
Health-Related Outcomes and Molecular Methods for the Characterization of A1 and A2 Cow’s Milk: Review and Update



Abstract
:Simple Summary
Abstract
1. Introduction
2. Milk Composition
3. Casein
4. Distinction between A1 and A2 β-Casein
5. Health-Related Outcomes of A1 and A2 Milk
6. New Molecular and Biochemical Methods for Characterization A1/A2 Milk
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Truswell, A.S. The A2 milk case: A critical review. Eur. J. Clin. Nutr. 2005, 59, 623–631. [Google Scholar] [CrossRef]
- Elliott, R.B.; Martin, J.M. Dietary protein: A trigger of insulin-dependent diabetes in the BB rat? Diabetologia 1984, 26, 297–299. [Google Scholar] [CrossRef]
- Elliott, R.B. Epidemiology of diabetes in Polynesia & New Zealand. In Epidemiology and Etiology of Insulin-Dependent Diabetes in the Young; Levy-Marchal, C., Czernichow, P., Eds.; Pediatric and Adolescent Endocrinology; Karger: Basel, Switzerland; New York, NY, USA, 1992; pp. 66–71. [Google Scholar]
- McLachlan, C.N.S. β-casein A1, ischaemic heart disease mortality, and other illnesses. Med. Hypotheses 2001, 56, 262–272. [Google Scholar] [CrossRef]
- European Food Safety Authority. Review of the potential health impact of β-casomorphins and related peptides. EFSA J. 2009, 7, 231. [Google Scholar] [CrossRef]
- de Gaudry, D.K.; Lohner, S.; Bischoff, K.; Schmucker, C.; Hoerrlein, S.; Roeger, C.; Schwingshackl, L.; Meerpohl, J.J. A1- and A2 beta-casein on health-related outcomes: A scoping review of animal studies. Eur. J. Nutr. 2022, 61, 1–21. [Google Scholar] [CrossRef]
- Gonzales-Malca, J.A.; Tirado-Kulieva, V.A.; Abanto-Lopez, M.S.; Aldana-Juarez, W.L.; Palacios-Zapata, C.M. Worldwide research on the health effects of bovine milk containing A1 and A2 beta-casein: Unraveling the current scenario and future trends through bibliometrics and text mining. Curr. Res. Food. Sci. 2023, 7, 100602. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Falvo, M.J. Protein—Which is best? J. Sports Sci. Med. 2004, 3, 118–130. [Google Scholar]
- Yalçin, A.S. Emerging therapeutic potential of whey proteins and peptides. Curr. Pharm. Des. 2006, 12, 1637–1643. [Google Scholar] [CrossRef]
- Parodi, P.W. A role for milk proteins and their peptides in cancer prevention. Curr. Pharm. Des. 2007, 13, 813–828. [Google Scholar] [CrossRef]
- Fox, P.F.; McSweeney, P.L.H.; O’Mahony, J.A.; Uniacke-Lowe, T. Dairy Chemistry and Biochemistry, 2nd ed.; Springer: Cham, Switzerland, 2015; ISBN 978-3-319-14892-2. [Google Scholar] [CrossRef]
- Phadungath, C. Casein micelle structure: A concise review. Songklanakarin J. Sci. Technol. 2005, 27, 201–212. [Google Scholar]
- Demirel, A.F.; Çak, B. Discussions of Effect A1 and A2 Milk Beta-Casein Gene on Health. Approaches Poult. Dairy Vet. Sci. 2018, 3, 216–221. [Google Scholar] [CrossRef]
- Walzem, R.L.; Dillard, C.J.; German, J.B. Whey components: Millennia of evolution create functionalities for mammalian nutrition: What we know and what we may be overlooking. Crit. Rev. Food Sci. Nutr. 2002, 42, 353–375. [Google Scholar] [CrossRef]
- Rahimi, Z.; Gholami, M.; Rahimi, Z.; Yari, K. Evaluation of beta-casein locus for detection of A1 and A2 alleles frequency using allele specific PCR in native cattle of Kermanshah, Iran. Biharean Biol. 2015, 9, 85–87. [Google Scholar]
- Anema, S.G. Heat-induced changes in caseins and casein micelles, including interactions with denatured whey proteins. Int. Dairy J. 2021, 122, 105–136. [Google Scholar] [CrossRef]
- Bogahawaththa, D.; Trajkovska, B.; Markoska, T.; Vasiljevic, T. Effects of pressurized thermal processing on native proteins of raw skim milk and its concentrate. J. Dairy Sci. 2021, 104, 2834–2842. [Google Scholar] [CrossRef]
- Akkerman, M.; Johansen, L.B.; Rauh, V.; Sørensen, J.; Larsen, L.B.; Poulsen, N.A. Relationship between casein micelle size, protein composition and stability of UHT milk. Int. Dairy J. 2021, 112, 104856. [Google Scholar] [CrossRef]
- Yuan, H.; Han, S.; Zhang, S.; Xue, Y.; Zhang, Y.; Lu, H.; Wang, S. Microbial Properties of Raw Milk throughout the Year and Their Relationships to Quality Parameters. Foods 2022, 11, 3077. [Google Scholar] [CrossRef]
- Murphy, S.C.; Martin, N.H.; Barbano, D.M.; Wiedmann, M. Influence of raw milk quality on processed dairy products: How do raw milk quality test results relate to product quality and yield? J. Dairy Sci. 2016, 99, 10128–10149. [Google Scholar] [CrossRef]
- Daniloski, D.; Cunha, N.M.D.; McCarthy, N.A.; O’Callaghan, T.F.; McParland, S.; Vasiljevic, T. Health-related outcomes of genetic polymorphism of bovine β-casein variants: A systematic review of randomised controlled trials. Trends Food Sci. Technol. 2021, 111, 233–248. [Google Scholar] [CrossRef]
- Li, X.; Spencer, G.W.K.; Ong, L.; Gras, S.L. Beta casein proteins—A comparison between caprine and bovine milk. Trends Food Sci. Technol. 2022, 121, 30–43. [Google Scholar] [CrossRef]
- de Kruif, C.G.; Huppertz, T.; Urban, V.S.; Petukhov, A.V. Casein micelles and their internal structure. Adv. Colloid Interface Sci. 2012, 171–172, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Caseins and the casein micelle: Their biological functions, structures, and behavior in foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, T. Chemistry of the Caseins. In Advanced Dairy Chemistry: Volume 1A: Proteins: Basic Aspects, 4th ed.; McSweeney, P.L.H., Fox, P.F., Eds.; Springer: Boston, MA, USA, 2013; pp. 135–160. [Google Scholar]
- Formaggioni, P.; Summer, A.; Malacarne, M.; Mariani, P. Milk protein polymorphism: Detection and diffusion of the genetic variants in Bos genus. Ann. Della Fac. Di Med. Vet. Univ. Di Parma 2000, 29, 127–165. [Google Scholar]
- Sebastiani, C.; Arcangeli, C.; Ciullo, M.; Torricelli, M.; Cinti, G.; Fisichella, S.; Biagetti, M. Frequencies Evaluation of β-Casein Gene Polymorphisms in Dairy Cows Reared in Central Italy. Animals 2020, 10, 252. [Google Scholar] [CrossRef]
- Barroso, A.; Dunner, S.; Canon, J. Technical note: Use of PCR-single-strand conformation polymorphism analysis for detection of bovine β-casein variants A1, A2, A3, and B. J. Anim. Sci. 1999, 77, 2629–2632. [Google Scholar] [CrossRef] [PubMed]
- McLean, D.M.; Graham, E.R.B.; Ponzoni, R.W.; Mckenzie, H.A. Effects of Milk Protein Genetic-Variants on Milk-Yield and Composition. J. Dairy Res. 1984, 51, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Dantas, A.; Kumar, H.; Prudencio, E.S.; de Avila, L.B.J.; Orellana-Palma, P.; Dosoky, N.S.; Nepovimova, E.; Kuca, K.; Cruz-Martins, N.; Verma, R.; et al. An approach on detection, quantification, technological properties, and trends market of A2 cow milk. Food Res. Int. 2023, 167, 112690. [Google Scholar] [CrossRef] [PubMed]
- Nadugala, B.H.; Pagel, C.N.; Raynes, J.K.; Ranadheera, C.S.; Logan, A. The effect of casein genetic variants, glycosylation and phosphorylation on bovine milk protein structure, technological properties, nutrition and product manufacture. Int. Dairy J. 2022, 133, 105440. [Google Scholar] [CrossRef]
- Sanchez, M.P.; Fritz, S.; Patry, C.; Delacroix-Buchet, A.; Boichard, D. Frequencies of milk protein variants and haplotypes estimated from genotypes of more than 1 million bulls and cows of 12 French cattle breeds. J. Dairy Sci. 2020, 103, 9124–9141. [Google Scholar] [CrossRef]
- Daniloski, D.; McCarthy, N.A.; Huppertz, T.; Vasiljevic, T. What is the impact of amino acid mutations in the primary structure of caseins on the composition and functionality of milk and dairy products? Curr. Res. Food. Sci. 2022, 5, 1701–1712. [Google Scholar] [CrossRef]
- Hoque, M.; Mondal, S. Safety of Milk and Dairy Products. In Food Safety and Human Health; Singh, R.L., Mondal, S., Eds.; Academic Press: London, UK, 2019; p. xvii, 385p. [Google Scholar]
- Giribaldi, M.; Lamberti, C.; Cirrincione, S.; Giuffrida, M.G.; Cavallarin, L. A2 Milk and BCM-7 Peptide as Emerging Parameters of Milk Quality. Front. Nutr. 2022, 9, 842375. [Google Scholar] [CrossRef]
- Markoska, T.; Daniloski, D.; Vasiljevic, T.; Huppertz, T. Structural Changes of β-Casein Induced by Temperature and pH Analysed by Nuclear Magnetic Resonance, Fourier-Transform Infrared Spectroscopy, and Chemometrics. Molecules 2021, 26, 7650. [Google Scholar] [CrossRef] [PubMed]
- Jinsmaa, Y.; Yoshikawa, M. Enzymatic release of neocasomorphin and β-casomorphin from bovine β-casein. Peptides 1999, 20, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Zhou, Y.; Li, H.; Chen, Y.; Dominik, G.; Dong, J.; Tang, Y.; Saavedra, J.M.; Liu, J. Effectiveness of Growing-Up Milk Containing Only A2 β-Casein on Digestive Comfort in Toddlers: A Randomized Controlled Trial in China. Nutrients 2023, 15, 1313. [Google Scholar] [CrossRef]
- Edwards, T.S.; Dawson, K.L.; Keenan, J.I.; Day, A.S. A Simple Method to Generate β-Casomorphin-7 by in Vitro Digestion of Casein from Bovine Milk. J. Funct. Foods 2021, 85, 104631. [Google Scholar] [CrossRef]
- de Vasconcelos, M.L.; Oliveira, L.M.F.S.; Hill, J.P.; Vidal, A.M.C. Difficulties in Establishing the Adverse Effects of β-Casomorphin-7 Released from β-Casein Variants—A Review. Foods 2023, 12, 3151. [Google Scholar] [CrossRef]
- Dantas, A.; Pierezan, M.D.; Camelo-Silva, C.; Zanetti, V.; Pimentel, T.C.; da Cruz, A.G.; Verruck, S. A discussion on A1-free milk: Nuances and comments beyond implications to the health. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2024; ISSN 1043-4526. [Google Scholar] [CrossRef]
- Asledottir, T.; Le, T.T.; Petrat-Melin, B.; Devold, T.G.; Larsen, L.B.; Vegarud, G.E. Identification of bioactive peptides and quantification of β-casomorphin-7 from bovine β-casein A1, A2 and I after ex vivo gastrointestinal digestion. Int. Dairy J. 2017, 71, 98–106. [Google Scholar] [CrossRef]
- Cattaneo, S.; Masotti, F.; Stuknyte, M.; De Noni, I. Impact of in vitro static digestion method on the release of beta-casomorphin-7 from bovine milk and cheeses with A1 or A2 beta-casein phenotypes. Food Chem. 2023, 404, 134617. [Google Scholar] [CrossRef]
- White, C.L.; Bray, G.A.; York, D.A. Intragastric β-casomorphin attenuates the suppression of fat intake by enterostatin. Peptides 2000, 21, 1377–1381. [Google Scholar] [CrossRef]
- Lambers, T.T.; Broeren, S.; Heck, J.; Bragt, M.; Huppertz, T. Processing affects beta-casomorphin peptide formation during simulated gastrointestinal digestion in both A1 and A2 milk. Int. Dairy J. 2021, 121, 105099. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Busetti, F.; Smolenski, G.; Johnson, S.K.; Solah, V.A. Release of beta-casomorphins during gastrointestinal digestion of reconstituted milk after heat treatment. LWT-Food Sci. Technol. 2021, 136, 110312. [Google Scholar] [CrossRef]
- Daniloski, D.; McCarthy, N.A.; Vasiljevic, T. Bovine β-Casomorphins: Friends or Foes? A comprehensive assessment of evidence from and studies. Trends Food Sci. Technol. 2021, 116, 681–700. [Google Scholar] [CrossRef]
- Chen, L.; Liu, B.; Zhao, J.; Jiang, T.; Zhou, W.; Li, J.; Liu, Y.; Qiao, W. Application of Beta-Casein A2 and Composition Thereof in Promoting Proliferation of Bifidobacterium. Patent WO/2021/003741, 8 June 2022. [Google Scholar]
- Fernandez-Rico, S.; Mondragon, A.D.C.; Lopez-Santamarina, A.; Cardelle-Cobas, A.; Regal, P.; Lamas, A.; Ibarra, I.S.; Cepeda, A.; Miranda, J.M. A2 Milk: New Perspectives for Food Technology and Human Health. Foods 2022, 11, 2387. [Google Scholar] [CrossRef] [PubMed]
- Claustre, J.; Toumi, F.; Trompette, A.; Jourdan, G.; Guignard, H.; Chayvialle, J.A.; Plaisancie, P. Effects of peptides derived from dietary proteins on mucus secretion in rat jejunum. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G521–G528. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Claustre, J.; Caillon, F.; Jourdan, G.; Chayvialle, J.A.; Plaisancié, P. Milk bioactive peptides and β-casomorphins induce mucus release in rat jejunum. J. Nutr. 2003, 133, 3499–3503. [Google Scholar] [CrossRef] [PubMed]
- Zoghbi, S.; Trompette, A.; Claustre, J.; El Homsi, M.; Garzón, J.; Scoazec, J.Y.; Plaisancié, P. β-Casomorphin-7 regulates the secretion and expression of gastrointestinal mucins through a μ-opioid pathway. Am. J. Physiol.-Gastr. Liver 2006, 290, G1105–G1113. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, M.R.; Kapila, R.; Saliganti, V. Consumption of β-casomorphins-7/5 induce inflammatory immune response in mice gut through Th2 pathway. J. Funct. Foods 2014, 8, 150–160. [Google Scholar] [CrossRef]
- Ul Haq, M.R.; Kapila, R.; Sharma, R.; Saliganti, V.; Kapila, S. Comparative evaluation of cow β-casein variants (A1/A2) consumption on Th-mediated inflammatory response in mouse gut. Eur. J. Nutr. 2014, 53, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Milan, A.M.; Shrestha, A.; Karlström, H.J.; Martinsson, J.A.; Nilsson, N.J.; Perry, J.K.; Day, L.; Barnett, M.P.G.; Cameron-Smith, D. Comparison of the impact of bovine milk β-casein variants on digestive comfort in females self-reporting dairy intolerance: A randomized controlled trial. Am. J. Clin. Nutr. 2020, 111, 149–160. [Google Scholar] [CrossRef]
- Sheng, X.; Li, Z.; Ni, J.; Yelland, G. Effects of Conventional Milk Versus Milk Containing Only A2 beta-Casein on Digestion in Chinese Children: A Randomized Study. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 375–382. [Google Scholar] [CrossRef]
- Barnett, M.P.; McNabb, W.C.; Roy, N.C.; Woodford, K.B.; Clarke, A.J. Dietary A1 beta-casein affects gastrointestinal transit time, dipeptidyl peptidase-4 activity, and inflammatory status relative to A2 beta-casein in Wistar rats. Int. J. Food Sci. Nutr. 2014, 65, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Kappes, R.; Schneider, V.; Schweizer, H.; Nüske, S.; Knob, D.A.; Neto, A.T.; Scholz, A.M. Effect of β-casein A1 or A2 milk on body composition, milk intake, and growth in Holstein, Simmental, and crossbred dairy calves of both sexes. J. Dairy Sci. 2024. [Google Scholar] [CrossRef] [PubMed]
- Reiche, A.M.; Martín-Hernández, M.C.; Spengler Neff, A.; Bapst, B.; Fleuti, C.; Dohme-Meier, F.; Hess, H.D.; Egger, L.; Portmann, R. The A1/A2 β-casein genotype of cows, but not their horn status, influences peptide generation during simulated digestion of milk. J. Dairy Sci. 2024, 13, S0022-0302(24)00574-5. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Liu, J.J.; Yang, D.; Xu, H.Q. Effect of β-Casomorphin-7 on Intestinal Mucosal Immunity in Aged Mice. Kafkas Univ. Veter. Fak. Derg. 2019, 25, 689–696. [Google Scholar] [CrossRef]
- Li, X.; Lu, X.; Liu, M.; Zhang, Y.; Jiang, Y.; Yang, X.; Man, C. The Immunomodulatory Effects of A2 β-Casein on Immunosuppressed Mice by Regulating Immune Responses and the Gut Microbiota. Nutrients 2024, 16, 519. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.F.; Chen, W.H.; Zhang, Y.S.; Zou, S.X. Effects of intra-gastric beta-casomorphin-7 on somatostatin and gastrin gene expression in rat gastric mucosa. World J. Gastroenterol. 2007, 13, 2094–2099. [Google Scholar] [CrossRef] [PubMed]
- Han, D.N.; Zhang, D.H.; Wang, L.P.; Zhang, Y.S. Protective effect of beta-casomorphin-7 on cardiomyopathy of streptozotocin-induced diabetic rats via inhibition of hyperglycemia and oxidative stress. Peptides 2013, 44, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Miao, J.F.; Zhang, Y.S. Protective effect of β-casomorphin-7 on type 1 diabetes rats induced with streptozotocin. Peptides 2010, 31, 1725–1729. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Miao, J.F.; Ma, C.; Sun, G.J.; Zhang, Y.S. ß-Casomorphin-7 Cause Decreasing in Oxidative Stress and Inhibiting NF-?B-iNOS-NO Signal Pathway in Pancreas of Diabetes Rats. J. Food Sci. 2012, 77, C278–C282. [Google Scholar] [CrossRef]
- Zhang, W.; Miao, J.F.; Ma, C.; Han, D.N.; Zhang, Y.S. β-Casomorphin-7 attenuates the development of nephropathy in type I diabetes via inhibition of epithelial-mesenchymal transition of renal tubular epithelial cells. Peptides 2012, 36, 186–191. [Google Scholar] [CrossRef]
- Zhang, W.; Miao, J.F.; Wang, S.S.; Zhang, Y.S. The Protective Effects of Beta-Casomorphin-7 against Glucose -Induced Renal Oxidative Stress In Vivo and Vitro. PLoS ONE 2013, 8, e63472. [Google Scholar] [CrossRef]
- Guantario, B.; Giribaldi, M.; Devirgiliis, C.; Finamore, A.; Colombino, E.; Capucchio, M.T.; Evangelista, R.; Motta, V.; Zinno, P.; Cirrincione, S.; et al. A Comprehensive Evaluation of the Impact of Bovine Milk Containing Different Beta-Casein Profiles on Gut Health of Ageing Mice. Nutrients 2020, 12, 2147. [Google Scholar] [CrossRef] [PubMed]
- de Gaudry, D.K.; Lohner, S.; Schmucker, C.; Kapp, P.; Motschall, E.; Horrlein, S.; Roger, C.; Meerpohl, J.J. Milk A1 beta-casein and health-related outcomes in humans: A systematic review. Nutr. Rev. 2019, 77, 278–306. [Google Scholar] [CrossRef]
- Jianqin, S.; Leiming, X.; Lu, X.; Yelland, G.W.; Ni, J.; Clarke, A.J. Effects of milk containing only A2 beta casein versus milk containing both A1 and A2 beta casein proteins on gastrointestinal physiology, symptoms of discomfort, and cognitive behavior of people with self-reported intolerance to traditional cows’ milk. Nutr. J. 2016, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Woodford, K.; Kukuljan, S.; Ho, S. Milk Intolerance, Beta-Casein and Lactose. Nutrients 2015, 7, 7285–7297. [Google Scholar] [CrossRef]
- Brooke-Taylor, S.; Dwyer, K.; Woodford, K.; Kost, N. Systematic Review of the Gastrointestinal Effects of A1 Compared with A2 beta-Casein. Adv. Nutr. 2017, 8, 739–748. [Google Scholar] [CrossRef]
- Kaplan, M.; Baydemir, B.; Gunar, B.B.; Arslan, A.; Duman, H.; Karav, S. Benefits of A2 Milk for Sports Nutrition, Health and Performance. Front. Nutr. 2022, 9, 935344. [Google Scholar] [CrossRef]
- Oglobline, A.N.; Padula, M.P.; Doble, P.A. Quality control of A1-free dairy. Food Control 2022, 135, 108685. [Google Scholar] [CrossRef]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Dairy products and the risk of type 2 diabetes: A systematic review and dose-response meta-analysis of cohort studies. Am. J. Clin. Nutr. 2013, 98, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
- Norris, J.M.; Barriga, K.; Klingensmith, G.; Hoffman, M.; Eisenbarth, G.S.; Erlich, H.A.; Rewers, M. Timing of initial cereal exposure in infancy and risk of islet autoimmunity. JAMA-J. Am. Med. Assoc. 2003, 290, 1713–1720. [Google Scholar] [CrossRef]
- Couper, J.J.; Steele, C.; Beresford, S.; Powell, T.; McCaul, K.; Pollard, A.; Gellert, S.; Tait, B.; Harrison, L.C.; Colman, P.G. Lack of association between duration of breast-feeding or introduction of cow’s milk and development of islet autoimmunity. Diabetes 1999, 48, 2145–2149. [Google Scholar] [CrossRef] [PubMed]
- Savilahti, E.; Saarinen, K.M. Early infant feeding and type 1 diabetes. Eur. J. Nutr. 2009, 48, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Chia, J.S.J.; McRae, J.L.; Kukuljan, S.; Woodford, K.; Elliott, R.B.; Swinburn, B.; Dwyer, K.M. A1 beta-casein milk protein and other environmental pre-disposing factors for type 1 diabetes. Nutr. Diabetes 2017, 7, e274. [Google Scholar] [CrossRef] [PubMed]
- Birgisdottir, B.E.; Hill, J.P.; Thorsson, A.V.; Thorsdottir, I. Lower consumption of cow milk protein A1 beta-casein at 2 years of age, rather than consumption among 11- to 14-year-old adolescents, may explain the lower incidence of type 1 diabetes in Iceland than in Scandinavia. Ann. Nutr. Metab. 2006, 50, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, S.M.; Läärä, E.; Hyppönen, E.; Reijonen, H.; Räsänen, L.; Aro, A.; Knip, M.; Ilonen, J.; Äkerblom, H.K.; Grp, C.D.F.S. Cow’s milk consumption, HLA-DQB1 genotype, and type 1 diabetes -: A nested case-control study of siblings of children with diabetes. Diabetes 2000, 49, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Laugesen, M.; Elliott, R. Ischaemic heart disease, Type 1 diabetes, and cow milk A1 β-casein. N. Z. Med. J. 2003, 116, U295. [Google Scholar]
- Wasilewska, J.; Sienkiewicz-Szlapka, E.; Kuzbida, E.; Jarmolowska, B.; Kaczmarski, M.; Kostyra, E. The exogenous opioid peptides and DPPIV serum activity in infants with apnoea expressed as apparent life threatening events (ALTE). Neuropeptides 2011, 45, 189–195. [Google Scholar] [CrossRef]
- Tailford, K.A.; Berry, C.L.; Thomas, A.C.; Campbell, J.H. A casein variant in cow’s milk is atherogenic. Atherosclerosis 2003, 170, 13–19. [Google Scholar] [CrossRef]
- Chin-Dusting, J.; Shennan, J.; Jones, E.; Williams, C.; Kingwell, B.; Dart, A. Effect of dietary supplementation with β-casein A1 or A2 on markers of disease development in individuals at high risk of cardiovascular disease. Br. J. Nutr. 2006, 95, 136–144. [Google Scholar] [CrossRef]
- Venn, B.J.; Skeaff, C.M.; Brown, R.; Mann, J.I.; Green, T.J. A comparison of the effects of A1 and A2 β-casein protein variants on blood cholesterol concentrations in New Zealand adults. Atherosclerosis 2006, 188, 175–178. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, Z.; Wang, X.; Cade, R.; Elmir, Z.; Fregly, M. Relation of beta-casomorphin to apnea in sudden infant death syndrome. Peptides 2003, 24, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, S.; Frediani, T.; Zingoni, A.M.; Ferruzzi, F.; Giardini, O.; Quintieri, F.; Barbato, M.; Deufemia, P.; Cardi, E. Food allergy and infantile autism. Panminerva Med. 1995, 37, 137–141. [Google Scholar] [PubMed]
- Reichelt, K.L.; Knivsberg, A.M. Can the pathophysiology of autism be explained by the nature of discovered urine peptides? Nutr. Neurosci. 2003, 6, 19–28. [Google Scholar] [CrossRef]
- Sokolov, O.; Kost, N.; Andreeva, O.; Korneeva, E.; Meshavkin, V.; Tarakanova, Y.; Dadayan, A.; Zolotarev, Y.; Grachev, S.; Mikheeva, I.; et al. Autistic children display elevated urine levels of bovine casomorphin-7 immunoreactivity. Peptides 2014, 56, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Dubynin, V.A.; Malinovskaya, I.V.; Ivleva, Y.A.; Andreeva, L.A.; Kamenskii, A.A.; Ashmarin, I.P. Delayed behavioral effects of beta-casomorphin-7 depend on age and gender of albino rat pups. Bull. Exp. Biol. Med. 2000, 130, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Dubynin, V.A.; Ivleva, Y.A.; Stovolosov, I.S.; Belyaeva, Y.A.; Dobryakova, Y.V.; Andreeva, L.A.; Alfeeva, L.Y.; Kamenskii, A.A.; Myasoedov, N.F. Effect of beta-casomorphines on mother-oriented (“child’s”) behavior of white rats. Dokl. Biol. Sci. 2007, 412, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Dubynin, V.A.; Malinovskaia, I.V.; Beliaeva, I.A.; Stovolosov, I.S.; Bespalova, Z.D.; Andreeva, L.A.; Kamenski, A.A.; Miasoedov, N.F. Delayed effect of exorphins on learning of albino rat pups. Izv. Akad. Nauk Ser. Biol. 2008, 35, 53–60. [Google Scholar] [CrossRef]
- Maklakova, A.S.; Dubynin, V.A.; Nazarenko, I.V.; Nezavibat’ko, V.N.; Alfeeva, L.A.; Kamenskii, A.A. Effects of β-casomorphin-7 on different types of learning in white rats. Bull. Exp. Biol. Med. 1995, 120, 1121–1124. [Google Scholar] [CrossRef]
- Hunter, L.C.; O’Hare, A.; Herron, W.J.; Fisher, L.A.; Jones, G.E. Opioid peptides and dipeptidyl peptidase in autism. Dev. Med. Child Neurol. 2003, 45, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Cass, H.; Gringras, P.; March, J.; McKendrick, I.; O’Hare, A.E.; Owen, L.; Pollin, C. Absence of urinary opioid peptides in children with autism. Arch. Dis. Child. 2008, 93, 745–750. [Google Scholar] [CrossRef]
- Fiedorowicz, E.; Jarmolowska, B.; Iwan, M.; Kostyra, E.; Obuchowicz, R.; Obuchowicz, M. The influence of μ-opioid receptor agonist and antagonist peptides on peripheral blood mononuclear cells (PBMCs). Peptides 2011, 32, 707–712. [Google Scholar] [CrossRef]
- Fiedorowicz, E.; Kaczmarski, M.; Cieslinska, A.; Sienkiewicz-Szlapka, E.; Jarmolowska, B.; Chwala, B.; Kostyra, E. beta-casomorphin-7 alters mu-opioid receptor and dipeptidyl peptidase IV genes expression in children with atopic dermatitis. Peptides 2014, 62, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Jarmolowska, B.; Bukalo, M.; Fiedorowicz, E.; Cieslinska, A.; Kordulewska, N.K.; Moszynska, M.; Swiatecki, A.; Kostyra, E. Role of Milk-Derived Opioid Peptides and Proline Dipeptidyl Peptidase-4 in Autism Spectrum Disorders. Nutrients 2019, 11, 87. [Google Scholar] [CrossRef]
- Lisson, M.; Lochnit, G.; Erhardt, G. Genetic variants of bovine β- and κ-casein result in different immunoglobulin E-binding epitopes after in vitro gastrointestinal digestion. J. Dairy Sci. 2013, 96, 5532–5543. [Google Scholar] [CrossRef]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.A.; López-Expósito, I.; Pihlanto, A.; Ramos, M.; Recio, I. Antioxidant activity of ovine casein hydrolysates:: Identification of active peptides by HPLC-MS/MS. Eur. Food Res. Technol. 2008, 227, 1061–1067. [Google Scholar] [CrossRef]
- Stobiecka, M.; Król, J.; Brodziak, A. Antioxidant Activity of Milk and Dairy Products. Animals 2022, 12, 245. [Google Scholar] [CrossRef]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of Milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar] [CrossRef]
- Jeong, H.; Park, Y.S.; Yoon, S.S. A2 milk consumption and its health benefits: An update. Food Sci. Biotechnol. 2024, 33, 491–503. [Google Scholar] [CrossRef]
- Giglioti, R.; Gutmanis, G.; Katiki, L.M.; Okino, C.H.; Oliveira, M.C.D.; Vercesi, A.E. New high-sensitive rhAmp method for A1 allele detection in A2 milk samples. Food Chem. 2020, 313, 126167. [Google Scholar] [CrossRef]
- Giglioti, R.; Okino, C.H.; Azevedo, B.T.; Gutmanis, G.; Katiki, L.M.; Oliveira, M.C.D.; Vercesi, A.E. Novel LNA probe-based assay for the A1 and A2 identification of β-casein gene in milk samples. Food Chem.-Mol. Sci. 2021, 3, 100055. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Munakata, K.; Muto, M.; Kuramoto, T. A highly sensitive PCR method for A1 allele detection in A2 milk samples without DNA isolation. Anim. Sci. J. 2023, 94, 13846. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Pan, S.S.; Wu, P.Z.; Li, M.; Liang, D.P. Determination of A1 and A2 β-Casein in Milk Using Characteristic Thermolytic Peptides via Liquid Chromatography-Mass Spectrometry. Molecules 2023, 28, 5200. [Google Scholar] [CrossRef] [PubMed]
- Vigolo, V.; Franzoi, M.; Cendron, F.; Salvadore, G.; Penasa, M.; Cassandro, M.; De Marchi, M. Characterization of the genetic polymorphism linked to the beta-casein A1/A2 alleles using different molecular and biochemical methods. J. Dairy Sci. 2022, 105, 8946–8955. [Google Scholar] [CrossRef] [PubMed]
- Fenaille, F.; Parisod, V.; Visani, P.; Populaire, S.; Tabet, J.C.; Guy, P.A. Modifications of milk constituents during processing: A preliminary benchmarking study. Int. Dairy J. 2006, 16, 728–739. [Google Scholar] [CrossRef]
- Daniloski, D.; Hailu, Y.; Brodkorb, A.; Vasiljevic, T.; McCarthy, N.A. Impact of β-casein phenotype on the physical properties of skim milk powders and their subsequent digestion characteristics. Food Hydrocoll. 2024, 152, 109918. [Google Scholar] [CrossRef]
- Fuerer, C.; Jenni, R.; Cardinaux, L.; Andetsion, F.; Wagniere, S.; Moulin, J.; Affolter, M. Protein fingerprinting and quantification of beta-casein variants by ultra-performance liquid chromatography-high-resolution mass spectrometry. J. Dairy Sci. 2020, 103, 1193–1207. [Google Scholar] [CrossRef] [PubMed]
- De Poi, R.; De Dominicis, E.; Gritti, E.; Fiorese, F.; Saner, S.; de Laureto, P.P. Development of an LC-MS Method for the Identification of β-Casein Genetic Variants in Bovine Milk. Food Anal. Methods 2020, 13, 2177–2187. [Google Scholar] [CrossRef]
- Singh, M.C.; Kumar, A.; Nimmanapalli, R.; Malik, M.; Aggarwal, A.; Kumar, V.; Kumar, M. High-resolution mass spectrometer-based identification of β-Casein variant (A2/A1) in the milk of Indian Holstein Friesian crossed cows. J. Food Compos. Anal. 2024, 128, 106002. [Google Scholar] [CrossRef]
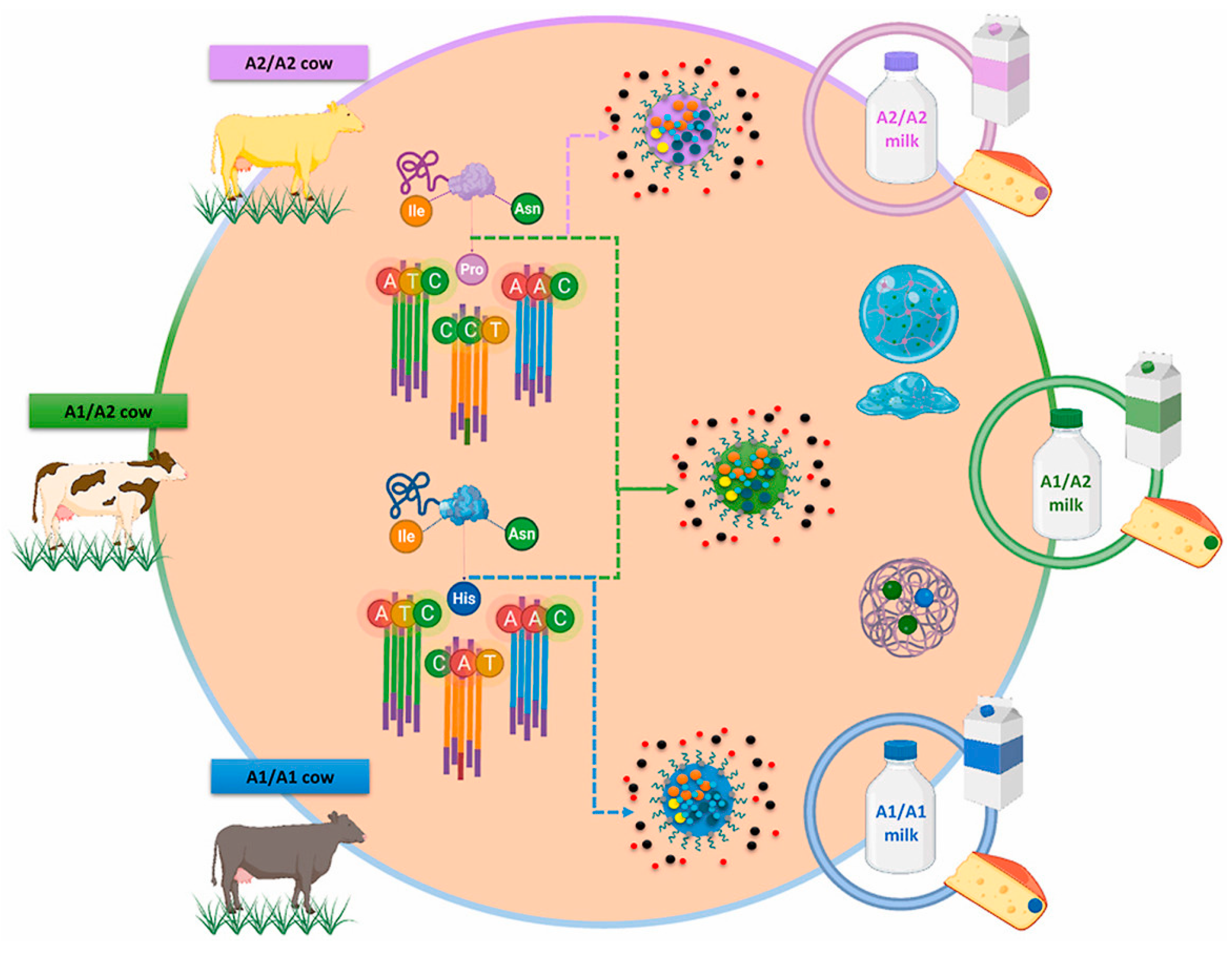

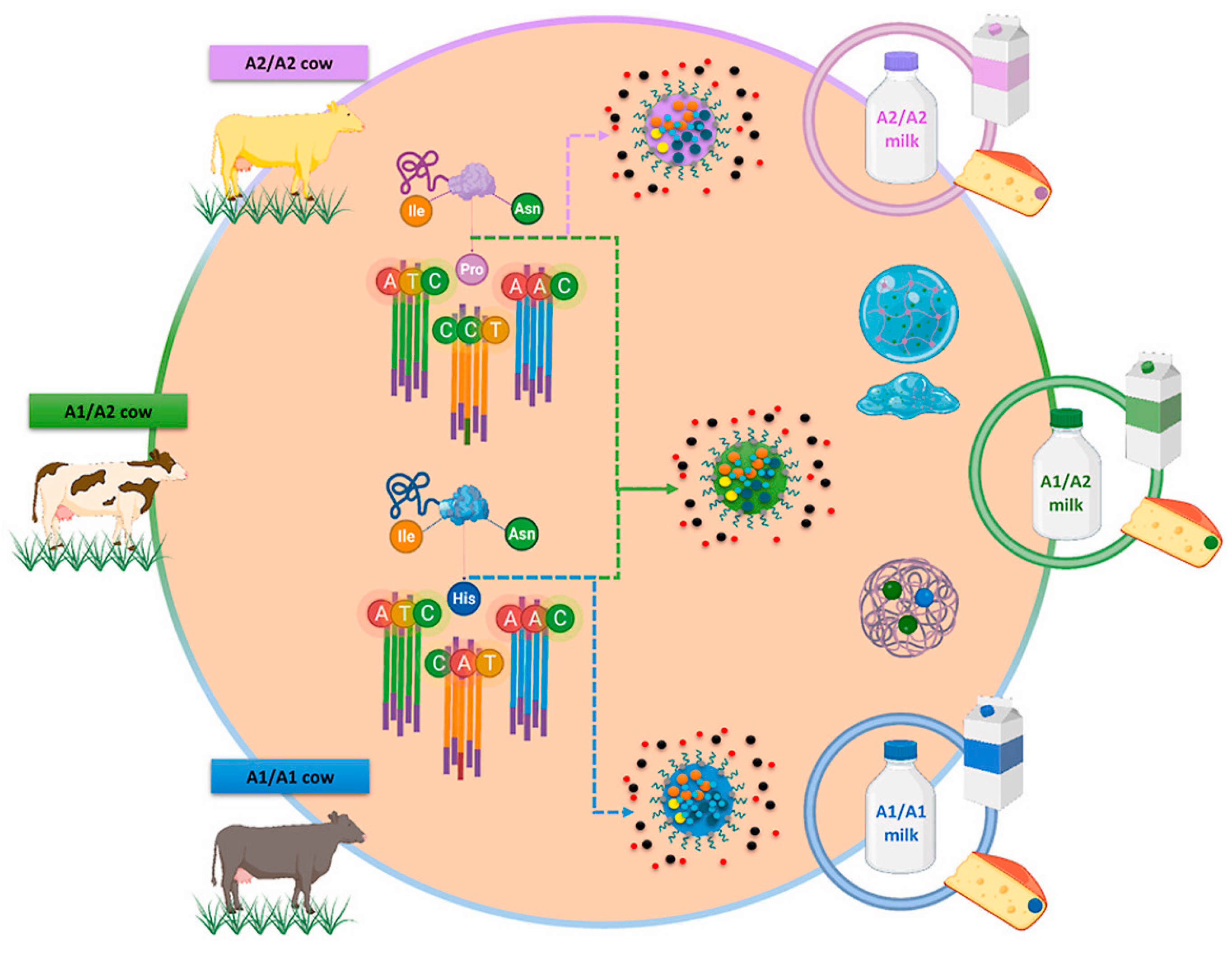{kind=link}
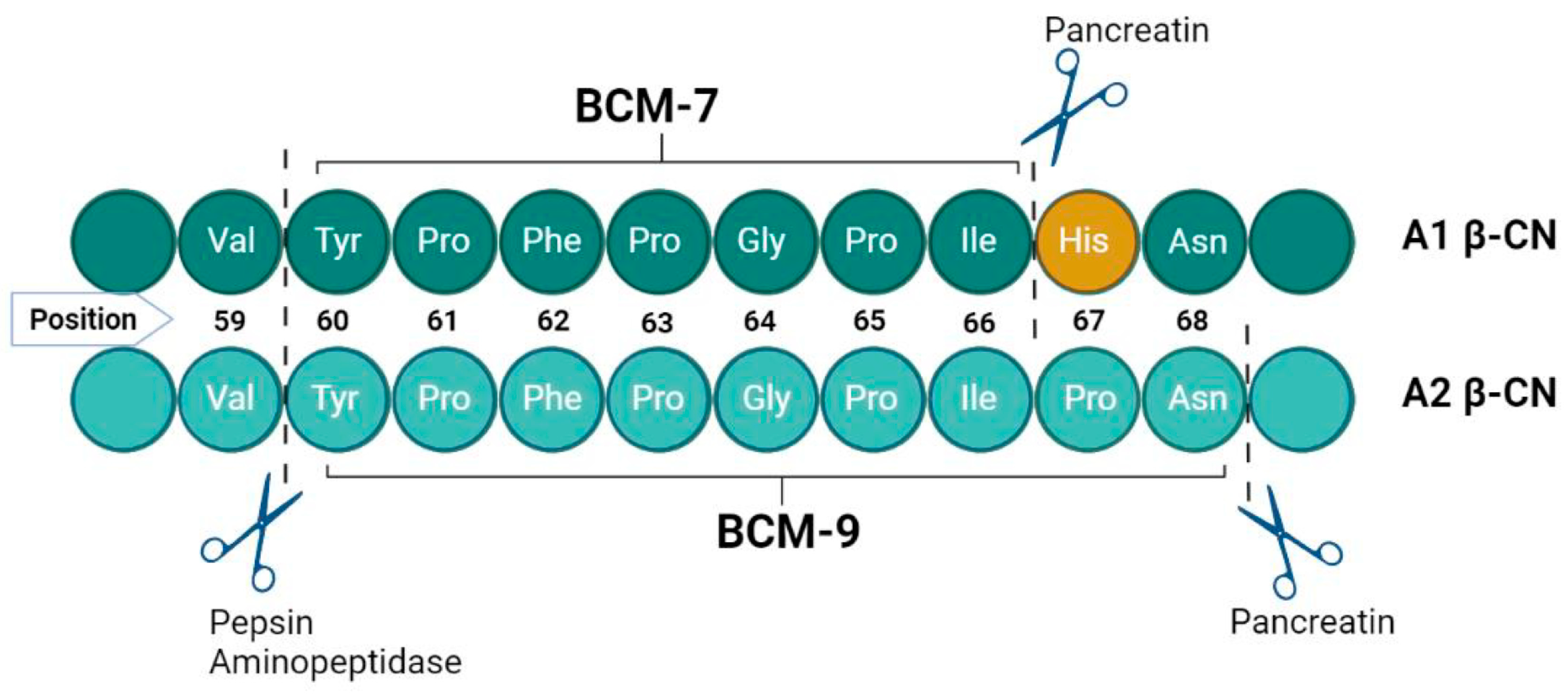{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borş, A.; Borş, S.-I.; Floriștean, V.-C. Health-Related Outcomes and Molecular Methods for the Characterization of A1 and A2 Cow’s Milk: Review and Update. Vet. Sci. 2024, 11, 172. https://doi.org/10.3390/vetsci11040172
Borş A, Borş S-I, Floriștean V-C. Health-Related Outcomes and Molecular Methods for the Characterization of A1 and A2 Cow’s Milk: Review and Update. Veterinary Sciences. 2024; 11(4):172. https://doi.org/10.3390/vetsci11040172
Chicago/Turabian StyleBorş, Alina, Silviu-Ionuț Borş, and Viorel-Cezar Floriștean. 2024. "Health-Related Outcomes and Molecular Methods for the Characterization of A1 and A2 Cow’s Milk: Review and Update" Veterinary Sciences 11, no. 4: 172. https://doi.org/10.3390/vetsci11040172
APA StyleBorş, A., Borş, S.-I., & Floriștean, V.-C. (2024). Health-Related Outcomes and Molecular Methods for the Characterization of A1 and A2 Cow’s Milk: Review and Update. Veterinary Sciences, 11(4), 172. https://doi.org/10.3390/vetsci11040172