
Microscopic Visualisation of Zoonotic Arbovirus Replication in Tick Cell and Organ Cultures Using Semliki Forest Virus Reporter Systems


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Tick Organ Cultures
2.3. Viruses
2.4. Monitoring of Virus-Infected Cultures
2.5. Harvest, Fixation, and Processing of Cells for EM
3. Results
3.1. Ultrastructural Visualisation of SFV in Infected Cells
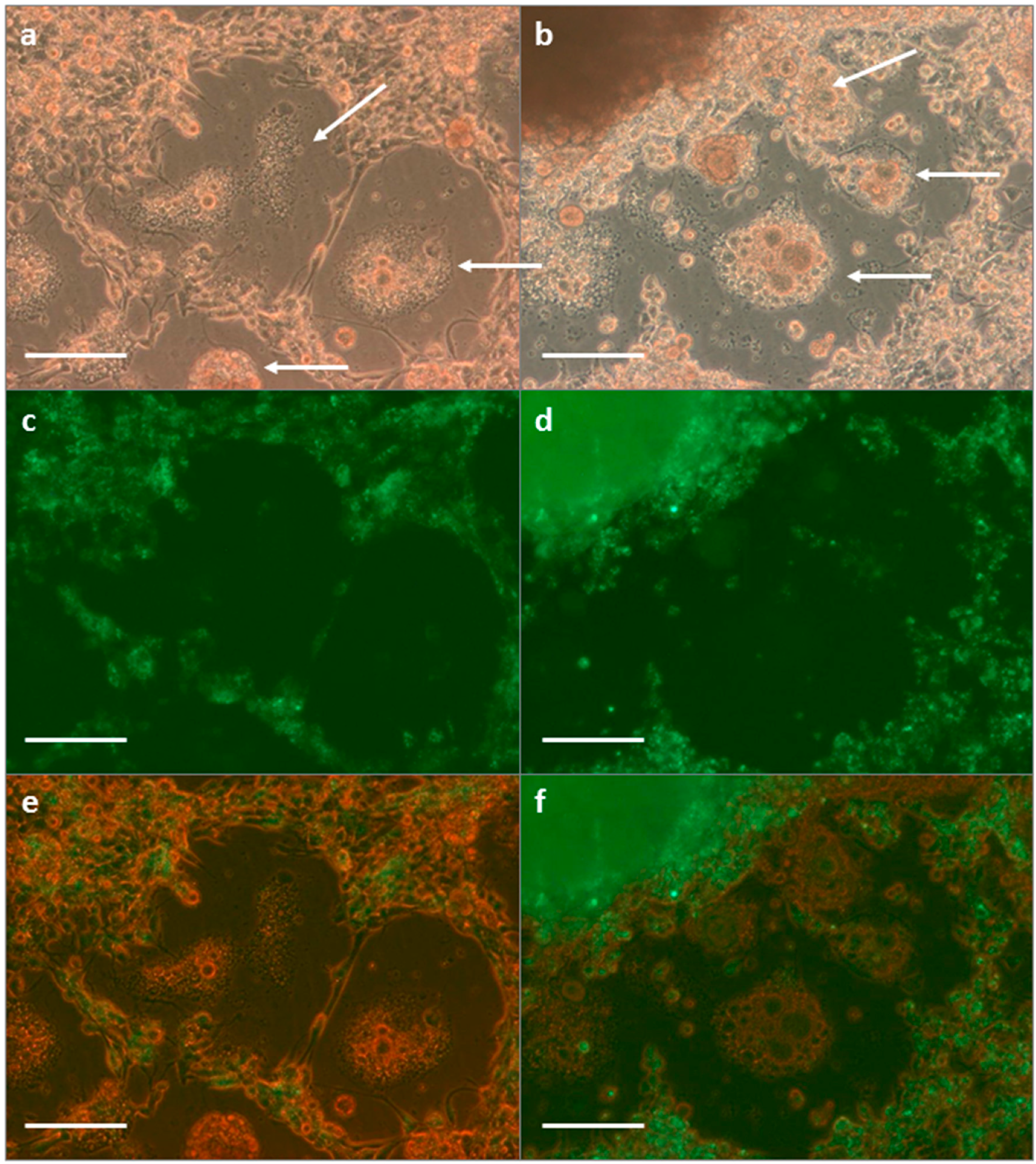
3.1.1. SFV in Tick Cell Line BDE/CTVM16
3.1.2. SFV in Mosquito Cell Line C6/36
3.1.3. SFV in BHK-21 Cells
3.2. Distribution and Cell Tropism of SFV in Infected Tick Organ and Cell Cultures
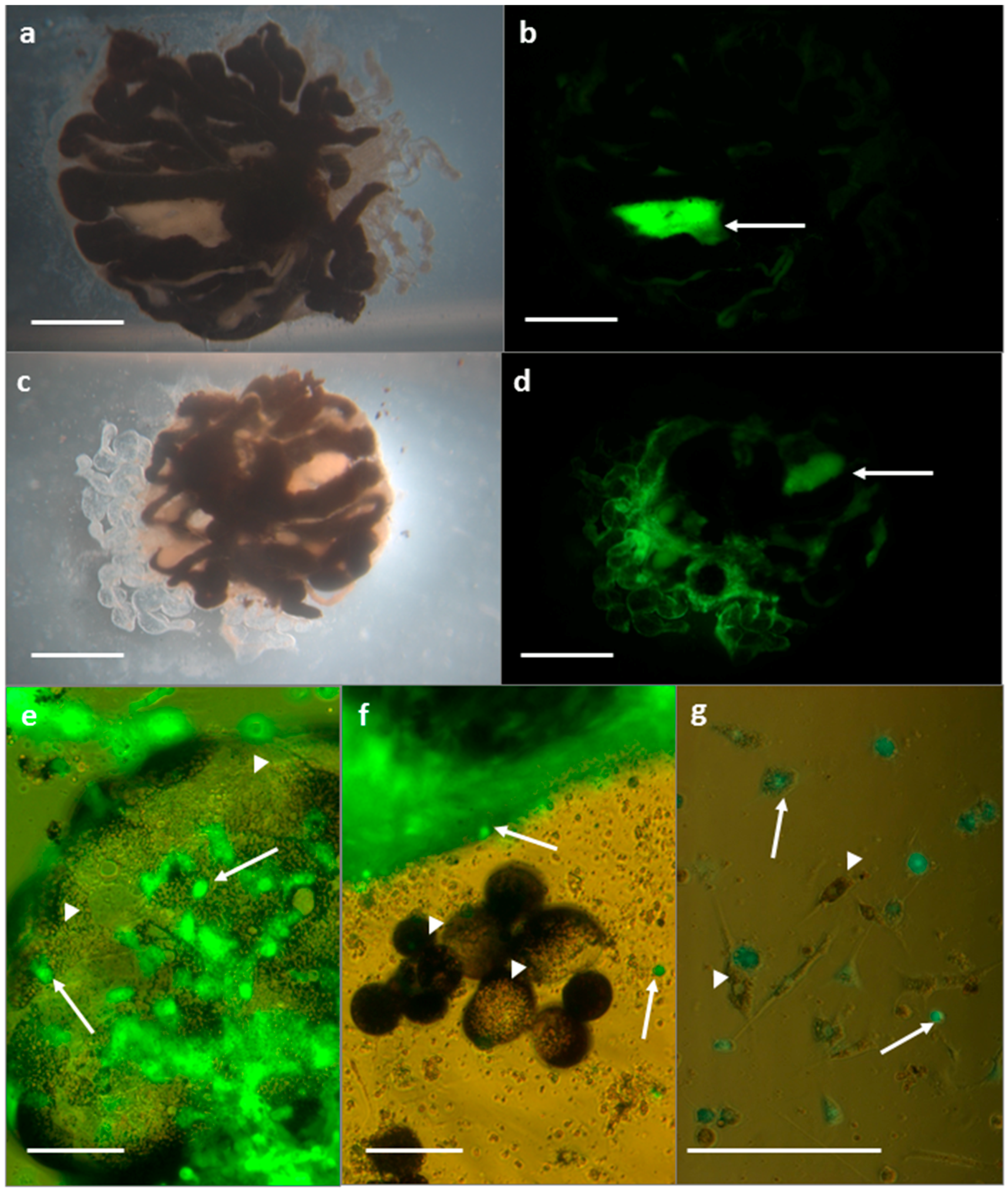
3.2.1. SFV in R. evertsi Organ Cultures
3.2.2. SFV in R. evertsi Cell Line REE/CTVM28
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CPV | cytopathic vacuole |
| EM | electron microscopy |
| hpi | hours post infection |
| MEB | midgut escape barrier |
| NSDV | Nairobi sheep disease virus |
| SFV | Semliki Forest virus |
| TBEV | tick-borne encephalitis virus |
References
- Labuda, M.; Nuttall, P.A. Tick-borne viruses. Parasitology 2004, 129, S221–S245. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, P.A. Molecular characterization of tick-virus interactions. Front. Biosci. 2009, 14, 2466–2483. [Google Scholar] [CrossRef]
- Attoui, H.; Stirling, J.M.; Munderloh, U.G.; Billoir, F.; Brookes, S.M.; Burroughs, J.N.; de Micco, P.; Mertens, P.P.C.; de Lamballerie, X. Complete sequence characterization of the genome of the St Croix River virus, a new orbivirus isolated from cells of Ixodes scapularis. J. Gen. Virol. 2001, 82, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Attoui, H. Endogenous tick viruses and modulation of tick-borne pathogen growth. Front. Cell. Infect. Microbiol. 2013, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Munz, E.; Reimann, M.; Mahnel, H. Nairobi sheep disease virus and Reovirus-like particles in the tick cell line TTC-243 from Rhipicephalus appendiculatus: Experiences with the handling of the tick cells, immunoperoxidase and ultrahistological studies. In Arboviruses in Arthropod Cells in Vitro; Yunker, C.E., Ed.; CRC Press: Boca Raton, FL, USA, 1987; Volume 1, pp. 133–147. [Google Scholar]
- Alberdi, M.P.; Dalby, M.J.; Rodriguez-Andres, J.; Fazakerley, J.K.; Kohl, A.; Bell-Sakyi, L. Detection and identification of putative bacterial endosymbionts and endogenous viruses in tick cell lines. Ticks Tick-Borne Dis. 2012, 3, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Attoui, H. Virus discovery using tick cell lines. Evol. Bioinform. 2016, 12 (Suppl. 2), 31–34. [Google Scholar] [CrossRef]
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLoS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, C.H.; Uzcategui, N.Y.; Gould, E.A.; Nuttall, P.A. Ixodid and argasid tick species and West Nile virus. Emerg. Infect. Dis. 2004, 10, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.P.; Bouquety, J.C.; Lesbordes, J.L.; Madelon, M.C.; Mathiot, C.C.; Meunier, D.M.Y.; Georges, A.J. Rift Valley fever virus and haemorrhagic fever in the Central African Republic. Ann. Inst. Pasteur/Virol. 1987, 138, 385–390. [Google Scholar] [CrossRef]
- Konstantinov, O.K. Ticks of the Ixodidae family as reservoir of arboviruses in the republic of Guinea. II. Arboviruses. Rev. Elev. Med. Vet. Pays Trop. 1990, 43, 15–22. [Google Scholar] [PubMed]
- Bell-Sakyi, L.; Kohl, A.; Bente, D.A.; Fazakerley, J.F. Tick cell lines for study of Crimean-Congo hemorrhagic fever virus and other arboviruses. Vector-Borne Zoonot. Dis. 2012, 12, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Leake, C.J.; Pudney, M.; Varma, M.G.R. Studies on arboviruses in established tick cell lines. In Invertebrate Systems in Vitro; Kurstak, E., Maramorosch, K., Dubendorfer, A., Eds.; Elsevier/North Holland Biomedical Press: Amsterdam, The Netherlands, 1980; pp. 327–335. [Google Scholar]
- Garcia, S.; Billecocq, A.; Crance, J.-M.; Munderloh, U.G.; Garin, D.; Bouloy, M. Nairovirus RNA sequences expressed by a Semliki Forest virus replicon induce RNA interference in tick cells. J. Virol. 2005, 79, 8942–8947. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.; Billecocq, A.; Crance, J.M.; Prins, M.; Garin, D.; Bouloy, M. Viral suppressors of RNA interference impair RNA silencing induced by a Semliki Forest virus replicon in tick cells. J. Gen. Virol. 2006, 87, 1985–1989. [Google Scholar] [CrossRef] [PubMed]
- Barry, G.; Alberdi, P.; Schnettler, E.; Weisheit, S.; Kohl, A.; Fazakerley, J.K.; Bell-Sakyi, L. Gene silencing in tick cell lines using small interfering or long double-stranded RNA. Exp. Appl. Acarol. 2013, 59, 319–338. [Google Scholar] [CrossRef] [PubMed]
- Moniuszko, A.; Rueckert, C.; Alberdi, M.P.; Barry, G.; Stevenson, B.; Fazakerley, J.K.; Kohl, A.; Bell-Sakyi, L. Coinfection of tick cell lines has variable effects on replication of intracellular bacterial and viral pathogens. Ticks Tick-Borne Dis. 2014, 5, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Siu, R.W.C.; Fragkoudis, R.; Simmonds, P.; Donald, C.L.; Chase-Topping, M.E.; Barry, G.; Attarzadeh-Yazdi, G.; Rodriguez-Andres, J.; Nash, A.A.; Merits, A.; et al. Antiviral RNA interference responses induced by Semliki Forest virus infection of mosquito cells: Characterization, origin, and frequency-dependent functions of virus-derived small interfering RNAs. J. Virol. 2011, 85, 2907–2917. [Google Scholar] [CrossRef] [PubMed]
- Fragkoudis, R.; Chi, Y.; Siu, R.W.C.; Barry, G.; Attarzadeh-Yadzi, G.; Merits, A.; Nash, A.A.; Fazakerley, J.F.; Kohl, A. Semliki Forest virus strongly reduces mosquito host defence signaling. Insect Mol. Biol. 2008, 17, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Pudney, M.; Leake, C.J.; Varma, M.G.R. Replication of arboviruses in arthropod in vitro systems. In Arctic and Tropical Arboviruses; Kurstak, E., Ed.; Academic Press: New York, NY, USA, 1979; pp. 245–262. [Google Scholar]
- Grimley, P.M.; Berezesky, I.K.; Friedman, R.M. Cytoplasmic structures associated with an arbovirus infection: Loci of viral ribonucleic acid synthesis. J. Virol. 1968, 2, 1326–1338. [Google Scholar] [PubMed]
- Virtanen, I.; Wartiovaara, J. Virus-induced cytoplasmic membrane structures associated with Semliki Forest virus infection studied by the freeze-etching method. J. Virol. 1974, 13, 222–225. [Google Scholar] [PubMed]
- Hellstrom, K.; Vihinen, H.; Kallio, K.; Jokilato, E.; Ahola, T. Correlative light and electron microscopy enables viral replication studies at the ultrastructural level. Methods 2015, 90, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Senigl, F. Maturation of tick-borne encephalitis virus in tick and mammalian cells. In Ticks and Tick-borne Pathogens: Into the 21st Century, Proceedings of the 3rd International Conference on Ticks and Tick-borne Pathogens, High Tatra Mountains, Slovakia, 30 August–3 September 1999; Kazimirova, M., Labuda, M., Nuttall, P.A., Eds.; Institute of Zoology, Slovak Academy of Sciences: Bratislava, Slovakia, 2000; pp. 29–38. [Google Scholar]
- Senigl, F.; Kopecky, J.; Grubhoffer, L. Distribution of E and NS1 proteins of TBE virus in mammalian and tick cells. Folia Microbiol. 2004, 49, 213–216. [Google Scholar] [CrossRef]
- Martin, H.M.; Vidler, B.O. In vitro growth of tick tissues (Rhipicephalus appendiculatus Neumann, 1901). Exp. Parasitol. 1962, 12, 192–203. [Google Scholar] [CrossRef]
- Yunker, C.E.; Cory, J. Growth of Colorado tick fever virus in primary tissue cultures of its vector, Dermacentor andersoni Stiles (Acarina: Ixodidae), with notes on tick tissue culture. Exp. Parasitol. 1967, 20, 267–277. [Google Scholar] [CrossRef]
- Chunikhin, S.P.; Khozinskaya, G.A.; Stefutkina, L.F.; Korolev, M.B. Mono- and mixed infections of tissue explants of ticks of the genus Hyalomma with tick-borne encephalitis and Powassan viruses. Parazitologiya 1984, 18, 116–122. (In Russian) [Google Scholar]
- Khozinskaya, G.A.; Chunikhin, S.P.; Khozinsky, V.V.; Stefutkina, L.F. Variability of Powassan virus cultured in tissue explants and organism of Hyalomma anatolicum ticks. Acta Virol. 1985, 29, 305–311. [Google Scholar] [PubMed]
- Bell, L.J. Tick tissue culture techniques in the study of arthropod-borne protozoa: The development of Theileria annulata in organ cultures of Hyalomma anatolicum anatolicum. In Acarology 6; Griffiths, D.A., Bowman, C.E., Eds.; Ellis Horwood Ltd.: Chichester, UK, 1984; Volume 2, pp. 1089–1095. [Google Scholar]
- Fragkoudis, R.; Breakwell, L.; McKimmie, C.; Boyd, A.; Barry, G.; Kohl, A.; Merits, A.; Fazakerley, J.K. The type I interferon system protects mice from Semliki Forest virus by preventing widespread virus dissemination in extraneural tissues, but does not mediate the restricted replication of avirulent virus in central nervous system neurons. J. Gen. Virol. 2007, 88, 3373–3384. [Google Scholar] [CrossRef] [PubMed]
- Tamberg, N.; Lulla, V.; Fragkoudis, R.; Lulla, A.; Fazakerley, J.K.; Merits, A. Insertion of EGFP into the replicase gene of Semliki Forest virus results in a novel, genetically stable marker virus. J. Gen. Virol. 2007, 88, 1225–1230. [Google Scholar] [CrossRef] [PubMed]
- Jose, J.; Tang, J.; Taylor, A.B.; Baker, T.S.; Kuhn, R.J. Fluorescent protein-tagged Sindbis virus E2 glycoprotein allows single particle analysis of virus budding from live cells. Viruses 2015, 7, 6182–6199. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L. Ehrlichia ruminantium grows in cell lines from four ixodid tick genera. J. Comp. Pathol. 2004, 130, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, A. Isolation of a Singh’s Aedes albopictus cell clone sensitive to Dengue and Chikungunya viruses. J. Gen. Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Attarzadeh-Yazdi, G.; Fragkoudis, R.; Chi, Y.; Siu, R.W.; Ülper, L.; Barry, G.; Rodriguez, J.A.; Nash, A.A.; Bouloy, M.; Merits, A.; et al. Cell-to-cell spread of the RNAi response suppresses Semliki Forest virus infection of mosquito cell cultures and cannot be antagonized by this virus. J. Virol. 2009, 83, 5735–5748. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, I. Characteristics of a hamster cell clone transformed by polyoma virus. J. Natl. Cancer Inst. 1963, 30, 795–815. [Google Scholar]
- Froshauer, S.; Kartenbeck, J.; Helenius, A. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1988, 107, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Frolova, E.I.; Gorchakov, R.; Pereboeva, L.; Atasheva, S.; Frolov, I. Functional Sindbis virus replicative complexes are formed at the plasma membrane. J. Virol. 2010, 84, 11679–11695. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Palomar, A.M.; Bradford, E.L.; Shkap, V. Propagation of the Israeli vaccine strain of Anaplasma centrale in tick cell lines. Vet. Microbiol. 2015, 179, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Hayasaki, D.; Yoshii, K.; Ueki, T.; Iwasaki, T.; Takashima, I. Sub-genomic replicons of Tick-borne encephalitis virus. Arch. Virol. 2004, 149, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Gehrke, R.; Heinz, F.X.; Davis, N.L.; Mandl, C.W. Heterologous gene expression by infectious and replicon vectors derived from tick-borne encephalitis virus and direct comparison of this flavivirus system with an alphavirus replicon. J. Gen. Virol. 2005, 86, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Booth, T.F.; Gould, E.A.; Nuttall, P.A. Structure and morphogenesis of Dugbe virus (Bunyaviridae, Nairovirus) studied by immunogold electron microscopy of ultrathin cryosections. Virus Res. 1991, 21, 199–212. [Google Scholar] [CrossRef]
- Offerdahl, D.K.; Dorward, D.W.; Hansen, B.T.; Bloom, M.E. A three-dimensional comparison of tick-borne flavivirus infection in mammalian and tick cell lines. PLoS ONE 2012, 7, E47912. [Google Scholar] [CrossRef] [PubMed]
- Gresíková, M.; Sekeyová, M.; Tempera, G.; Guglielmino, S.; Castro, A. Identification of a Sindbis virus strain isolated from Hyalomma marginatum ticks in Italy. Acta Virol. 1978, 22, 231–232. [Google Scholar] [PubMed]
- Lwande, O.W.; Lutomiah, J.; Obanda, V.; Gakuya, F.; Mutisya, J.; Mulwa, F.; Michuki, G.; Chepkorir, E.; Fischer, A.; Venter, M.; et al. Isolation of tick and mosquito-borne arboviruses from ticks sampled from livestock and wild animal hosts in Ijara District, Kenya. Vector-Borne Zoonot. Dis. 2013, 13, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Linthicum, K.J.; Logan, T.M.; Bailey, C.L.; Gordon, S.W.; Peters, C.J.; Monath, T.P.; Osorio, J.; Francy, D.B.; McLean, R.G.; Leduc, J.W.; et al. Venezuelan equine encephalomyelitis virus infection in and transmission by the tick Amblyomma cajennense (Arachnida: Ixodidae). J. Med. Entomol. 1991, 28, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Linthicum, K.J.; Logan, T.M. Laboratory transmission of Venezuelan equine encephalitis virus by the tick Hyalomma truncatum. Trans. R. Soc. Trop. Med. Hyg. 1994, 88, 126. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | First Sample | Second Sample | |
|---|---|---|---|
| BDE/CTVM16 | Mock-infected | 24 hpi | 48 hpi |
| SFV-infected | 24 hpi | ||
| C6/36 | Mock-infected | 24 hpi | 24 hpi |
| SFV-infected | 10 hpi | ||
| BHK-21 | Mock-infected | 10 hpi | 10 hpi |
| SFV-infected | 6 hpi | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell-Sakyi, L.; Weisheit, S.; Rückert, C.; Barry, G.; Fazakerley, J.; Fragkoudis, R. Microscopic Visualisation of Zoonotic Arbovirus Replication in Tick Cell and Organ Cultures Using Semliki Forest Virus Reporter Systems. Vet. Sci. 2016, 3, 28. https://doi.org/10.3390/vetsci3040028
Bell-Sakyi L, Weisheit S, Rückert C, Barry G, Fazakerley J, Fragkoudis R. Microscopic Visualisation of Zoonotic Arbovirus Replication in Tick Cell and Organ Cultures Using Semliki Forest Virus Reporter Systems. Veterinary Sciences. 2016; 3(4):28. https://doi.org/10.3390/vetsci3040028
Chicago/Turabian StyleBell-Sakyi, Lesley, Sabine Weisheit, Claudia Rückert, Gerald Barry, John Fazakerley, and Rennos Fragkoudis. 2016. "Microscopic Visualisation of Zoonotic Arbovirus Replication in Tick Cell and Organ Cultures Using Semliki Forest Virus Reporter Systems" Veterinary Sciences 3, no. 4: 28. https://doi.org/10.3390/vetsci3040028
APA StyleBell-Sakyi, L., Weisheit, S., Rückert, C., Barry, G., Fazakerley, J., & Fragkoudis, R. (2016). Microscopic Visualisation of Zoonotic Arbovirus Replication in Tick Cell and Organ Cultures Using Semliki Forest Virus Reporter Systems. Veterinary Sciences, 3(4), 28. https://doi.org/10.3390/vetsci3040028