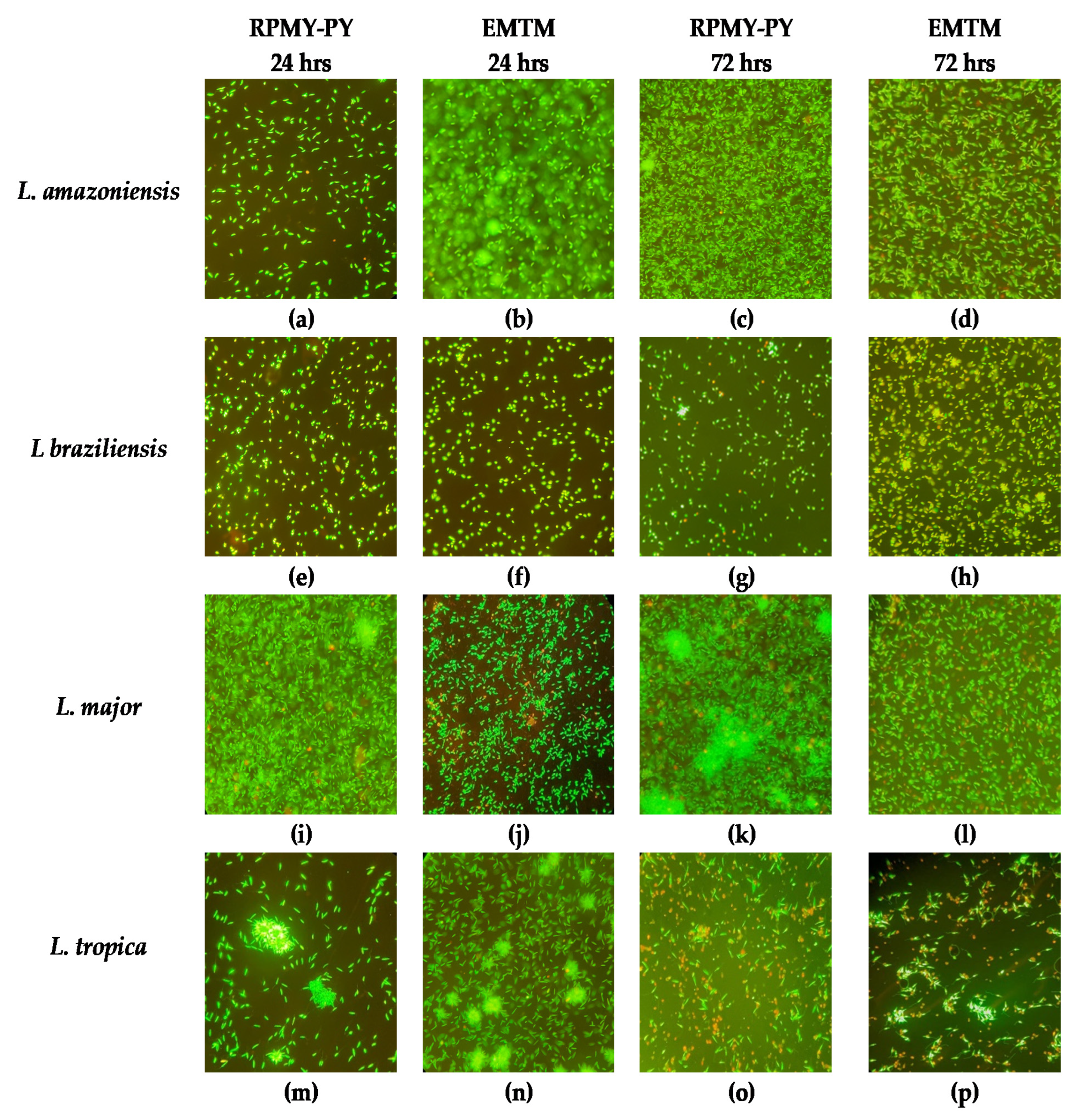
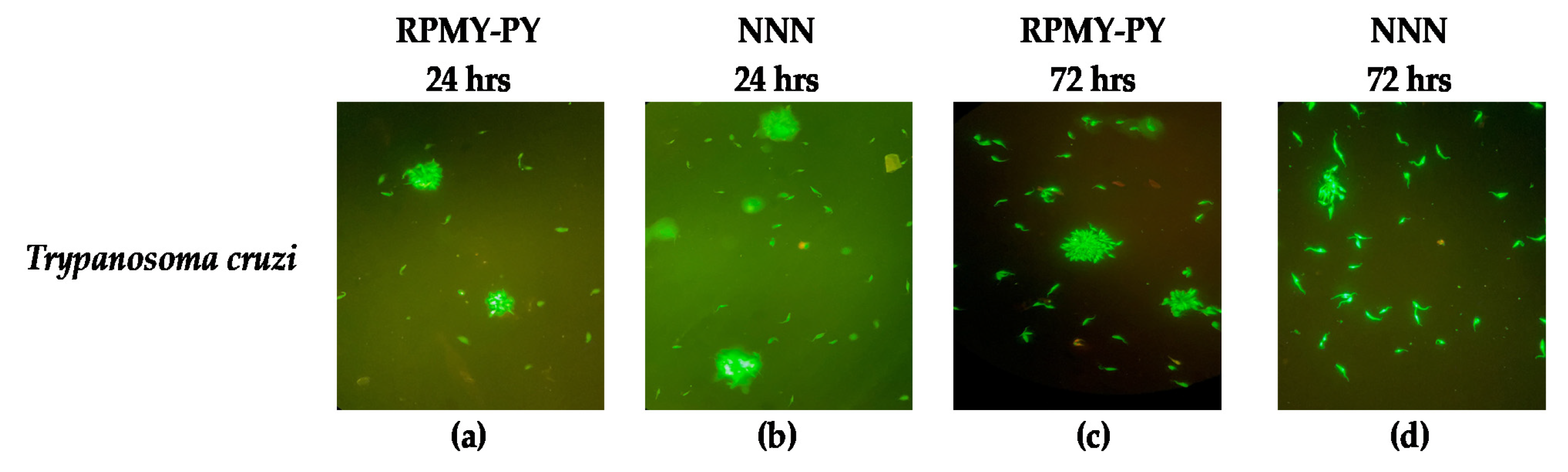
2.3. Morphological Analysis
2.4. Statistical Analysis
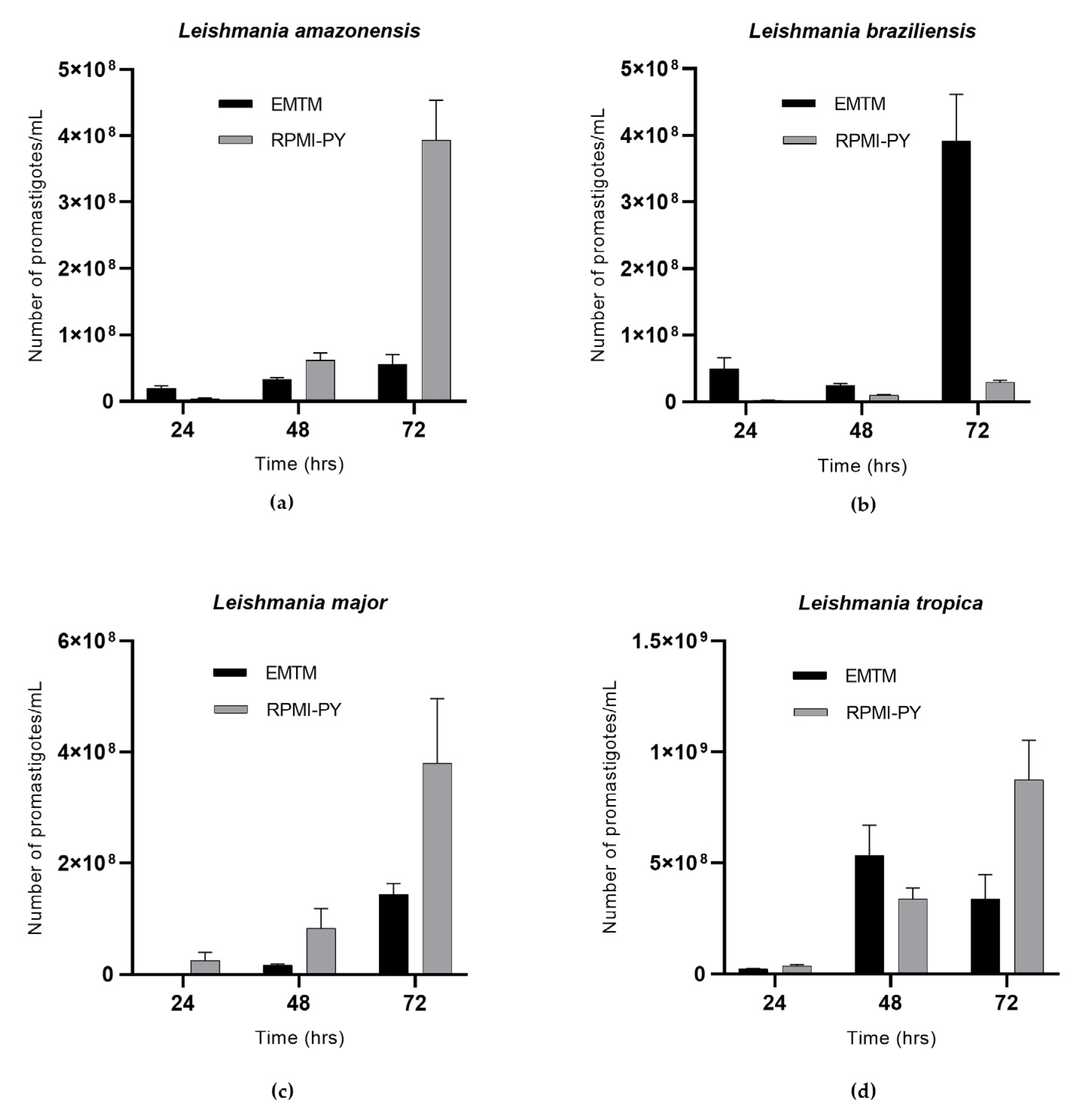
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schuster, F.L.; Sullivan, J.J. Cultivation of Clinically Significant Hemoflagellates. Clin. Microbiol. Rev. 2002, 15, 374–389. [Google Scholar] [CrossRef] [Green Version]
- Kaufer, A.; Ellis, J.; Stark, D.; Barratt, J. The Evolution of Trypanosomatid Taxonomy. Parasites Vectors 2017, 10, 287. [Google Scholar] [CrossRef] [Green Version]
- Herrera, G.; Barragán, N.; Luna, N.; Martínez, D.; De Martino, F.; Medina, J.; Niño, S.; Páez, L.; Ramírez, A.; Vega, L.; et al. An Interactive Database of Leishmania Species Distribution in the Americas. Sci. Data 2020, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gow, I.; Smith, N.C.; Stark, D.; Ellis, J. Laboratory Diagnostics for Human Leishmania Infections: A Polymerase Chain Reaction-Focussed Review of Detection and Identification Methods. Parasites Vectors 2022, 15, 412. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.M.; Oghumu, S.; Gupta, G.; McGwire, B.S.; Drew, M.E.; Satoskar, A.R. Mechanisms of Cellular Invasion by Intracellular Parasites. Cell Mol. Life Sci. 2014, 71, 1245–1263. [Google Scholar] [CrossRef] [Green Version]
- Steverding, D. The History of Leishmaniasis. Parasit Vectors 2017, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Masucci, M.; Donato, G.; Persichetti, M.F.; Priolo, V.; Castelli, G.; Bruno, F.; Pennisi, M.G. Hemogram Findings in Cats from an Area Endemic for Leishmania Infantum and Feline Immunodeficiency Virus Infections. Vet. Sci. 2022, 9, 508. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Kjos, S.; Yabsley, M.J.; Montgomery, S.P. Trypanosoma Cruzi and Chagas’ Disease in the United States. Clin. Microbiol. Rev. 2011, 24, 655–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carod-Artal, F.J. American Trypanosomiasis. Handb. Clin. Neurol. 2013, 114, 103–123. [Google Scholar] [CrossRef]
- Macaluso, G.; Grippi, F.; Di Bella, S.; Blanda, V.; Gucciardi, F.; Torina, A.; Guercio, A.; Cannella, V. A Review on the Immunological Response against Trypanosoma Cruzi. Pathogens 2023, 12, 282. [Google Scholar] [CrossRef]
- De-Simone, S.G.; Bourguignon, S.C.; Gonçalves, P.S.; Lechuga, G.C.; Provance, D.W. Metabolic Alteration of Trypanosoma Cruzi during Differentiation of Epimastigote to Trypomastigote Forms. Pathogens 2022, 11, 268. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Toloza, G.; Ferreira, A. Trypanosoma Cruzi Evades the Complement System as an Efficient Strategy to Survive in the Mammalian Host: The Specific Roles of Host/Parasite Molecules and Trypanosoma Cruzi Calreticulin. Front. Microbiol. 2017, 8, 1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verso, M.G.; Vitale, F.; Castelli, G.; Bruno, F.; Migliazzo, A.; Bongiorno, M.R.; Santi, F.; Pistone, G.; Amodio, E.; Picciotto, D. Suspected Cutaneous Leishmaniasis in a Sample of Westerns Sicily Residents: What Correlation with Occupation? Med. Lav. 2017, 108, 123–129. [Google Scholar] [CrossRef]
- Sundar, S.; Rai, M. Laboratory Diagnosis of Visceral Leishmaniasis. Clin. Vaccine Immunol. 2002, 9, 951–958. [Google Scholar] [CrossRef] [Green Version]
- Castelli, G.; Bruno, F.; Reale, S.; Catanzaro, S.; Valenza, V.; Vitale, F. Molecular Diagnosis of Leishmaniasis: Quantification of Parasite Load by a Real-Time PCR Assay with High Sensitivity. Pathogens 2021, 10, 865. [Google Scholar] [CrossRef]
- Srivastava, P.; Dayama, A.; Mehrotra, S.; Sundar, S. Diagnosis of Visceral Leishmaniasis. Trans. R Soc. Trop. Med. Hyg. 2011, 105, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visvesvara, G.S.; Garcia, L.S. Culture of Protozoan Parasites. Clin. Microbiol. Rev. 2002, 15, 327–328. [Google Scholar] [CrossRef] [Green Version]
- Merlen, T.; Sereno, D.; Brajon, N.; Rostand, F.; Lemesre, J.L. Leishmania Spp: Completely Defined Medium without Serum and Macromolecules (CDM/LP) for the Continuous in Vitro Cultivation of Infective Promastigote Forms. Am. J. Trop. Med. Hyg. 1999, 60, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rodrigues, I.A.; da Silva, B.A.; dos Santos, A.L.S.; Vermelho, A.B.; Alviano, C.S.; Rosa, M.d.S.S. A New Experimental Culture Medium for Cultivation of Leishmania Amazonensis: Its Efficacy for the Continuous in Vitro Growth and Differentiation of Infective Promastigote Forms. Parasitol. Res. 2010, 106, 1249–1252. [Google Scholar] [CrossRef]
- Habibzadeh, S.; Doroud, D.; Taheri, T.; Seyed, N.; Rafati, S. Leishmania Parasite: The Impact of New Serum-Free Medium as an Alternative for Fetal Bovine Serum. Iran. Biomed. J. 2021, 25, 349–358. [Google Scholar] [CrossRef]
- Ladopoulos, T.; Ntais, P.; Tsirigotakis, N.; Dokianakis, E.; Antoniou, M. The Proliferation Potential of Promastigotes of the Main Leishmania Species of the Old World in NNN Culture Medium Prepared Using Blood of Four Different Mammals. Exp. Parasitol. 2015, 157, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.C. Trypanosoma Cruzi: Intracellular Stages Grown in a Cell-Free Medium at 37 C. Exp. Parasitol. 1978, 45, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Balanco, J.M.; Pral, E.M.; da Silva, S.; Bijovsky, A.T.; Mortara, R.A.; Alfieri, S.C. Axenic Cultivation and Partial Characterization of Leishmania Braziliensis Amastigote-like Stages. Parasitology 1998, 116, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Limoncu, M.E.; Balcioğlu, I.C.; Yereli, K.; Ozbel, Y.; Ozbilgin, A. A New Experimental in Vitro Culture Medium for Cultivation of Leishmania Species. J. Clin. Microbiol. 1997, 35, 2430–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, D.A. In Vitro Methods for Parasite Cultivation. Leishmania; Taylor, A.E.R., Baker, J.R., Eds.; Academic Press Inc.: London, UK, 1987; pp. 52–75. [Google Scholar]
- Castelli, G.; Galante, A.; Verde, V.L.; Migliazzo, A.; Reale, S.; Lupo, T.; Piazza, M.; Vitale, F.; Bruno, F. Evaluation of Two Modified Culture Media for Leishmania Infantum Cultivation Versus Different Culture Media. J. Parasitol. 2014, 100, 228–230. [Google Scholar] [CrossRef]
- Hendricks, L.D.; Wood, D.E.; Hajduk, M.E. Haemoflagellates: Commercially Available Liquid Media for Rapid Cultivation. Parasitology 1978, 76, 309–316. [Google Scholar] [CrossRef]
- Palomino, J.C. Peptone-Yeast Autolysate-Fetal Bovine Serum 10, a Simple, Inexpensive Liquid Medium for Cultivation of Leishmania Spp. J. Clin. Microbiol. 1982, 15, 949–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasew, G.; Kebede, A.; Wolday, D.; Gadisa, E.; Britton, S.; Eidsmo, L.; Akuffo, H. Low-Cost Liquid Medium for in Vitro Cultivation of Leishmania Parasites in Low-Income Countries. Glob. Health Action 2009, 2, 2046. [Google Scholar] [CrossRef]
- Ali, S.A.; Iqbal, J.; Ahmad, B.; Masoom, M. A Semisynthetic Fetal Calf Serum-Free Liquid Medium for in Vitro Cultivation of Leishmania Promastigotes. Am. J. Trop. Med. Hyg. 1998, 59, 163–165. [Google Scholar] [CrossRef]
- Limoncu, M.E.; Özbilgin, A.; Balcioḡlu, İ.C.; Özbel, Y. Evaluation of Three New Culture Media for the Cultivation and Isolation of Leishmania Parasites. J. Basic Microbiol. 2004, 44, 197–202. [Google Scholar] [CrossRef]
- Berens, R.L.; Brun, R.; Krassner, S.M. A Simple Monophasic Medium for Axenic Culture of Hemoflagellates. J. Parasitol. 1976, 62, 360. [Google Scholar] [CrossRef] [PubMed]
- Abhishek, S.; Chaturvedi, S.K.; Gupta, A.K.; Ahmed, G.; Das, V.R.; Narayan, S. A Simple Monophasic LGPY Medium for Routine Maintenance of Leishmania Donovani Promastigotes. Parasitol. Res. 2021, 120, 2969–2971. [Google Scholar] [CrossRef]
- Tobie, E.J.; Von Brand, T.; Mehlman, B. Cultural and Physiological Observations on Trypanosoma Rhodesiense and Trypanosoma Gambiense. J. Parasitol. 1950, 36, 48–54. [Google Scholar] [CrossRef]
- Evans, D.A. An Inexpensive Easily Available Replacement for Foetal Calf Serum in Media for the in Vitro Cultivation of Leishmania Spp. Z. Für Parasitenkd. 1986, 72, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Evaluation of Modified Novy-MacNeal-Nicolle Medium for Isolation of Leishmania Parasites from Cutaneous Lesions of Patients in Iran. Available online: https://scialert.net/abstract/?doi=jp.2009.56.62 (accessed on 12 January 2023).
- Liegler, T.J.; Hyun, W.; Yen, T.S.; Stites, D.P. Detection and Quantification of Live, Apoptotic, and Necrotic Human Peripheral Lymphocytes by Single-Laser Flow Cytometry. Clin. Diagn. Lab. Immunol. 1995, 2, 369–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, G.; Bruno, F.; Vitale, F.; Roberti, M.; Colomba, C.; Giacomini, E.; Guidotti, L.; Cascio, A.; Tolomeo, M. In Vitro Antileishmanial Activity of Trans-Stilbene and Terphenyl Compounds. Exp. Parasitol. 2016, 166, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhagat, S.; Parikh, Y.; Singh, S.; Sengupta, S. A Novel Nanoliposomal Formulation of the FDA Approved Drug Halofantrine Causes Cell Death of Leishmania Donovani Promastigotes in Vitro. Colloids Surf. A Physicochem. Eng. Asp. 2019, 582, 123852. [Google Scholar] [CrossRef]
- Bruno, F.; Castelli, G.; Migliazzo, A.; Piazza, M.; Galante, A.; Verde, V.; Calderone, S.; Nucatolo, G.; Vitale, F. Cytotoxic Screening and in Vitro Evaluation of Pentadecane Compound against Leishmania Infantum Promastigotes and Amastigotes. J. Parasitol. 2015, 101, 701–705. [Google Scholar] [CrossRef] [Green Version]
- Rebouças-Silva, J.; Celes, F.S.; Lima, J.B.; Barud, H.S.; de Oliveira, C.I.; Berretta, A.A.; Borges, V.M. Parasite Killing of Leishmania (V) Braziliensis by Standardized Propolis Extracts. Evid. Based Complement. Altern. Med. 2017, 2017, 6067172. [Google Scholar] [CrossRef] [Green Version]
- Steiger, R.F.; Steiger, E. Cultivation of Leishmania Donovani and Leishmania Braziliensis in Defined Media: Nutritional Requirements. J. Protozool. 1977, 24, 437–441. [Google Scholar] [CrossRef]
- Niño, A.; Camacho, M. Leishmania (Viannia) Braziliensis Growth in Vitro Culture Relies More on Folic Acid Availability than Leihsmania (Leishmania) Amazonensis. Mem. Inst. Oswaldo Cruz. 2005, 100, 309–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaddu, J.B.; Nyamori, M.P. Nutrient Broth for the Cultivation of Leishmania. J. Parasitol. 1990, 76, 265–266. [Google Scholar] [CrossRef] [PubMed]
- Ozbilgin, A.; Ozbel, Y.; Alkan, M.Z.; Atambay, M.; Ozcel, M.A. Cultivation of Leishmania Sp. in Nutrient Broth. J. Egypt Soc. Parasitol. 1995, 25, 437–441. [Google Scholar] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castelli, G.; Oliveri, E.; Valenza, V.; Giardina, S.; Facciponte, F.; La Russa, F.; Vitale, F.; Bruno, F. Cultivation of Protozoa Parasites In Vitro: Growth Potential in Conventional Culture Media versus RPMI-PY Medium. Vet. Sci. 2023, 10, 252. https://doi.org/10.3390/vetsci10040252
Castelli G, Oliveri E, Valenza V, Giardina S, Facciponte F, La Russa F, Vitale F, Bruno F. Cultivation of Protozoa Parasites In Vitro: Growth Potential in Conventional Culture Media versus RPMI-PY Medium. Veterinary Sciences. 2023; 10(4):252. https://doi.org/10.3390/vetsci10040252
Chicago/Turabian StyleCastelli, Germano, Eugenia Oliveri, Viviana Valenza, Susanna Giardina, Flavia Facciponte, Francesco La Russa, Fabrizio Vitale, and Federica Bruno. 2023. "Cultivation of Protozoa Parasites In Vitro: Growth Potential in Conventional Culture Media versus RPMI-PY Medium" Veterinary Sciences 10, no. 4: 252. https://doi.org/10.3390/vetsci10040252
APA StyleCastelli, G., Oliveri, E., Valenza, V., Giardina, S., Facciponte, F., La Russa, F., Vitale, F., & Bruno, F. (2023). Cultivation of Protozoa Parasites In Vitro: Growth Potential in Conventional Culture Media versus RPMI-PY Medium. Veterinary Sciences, 10(4), 252. https://doi.org/10.3390/vetsci10040252