
Pathological Study on Trigeminal Ganglionitis Among Rabid Dogs in the Philippines

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Clinical Signs
2.2. Histopathological Examination
2.3. Digitization of Tissue Slides
2.4. IHC Analysis
2.5. Indirect Immunofluorescence Double-Staining
3. Results
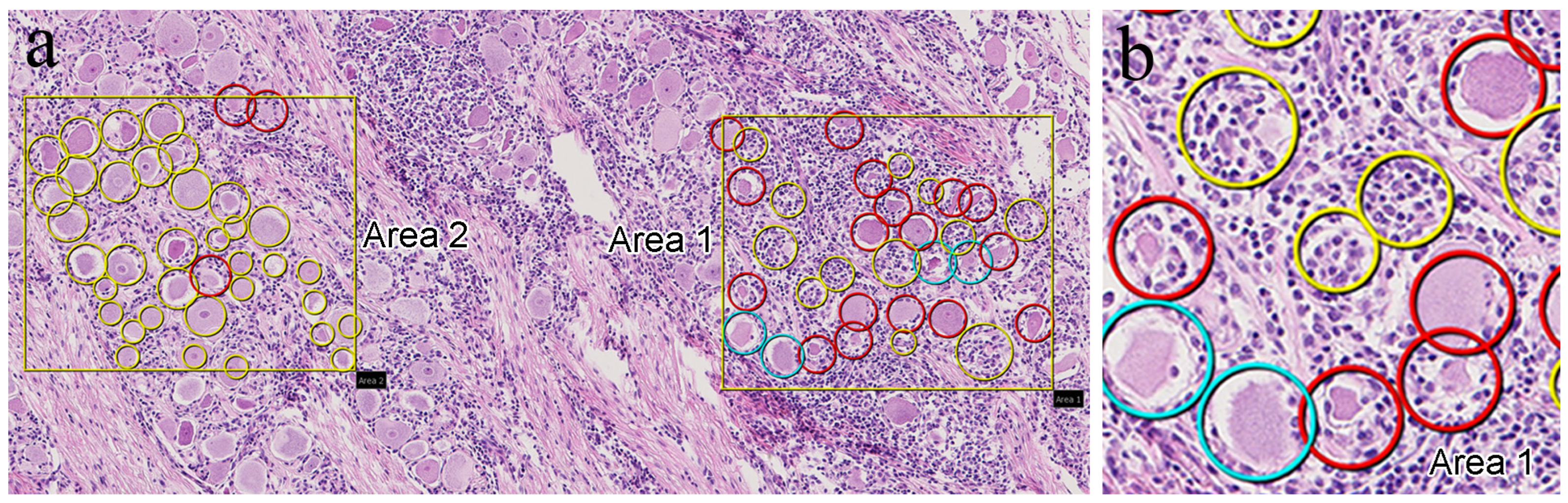
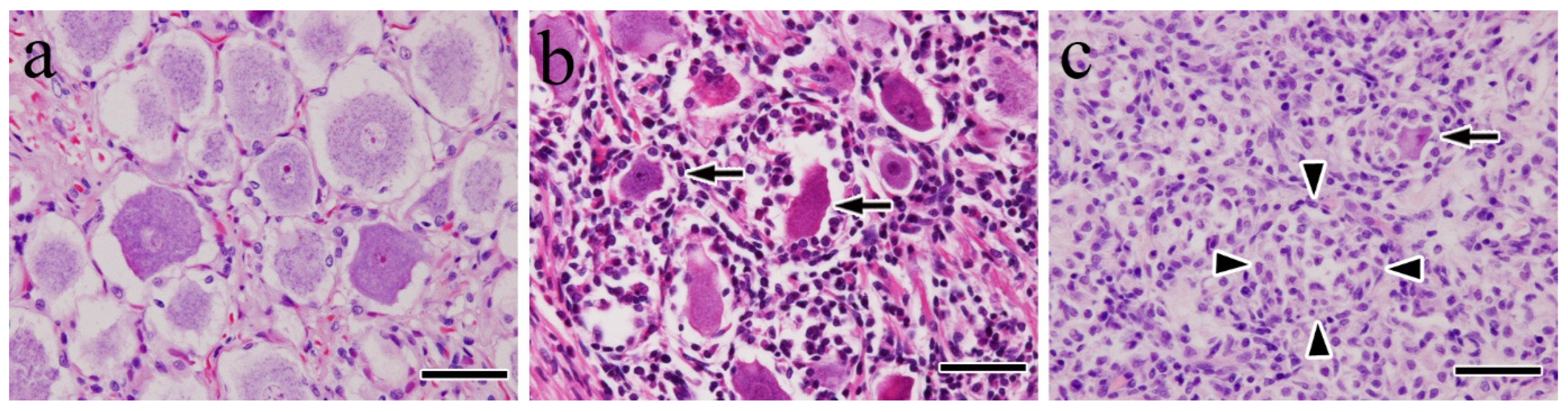
3.1. Histopathological Findings
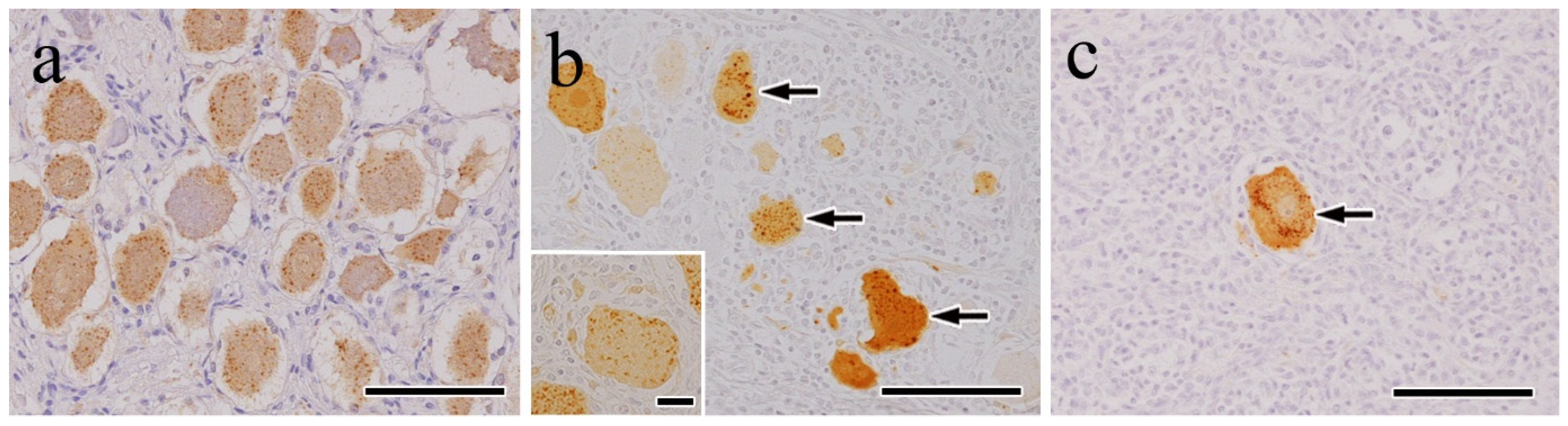
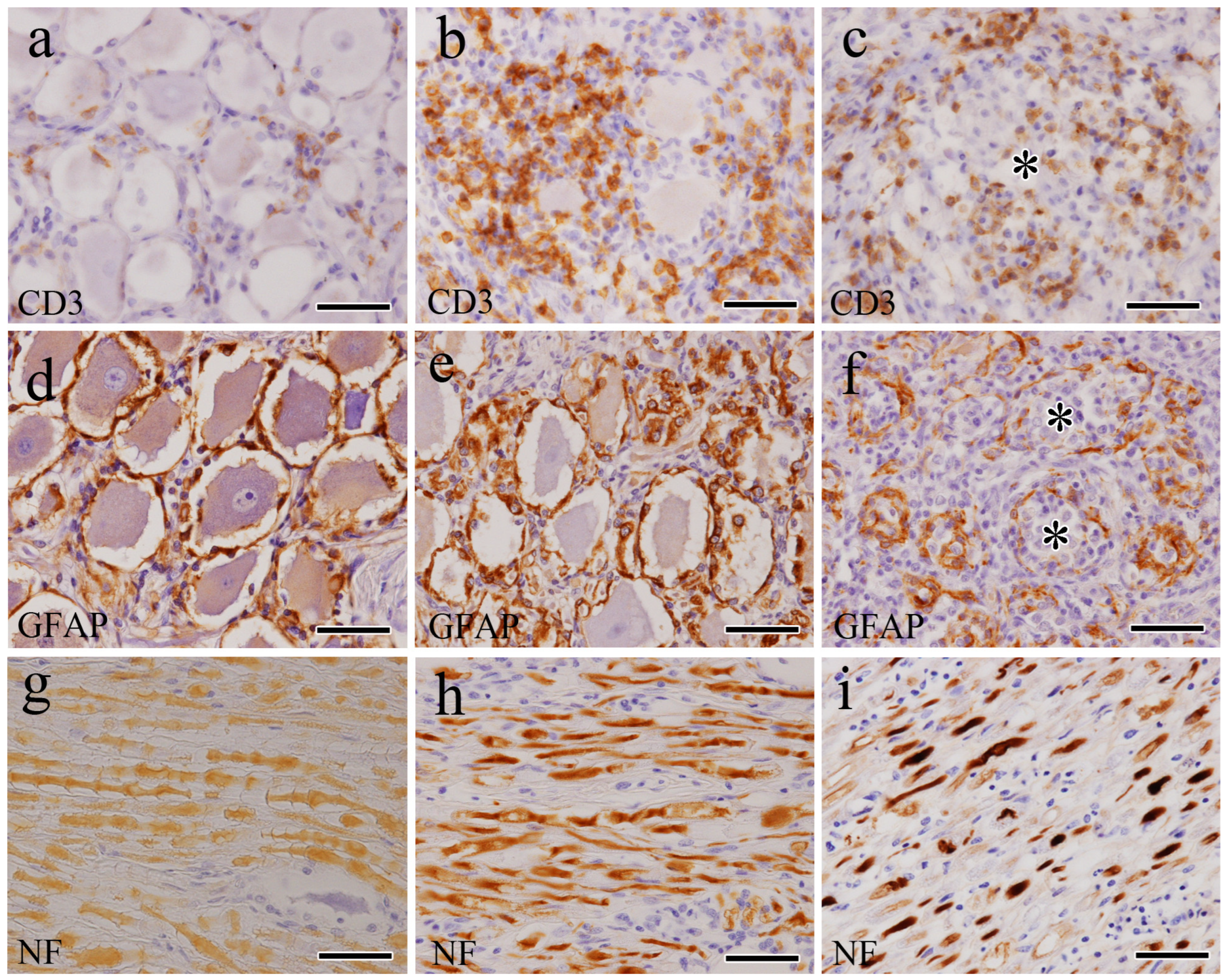
3.2. IHC Findings
3.3. Indirect Immunofluorescence Double-Stain Findings
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wunner, W.H. Rabies Virus. In Rabies; Jackson, A.C., Wunner, W.H., Eds.; Academic Press: Cambridge, MA, USA, 2007; pp. 23–68. [Google Scholar]
- Kumar, A.; Bhatt, S.; Kumar, A.; Rana, T. Canine Rabies: An Epidemiological Significance, Pathogenesis, Diagnosis, Prevention, and Public Health Issues. Comp. Immunol. Microbiol. Infect. Dis. 2023, 97, 101992. [Google Scholar] [CrossRef] [PubMed]
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Global Alliance for Rabies Control Partners for Rabies Prevention. Estimating the Global Burden of Endemic Canine Rabies. PLOS Negl. Trop. Dis. 2015, 9, e0003709. [Google Scholar]
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Müller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Primers 2017, 3, 17091. [Google Scholar] [CrossRef] [PubMed]
- Knobel, D.L.; Cleaveland, S.; Coleman, P.G.; Fèvre, E.M.; Meltzer, M.I.; Miranda, M.E.G.; Shaw, A.; Zinsstag, J.; Meslin, F.-X. Re-evaluating the Burden of Rabies in Africa and Asia. Bull. World Health Organ. 2005, 83, 360–368. [Google Scholar]
- Lentz, T.L.; Burrage, T.G.; Smith, A.L.; Crick, J.; Tignor, G.H. Is the Acetylcholine Receptor a Rabies Virus Receptor? Science 1982, 215, 182–184. [Google Scholar] [CrossRef]
- Ajayi, B.B.; Rabo, J.S.; Baba, S.S. Rabies in Apparently Healthy Dogs: Histological and Immunohistochemical Studies. Niger. Postgrad. Med. J. 2006, 13, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, A.; Charlton, K. Experimental Rabies Infection of Non-nervous Tissues in Skunks (Mephitis mephitis) and Foxes (Vulpes vulpes). Vet. Pathol. 1994, 31, 93–102. [Google Scholar] [CrossRef]
- Jackson, A.C.; Ye, H.; Phelan, C.C.; Ridaura-Sanz, C.; Zheng, Q.; Li, Z.; Wan, X.; Lopez-Corella, E. Extraneural Organ Involvement in Human Rabies. Lab. Investig. 1999, 79, 945–951. [Google Scholar]
- Jogai, S.; Radotra, B.D.; Banerjee, A.K. Rabies Viral Antigen in Extracranial Organs: A Post-Mortem Study. Neuropathol. Appl. Neurobiol. 2002, 28, 334–338. [Google Scholar] [CrossRef]
- de Morais, C.F.; de Assis, R.V. Cardiac Involvement in Human Rabies. Case Report. Rev. Inst. Med. Trop. Sao Paulo 1985, 27, 145–149. [Google Scholar] [CrossRef]
- Tobiume, M.; Sato, Y.; Katano, H.; Nakajima, N.; Tanaka, K.; Noguchi, A.; Inoue, S.; Hasegawa, H.; Iwasa, Y.; Tanaka, J.; et al. Rabies Virus Dissemination in Neural Tissues of Autopsy Cases Due to Rabies Imported into Japan from the Philippines: Immunohistochemistry. Pathol. Int. 2009, 59, 555–566. [Google Scholar] [CrossRef]
- Lackay, S.N.; Kuang, Y.; Fu, Z.F. Rabies in Small Animals. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 851–861. [Google Scholar] [CrossRef]
- Whalen, L.R.; Kitchell, R.L. Electrophysiologic Studies of the Cutaneous Nerves of the Head of the Dog. Am. J. Vet. Res. 1983, 44, 615–627. [Google Scholar] [CrossRef]
- Bathla, G.; Hegde, A.N. The Trigeminal Nerve: An Illustrated Review of Its Imaging Anatomy and Pathology. Clin. Radiol. 2013, 68, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Kent, M.; Song, R.B.; Glass, E.N.; de Lahunta, A.D. A Salivation Abnormality with Trigeminal Nerve Dysfunction in Dogs. J. Vet. Dent. 2019, 36, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, P.D.; Bush, W.W.; Glass, E.N. Trigeminal Neuropathy in Dogs: A Retrospective Study of 29 Cases (1991–2000). J. Am. Anim. Hosp. Assoc. 2002, 38, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Swain, C.E.; Cherubini, G.B.; Mantis, P. Low Field MRI Measurements of the Normal Canine Trigeminal Nerve. Front. Vet. Sci. 2020, 7, 274. [Google Scholar] [CrossRef]
- Laast, V.A.; Pardo, C.A.; Tarwater, P.M.; Queen, S.E.; Reinhart, T.A.; Ghosh, M.; Adams, R.J.; Zink, M.C.; Mankowski, J.L. Pathogenesis of Simian Immunodeficiency Virus-Induced Alterations in Macaque Trigeminal Ganglia. J. Neuropathol. Exp. Neurol. 2007, 66, 26–34. [Google Scholar] [CrossRef]
- Boonsriroj, H.; Manalo, D.L.; Kimitsuki, K.; Shimatsu, T.; Shiwa, N.; Shinozaki, H.; Takahashi, Y.; Tanaka, N.; Inoue, S.; Park, C.-H. A Pathological Study of the Salivary Glands of Rabid Dogs in the Philippines. J. Vet. Med. Sci. 2016, 78, 35–42. [Google Scholar] [CrossRef]
- Jackson, A.C. Pathogenesis. In Rabies; Jackson, A.C., Wunner, W.H., Eds.; Academic Press: Cambridge, MA, USA, 2007; pp. 341–381. [Google Scholar]
- Murphy, F.A.; Harrison, A.K.; Winn, W.C.; Bauer, S.P. Comparative Pathogenesis of Rabies and Rabies-Like Viruses: Infection of the Central Nervous System and Centrifugal Spread of Virus to Peripheral Tissues. Lab. Investig. 1973, 29, 1–16. [Google Scholar]
- Tsiang, H.; Ceccaldi, P.E.; Lycke, E.J. Rabies Virus Infection and Transport in Human Sensory Dorsal Root Ganglia Neurons. J. Gen. Virol. 1991, 72, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, C.E.; Niezgoda, M.; Hanlon, C. Rabies in Terrestrial Animals. In Rabies; Jackson, A.C., Wunner, W.H., Eds.; Aca-Demic Press: Cambridge, MA, USA, 2007; pp. 217–220. [Google Scholar]
- Cummings, J.F.; de Lahunta, A.D.; Mitchell, W.J. Ganglioradiculitis in the Dog. A Clinical, Light- and Electron-Microscopic Study. Acta Neuropathol. 1983, 60, 29–39. [Google Scholar] [CrossRef]
- Funamoto, M.; Nibe, K.; Morozumi, M.; Edamura, K.; Uchida, K. Pathological Features of Ganglioradiculitis (Sensory Neuropathy) in Two Dogs. J. Vet. Med. Sci. 2007, 69, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Porter, B.; Schatzberg, S.; McDonough, S.; Mertens, D.; de Lahunta, A.D. Ganglioradiculitis (Sensory Neuronopathy) in a Dog: Clinical, Morphologic, and Immunohistochemical Findings. Vet. Pathol. 2002, 39, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Summers, B.A.; Cummings, J.F.; de Lahunta, A. Diseases of the Peripheral Nervous System. In Veterinary Neuropathology; Mosby: St. Louis, MO, USA, 1995; pp. 412–430. [Google Scholar]
- Mitrabhakdi, E.; Shuangshoti, S.; Wannakrairot, P.; Lewis, R.A.; Susuki, K.; Laothamatas, J.; Hemachudha, T. Difference in Neuropathogenetic Mechanisms in Human Furious and Paralytic Rabies. J. Neurol. Sci. 2005, 238, 3–10. [Google Scholar] [CrossRef]
- Tangchai, P.; Vejjajiva, A. Pathology of the Peripheral Nervous System in Human Rabies. A Study of Nine Autopsy Cases. Brain 1971, 94, 299–306. [Google Scholar] [CrossRef]
- O’Toole, D.; Mills, K.; Ellis, J.; Welch, V.; Fillerup, M. Poliomyelomalacia and Ganglioneuritis in a Horse with Paralytic Rabies. J. Vet. Diagn. Invest. 1993, 5, 94–97. [Google Scholar] [CrossRef]
- Abreu, C.C.; Nakayama, P.A.; Nogueira, C.I.; Mesquita, L.P.; Lopes, P.F.R.; Wouters, F.; Varaschin, M.S., Jr.; Bezerra, P.S. Histopathology and Immunohistochemistry of Tissues Outside Central Nervous System in Bovine Rabies. J. Neurovirol. 2014, 20, 388–397. [Google Scholar] [CrossRef]
- Rossiter, J.P.; Hsu, L.; Jackson, A.C. Selective Vulnerability of Dorsal Root Ganglia Neurons in Experimental Rabies after Peripheral Inoculation of CVS-11 in Adult Mice. Acta Neuropathol. 2009, 118, 249–259. [Google Scholar] [CrossRef]
- Hamir, A.N.; Moser, G.; Rupprecht, C.E. Clinicopathologic Variation in Raccoons Infected with Different Street Rabies Virus Isolates. J. Vet. Diagn. Investig. 1996, 8, 31–37. [Google Scholar] [CrossRef]
- Pardo, C.A.; McArthur, J.C.; Griffin, J.W. HIV Neuropathy: Insights in the Pathology of HIV Peripheral Nerve Disease. J. Peripher. Nerv. Syst. 2001, 6, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Burdo, T.H.; Orzechowski, K.; Knight, H.L.; Miller, A.D.; Williams, K. Dorsal Root Ganglia Damage in SIV-Infected Rhesus Macaques: An Animal Model of HIV-Induced Sensory Neuropathy. Am. J. Pathol. 2012, 180, 1362–1369. [Google Scholar] [CrossRef] [PubMed]
- Lakritz, J.R.; Bodair, A.; Shah, N.; O’Donnell, R.; Polydefkis, M.J.; Miller, A.D.; Burdo, T.H. Monocyte Traffic, Dorsal Root Ganglion Histopathology, and Loss of Intraepidermal Nerve Fiber Density in SIV Peripheral Neuropathy. Am. J. Pathol. 2015, 185, 1912–1923. [Google Scholar] [CrossRef]
- Rizzo, S.; Basso, C.; Troost, D.; Aronica, E.; Frigo, A.C.; Driessen, A.H.G.; Thiene, G.; Wilde, A.A.M.; van der Wal, A.C.V. T-Cell-Mediated Inflammatory Activity in the Stellate Ganglia of Patients with Ion-Channel Disease and Severe Ventricular Arrhythmias. Circ. Arrhythm. Electrophysiol. 2014, 7, 224–229. [Google Scholar] [CrossRef]
- Davis, B.M.; Rall, G.F.; Schnell, M.J. Everything You Always Wanted to Know About Rabies Virus (but Were Afraid to Ask). Annu. Rev. Virol. 2015, 2, 451–471. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, J.; Song, Y.; Liu, X.; Qian, M.; Huang, P.; Li, Y.; Zhao, L.; Wang, H. Regulation of Innate Immune Responses by Rabies Virus. Animal Model. Exp. Med. 2022, 5, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Jager, S.E.; Pallesen, L.T.; Richner, M.; Harley, P.; Hore, Z.; McMahon, S.; Denk, F.; Vaegter, C.B. Changes in the Transcriptional Fingerprint of Satellite Glial Cells Following Peripheral Nerve Injury. Glia 2020, 68, 1375–1395. [Google Scholar] [CrossRef]
- Donegan, M.; Kernisant, M.; Cua, C.; Jasmin, L.; Ohara, P.T. Satellite Glial Cell Proliferation in the Trigeminal Ganglia After Chronic Constriction Injury of the Infraorbital Nerve. Glia 2013, 61, 2000–2008. [Google Scholar] [CrossRef]
- Chen, P.; Piao, X.; Bonaldo, P. Role of Macrophages in Wallerian Degeneration and Axonal Regeneration After Peripheral Nerve Injury. Acta Neuropathol. 2015, 130, 605–618. [Google Scholar] [CrossRef]
- de Lanunta, A.; Glass, E. Lower Motor Neuron: General Somatic Efferent, Cranial Nerve. In Veterinary Neuroanatomy and Clinical Neurology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 139–144. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody (a) | Host | Clone | Dilution | Antigen Retrieval (b) | Source | Target Cells |
|---|---|---|---|---|---|---|
| CD3 | Rabbit | polyclonal | Prediluted | MW, 170 W, 10 min | Agilent (Dako) | T cells |
| CD20 | Rabbit | polyclonal | 1:400 | MW, 170 W, 10 min | Thermo | B cells |
| GFAP | Rabbit | polyclonal | Prediluted | No treatment | Nichirei | Satellite cells |
| Iba-1 | Rabbit | polyclonal | 1:500 | No treatment | Wako | Macrophages |
| HLA-DR | Mouse | TAL.1B5 | 1:500 | MW, 170 W, 10 min | Agilent (Dako) | Macrophages |
| NF | Mouse | 2F11 | Prediluted | Pro-K, 37 °C, 10 min | Agilent (Dako) | Ganglions, axons |
| S-100 | Mouse | 4C4.9 | Prediluted | MW, 170 W, 10 min | Thermo | Schwann cells |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iamohbhars, N.; Cabic, A.G.B.; Markbordee, B.; Shiina, R.; Tamura, N.; Shiwa-Sudo, N.; Kimitsuki, K.; Espino, M.J.M.; Manalo, D.L.; Inoue, S.; et al. Pathological Study on Trigeminal Ganglionitis Among Rabid Dogs in the Philippines. Vet. Sci. 2025, 12, 299. https://doi.org/10.3390/vetsci12040299
Iamohbhars N, Cabic AGB, Markbordee B, Shiina R, Tamura N, Shiwa-Sudo N, Kimitsuki K, Espino MJM, Manalo DL, Inoue S, et al. Pathological Study on Trigeminal Ganglionitis Among Rabid Dogs in the Philippines. Veterinary Sciences. 2025; 12(4):299. https://doi.org/10.3390/vetsci12040299
Chicago/Turabian StyleIamohbhars, Nuttipa, Alpha Grace B. Cabic, Boonkanit Markbordee, Ryota Shiina, Natsumi Tamura, Nozomi Shiwa-Sudo, Kazunori Kimitsuki, Mark Joseph M. Espino, Daria Llenaresas Manalo, Satoshi Inoue, and et al. 2025. "Pathological Study on Trigeminal Ganglionitis Among Rabid Dogs in the Philippines" Veterinary Sciences 12, no. 4: 299. https://doi.org/10.3390/vetsci12040299
APA StyleIamohbhars, N., Cabic, A. G. B., Markbordee, B., Shiina, R., Tamura, N., Shiwa-Sudo, N., Kimitsuki, K., Espino, M. J. M., Manalo, D. L., Inoue, S., & Park, C.-H. (2025). Pathological Study on Trigeminal Ganglionitis Among Rabid Dogs in the Philippines. Veterinary Sciences, 12(4), 299. https://doi.org/10.3390/vetsci12040299